
ABSTRACT
Temperate regions experiencing sub-zero temperatures negatively impact grapevine yield. Potassium has been claimed as a cryoprotectant to improve cold hardiness in grapevine. This study investigated the effect of foliar application of liquid potassium-based fertilizer, ReaXTM, on cold-hardiness of grapevine Vitis spp. “Chambourcin” along with its effect on yield and fruit quality. The vines were sprayed four to five times between the fruit set and veraison stage at a concentration of 1.5% (v/v) for two consecutive seasons. Petioles were analyzed for nutrients, clusters for yield and fruit quality, and cold hardiness was determined by differential thermal analysis and bud injury assessment. Potassium deficiency was observed in all treatments, and its content did not increase significantly in treated vines. Berry potassium levels and total soluble solids were generally higher in K-treated vines, however, there was no significant effect on yield and other fruit quality traits. Significant differences in cold hardiness levels were observed in both dormant seasons. Foliar application of potassium is a promising cultural practice to increase cold hardiness, but further studies are needed to understand the limits of its effectiveness.
Introduction
Cold injury is a major problem in grapevine production, causing an average of 5–15% annual crop loss worldwide (Zabadal et al., Citation2007). Cold-sensitive Vitis vinifera cultivars are more prone to sub-zero temperatures, especially in regions of Europe and the eastern and midwestern United States (Lamichhane, Citation2021; Zabadal et al., Citation2007). Within the eastern United States, major freezing events since 2007 have caused around $250 million of economic damage to the grapevine industry annually (Poni et al., Citation2022). For example, grape growers in Ohio reported an economic loss of $4 million in 335 hectares by the 2014 polar vortex that damaged 29%, 57%, and 97% of native, hybrid, and vinifera grape cultivars, respectively (Dami and Lewis, Citation2014). Along with affecting crop growth and development, winter injury also affects the wine quality, therefore, grape growers suffer from direct crop losses as well as from indirect losses due to the reduction in wine sales (Zabadal et al., Citation2007).
Although a grapevine’s genetic makeup is the main determinant of its potential cold hardiness, management practices provide short-term or mid-term crop protection against freeze damage (Dami, Citation2022). For example, several cost-effective and easily applied sprayable products are available that can be useful in mitigating freezing injury in plants. These include cryoprotectants (glycine betadine, Agro-K’s Potassium Dextrose-Lac®- KDL), dormant oils, growth regulators (ProTone®, FrostShield®), and ice nucleation inhibitors (ice nucleation active bacteria) (Centinari et al., Citation2016; Dami, Citation2022). Potassium (K+), an essential plant nutrient, has also been considered a potential cryoprotectant to mitigate freezing injury in plants. The highly mobile K+ macronutrient plays an important role in cell division, enzyme activation, neutralization of organic acids, increasing proline, protein, and sugar synthesis and translocation. The uptake and transportation of K+ within the plant depend on the K+ availability in the soil and K+ in the leaves and storage reserves within the perennial vine structures, which can be redistributed (Rogiers et al., Citation2017).
K+ is the most abundant cation during all stages of berry development (Rogiers et al., Citation2017), which influences sugar accumulation in berries, wine pH, and color (Mpelasoka et al., Citation2003; Rogiers et al., Citation2017). Evidence of potassium playing a role in cold hardiness has been reported, where potassium application reduced shoot mortality and resulted in the accumulation of carbohydrates, abscisic acid (ABA), and phenolic compounds, increasing the overall survivability of grapevines under cold stress (Karimi, Citation2017; Sarikhani et al., Citation2014). Although the mechanism is not clear, K+ accumulation may lead to an increase in cell solute concentration (by increasing the accumulation of soluble sugars) (Karimi, Citation2017; Sarikhani et al., Citation2014) and decrease the osmotic potential (Centinari et al., Citation2016), thus reducing the cell freezing point. Along with inducing cold hardiness, potassium was also found to increase endogenous ABA concentrations (Karimi, Citation2017), which may delay grapevine budburst (Wang et al., Citation2020), thus preventing sensitive young shoots from exposure to late-spring frost conditions. Previous studies have been conducted to evaluate the effect of foliar applications of potassium-based fertilizers on cold hardiness of grapevine, however, the effectiveness of these products in field conditions is still questionable (Centinari et al., Citation2016, Citation2018; Sarikhani et al., Citation2014).
The main objective of this study was to investigate the effects of foliar application of liquid potassium-based fertilizer ReaXTM on the cold hardiness of field-grown grapevines. We hypothesized that vines fertilized with potassium would have higher cold hardiness compared to non-treated (control) vines. Since K+ accumulation influences fruit quality (sugars, pH, acidity) and plant growth, this study also evaluated if the potassium applications impacted grapevine yield or fruit quality.
Materials and Methods
Plant Material and Experimental Design
This study was conducted on field-grown mature vines of Vitis spp. “Chambourcin” trained on a high cordon trellis with a spacing of 3 m × 1.22 m. Vitis spp. “Chambourcin” (Seyve-Villard 12,417 × Seibel 7053) is a French-American hybrid cultivar commonly grown in Ohio with moderate cold hardiness and can survive temperatures as low as −23°C (Zabadal et al., Citation2007). The vines were planted in 1996 at the Horticultural unit located at The Ohio State University, Wooster, Ohio, USA. There was a total of seven rows of “Chambourcin” vines, each consisting of 60 vines. We used a completely randomized design to select 20 vines (10 controls and 10 K-treated) within two rows. In addition to pruning, shoot thinning was performed on these vines to control yield. This cultivar has the propensity to overcrop, thus cluster thinning is required (Dami, Citation2005). We used a liquid potassium-based fertilizer, ReaXTM (25% K2O) (Loveland Products, Inc.) and applied it to the shoots (leaves and clusters) of the treated vines until they were fully wet. As per the recommendations of the product label, K-treated vines were sprayed four to five times with 1.5% (v/v) of ReaXTM (0.38% K2O) between the fruit set (modified Eichhorn-Lorenz or EL 27) and the veraison stage (modified EL 37) (Dry and Coombe, Citation2004) (). Control vines were not sprayed. The foliar application was scheduled each year on dry sunny days (Temperature = 20–22℃) from late June to late August to ensure maximum nutrient absorption by plant leaves and avoid wash-off due to rainfall
Table 1. Treatment dates and phenology of Chambourcin grapevines in 2021 and 2022.
Weather Data
Air temperature, Growing Degree Days (GDD), and precipitation data were collected from the CFAES Weather System (https://weather.cfaes.osu.edu/). Further, to understand the temperature patterns during the dormant seasons and their effect on grapevine acclimation, we computed mean temperatures for both experimental years and the number of days below freezing before the coldest event each year.
Petiole Nutrient Analysis
To determine the nutritional status of grapevines, petioles were collected during the growing season of 2022. Two hundred petioles were collected at stage EL 36–37 (50% veraison) and then randomly subdivided into four replicates per treatment. Petiole samples were shipped to Brookside Laboratories (New Bremen, OH). The analysis provided a complete determination of petiole macro- and micro-nutrients and is based on published methods (Miller and Horneck, Citation2013).
Yield, Fruit Quality, and Berry K
The crop was harvested on October 19, 2021, and October 25, 2022. The number of clusters per vine (n = 10) were counted and the clusters were weighed to determine yield (kg) per vine. Yield per vine was further multiplied by vine density (2732 vines per hectare with a spacing of 3 m × 1.22 m) to estimate average yield (tons. ha−1). Mean cluster weight (g) was calculated by dividing the yield per vine by the number of clusters per vine. During harvest, 400 berries from each of the 10 K-treated and control vines were randomly collected and divided into four replicates, each consisting of 100 berries placed in a plastic bag. Each of the four plastic bags was then weighed to calculate the 100-berry weight. Following the weight measurement, the berry samples (4 replicates) were taken on ice to the laboratory where they were crushed and the juice was extracted for fruit quality analysis (i.e., total soluble solids [TSS], pH, titratable acidity [TA], and fruit maturity index [FMI]). Thirty five (35) mL of the extracted juice was transferred to 50 mL falcon tubes that were centrifuged at 800 rpm for 8 minutes. Following centrifugation, 10 mL of supernatant was transferred to the titration workstation (Denver Instrument Model 350, Denver, CO) to measure pH and TA. TSS content was measured using a digital refractometer (MISCO, Cleveland, OH) and expressed in degree Brix (°Brix). FMI was calculated as TSS/TA * 10.
During harvest, an additional 200 berries were randomly collected from the K-treatment and control vines for analysis of potassium levels in the fruit. Berries were randomly divided into four replicates, each consisting of 50 berries, placed in plastic bags, transported to the laboratory on ice, and stored at −20°C before being shipped to the ETS Laboratories (St. Helena, CA). After pressing and maceration of the berries, potassium content from the juice was measured using the Flame Atomic Emission Spectroscopy method (Hanson and Horneck, Citation1997) and expressed in mg/L.
Cold Hardiness
The cold hardiness level of the vines was determined using differential thermal analysis (DTA) during both years of the study (Wolf and Cook, Citation1994). Water contains latent heat energy that gets released when water freezes. This latent heat energy can be detected by thermoelectric modules in terms of an increase in voltage (mV), thus, generating Low-Temperature Exotherm (LTE) that denotes the death of dormant bud tissues (Pierquet and Stushnoff, Citation1980). Dormant canes from five randomly selected K-treated and control vines each (one cane per vine) were collected monthly from fall to late winter-early spring during dormancy (2021–2022 and 2022–2023). The buds were excised from the canes and placed on moist filter paper in a petri dish to avoid dehydration. Five excised buds from each cane were placed in each thermoelectric module (TEM) and the module box was placed into a Tenney environmental chamber (Thermal Products Solutions, New Columbia, PA). The Tenney environmental chamber was programmed to gradually decrease the temperature from room temperature (~18.5℃) to −40°C at the rate of 4℃/hour. The median LTE value was used to determine the LT50 (Lethal temperature for 50% bud injury).
The 2022 winter provided the opportunity to evaluate freeze damage in the field when the vines experienced a five-day freeze event which started on December 23, 2022 (minimum temperature dropped from −0.7°C to −21.2°C in 24 hours). To assess bud injury in the field due to this freeze event, canes were collected on January 11, 2023. Canes (node positions 3 to 10) were randomly selected from the K-treated and control vines. In total, five replicates for the K-treated and controls, each consisting of 20 buds, were assessed for injury of primary and secondary buds as the percentage of dead buds out of total buds collected.
Data Analysis
Data analyzes were conducted in R Studio (version 2023.12.0.369). Significance in the comparisons between control and K-treated vines was determined using t-tests, with a significance level set at p ≤ .05. Before conducting the t-tests, normality of the data was evaluated via the Shapiro-Wilk test, and homogeneity variance was examined using Levene’s test. Welch’s t-tests were applied for normally distributed data, while the Wilcoxon rank-sum test was used for non-normally distributed data.
Results
Weather
In the 2021–2022 winter season, the lowest temperature recorded was −21°C on January 27, 2022 (). During the second-year winter season (2022–2023), the vineyard experienced the lowest temperature on December 23, 2022, which was −21.2℃, (, ). The average air temperatures prior to the freeze event of January 27, 2022, gradually decreased and remained below 0°C for at least 15 consecutive days, whereas the temperatures inconsistently fluctuated above or below 0°C before the freeze event on December 23, 2022 ().
Figure 1. Daily maximum (red) and minimum (blue) air temperatures from September 1 to April 30 in: a) 2021–22 and b) 2022–23. “*” denotes the lowest temperatures recorded during the winter seasons of both experimental years (−21°C on January 27, 2022; −21.2°C on December 23, 2022). Air temperature data were not available for Feb 24–28, 2023.
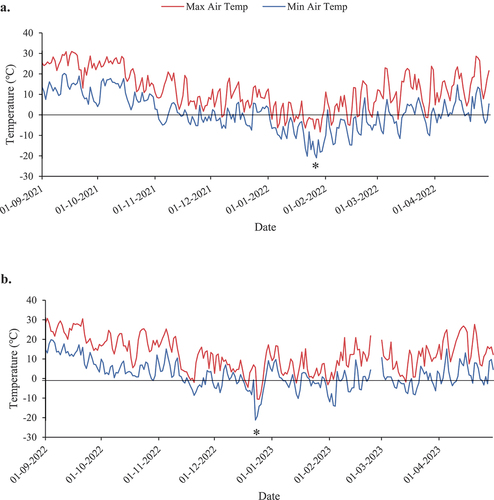
Table 2. Weather parameters during 2021–2022 and 2022–2023 experimental seasons.
Crop yield and fruit quality are influenced by the environmental conditions during the growing season of the crop. For this study, we recorded different weather parameters of the growing season from January 1 to October 31. During the 2021–2022 growing season, the maximum temperature was 34.3°C on June 29, 2021. During the 2022–2023 growing season, the maximum temp was 34.2°C on 15 June 2022 (). During the growing season, cumulative GDD was higher in 2021 (1817) as compared to year 2022 (1689), which was comparable to the average cumulative GDD (1634) of the 30-year average (1991–2020). The fruit was harvested on October 19, 2021, and October 25, 2022; therefore, the cumulative GDD from January 1 until the harvest date was 1795 and 1681, respectively (). On the other hand, the annual rainfall was higher during 2022 (104 cm) than that during 2021 (84 cm), which in turn was lower than the 30-year average (89 cm).
Petiole Nutrient Analysis
Potassium levels were not significantly different between the K-treatment and control petioles (). Among the macronutrients, nitrogen (N), phosphorus (P), and magnesium (Mg) levels were significantly different between the K treatment and controls, with N levels being higher and P and Mg being lower in the petioles of K-treated vines. Sodium (Na) levels in K-treated petioles were 3.5-fold higher than that in controls. All the other nutrient levels were not significantly different between K-treated and control petioles (p-value > .05). Also of note, K, P, boron (B), and iron (Fe) were below the normal range, whereas N, Mg, Manganese (Mn), and Calcium (Ca) were above the normal range for both the control and K-treated petioles (Dami, Citation2005). Actually, the observed petiole K levels (0.73–0.84%) were considered deficient. This was unexpected especially since we applied a foliar K fertilizer multiple times, and we did not observe any leaf symptoms of K deficiency.
Table 3. Petiole nutrients in Chambourcin during the 2022 growing season.
Yield, Fruit Quality, and Berry K
Crop weight per vine averaged 4.97 kg for K-treated and to 5.68 kg for controls in 2021 (). The mean crop weights were lower in 2022, averaging 4.32 kg in K-treated and 4.73 kg in control vines. As expected and due to cluster thinning, yield components were not significantly different between the control and K-treated vines in both years, except for cluster weight in 2021, where the K-treated vines had significantly lower cluster weight compared to the control vines. Generally, yield components were higher in 2021 than in 2022 likely due to heavier clusters and berries in the first year. This also holds true as to why control vines had higher (though non-significant) yields compared to K-treated vines during both years of the experiment.
Table 4. Yield components of Chambourcin for the years 2021 and 2022.
The average sugar content (TSS) for the K-treated vines was 23.3 and 22.4 °Brix for 2021 and 2022, respectively, and was 22.5 and 21.6 °Brix for control vines, so was significantly higher in K-treated vines as compared to the controls in both years (). TSS increased by 4% in K-treated berries in both years. The average pH levels for the control and K-treated samples were not significantly different in 2021 (3.26 and 3.29) or in 2022 (3.21 and 3.20). Fruit TA levels were significantly lower in K-treated vines (8.4 g/L) compared to the controls (9 g/L) in 2021, however, the differences were non-significant in 2022 (11.3 g/L in K-treated and 11.5 g/L in controls). Due to higher TSS and lower TA, the fruit maturity index (FMI) was significantly higher in K-treated vines in 2021.
Table 5. Fruit parameters in Chambourcin for the years 2021 and 2022.
Potassium levels in the berries in 2021 were 925 mg/L in controls and 1017 mg/L in K-treated vines and were not significantly different. However, in 2022, K-treated vines had significantly higher K (1450 mg/L), approximately 20% higher, than the control vines (1212 mg/L) ().
Cold Hardiness
Differential thermal analysis during winter 2021–2022 showed that the mean LT50 values for buds collected on December 20, 2021 were similar for K-treated (−23.6℃) and control vines (−24℃) (). The LT50 values increased a little for bud samples collected on January 3, 2022, and the differences remained non-significant (−23.1°C for controls and −22.2°C for K-treated vines). After the freezing event of January 27, 2022, the buds of K-treated vines collected on February 9 and February 24 during the winter of 2021–2022 had significantly lower LT50 values compared to the controls (p-values = 0.01 and 0.02, respectively). Cold hardiness in K-treated vines increased by 3.3°C and 1.5°C, respectively, compared to the controls. Later in March, LT50 values were not significantly different in buds from K-treated (−18.5℃) and control vines (−18.8℃). The bud LT50 values in both K-treated and control vines reached their lowest values after the vineyard experienced the lowest air temperature of the season, i.e., −21°C during the last week of January 2022. Because the cold hardiness levels (LT50) were maintained below −22°C until the end of February 2022, the extreme air temperatures (−21℃), did not cause bud injury in the field.
Figure 2. Mean LT50 values for K-treated and control buds collected during the dormant seasons of: a) 2021–22, and b) 2022–23. Error bars denote standard errors. “*” denotes a significant difference between treatments at p < .05.
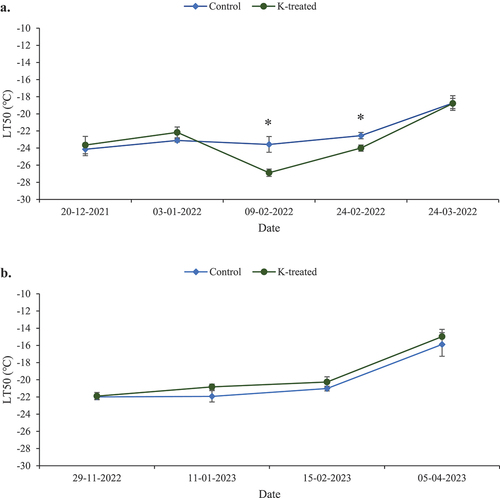
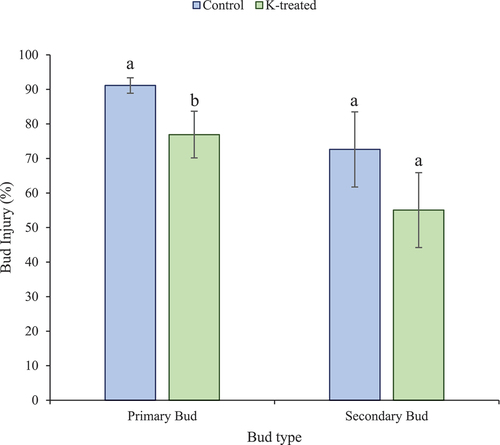
During winter 2022–2023, no significant difference in the LT50 values was observed between the K-treated and control bud samples at any collection date (). LT50 values remained between −20°C and −22°C from late November 2022 until mid-February 2023. Later, the cold-hardiness levels decreased and LT50 values were higher in April 2023, but still non-significantly different between K-treated and control vines (−16°C in controls and −15°C in K-treated canes). After the freeze event (lowest temperature = −21.2℃) on December 23, 2022, buds were collected on January 11, 2023, for injury assessment. Primary bud injury was significantly lower for K-treated vines (77%) than the controls (91%) (p-value = .05). Secondary bud injury (55% for K-treated and 73% for controls) was less than that of primary buds, but not significantly different between controls and K-treated vines (p-value = .13) (). Due to a significantly high level of primary bud injury from the December 2022 freezing event, the LT50 values for January, February, and April of 2023 were not reliable, since it is likely that buds were already dead before the DTA tests were performed.
Figure 3. Percent bud injury for K-treated (green) and control (blue) buds collected on January 11, 2023. Mean values marked with different letters denote a significant difference at p < .05. Error bars denote standard errors.
Discussion
In this study, liquid-based potassium fertilizer, ReaXTM (0-0-25), was applied to the leaves of grapevines four to five times at a concentration of 1.5% (v/v) during the growing seasons of two consecutive years. ReaXTM contains 25% K2O, therefore, the total K2O (%) applied to the plants with five foliar applications was approximately 1.88% K2O per year or 0.38% K2O per application. The rate of 1.5% (v/v) was selected because it corresponded to the highest rate recommended for grapes by the product label as higher concentrations would lead to phytotoxicity (Centinari et al., Citation2016; Wilson, Citation2001).
Previous studies tested the effects of various concentrations of potassium on grapevine cold hardiness (Centinari et al., Citation2016, Citation2018; Sarikhani et al., Citation2014). Sarikhani et al. (Citation2014) applied 0%, 0.25%, 0.5%, and 1% K2O (w/v) in each of five foliar applications on “Bidaneh Sefid” grapevine (Vitis vinifera L.). They found a continuous increase in bud and cane cold hardiness, proline, soluble sugars, and protein concentrations with higher K concentrations. Although vines showed some response to the lower concentrations of the fertilizer, application of K2O at 1% (w/v) concentration was considered most effective in increasing the cold hardiness of the canes and the buds. Similarly, Karimi (Citation2017) applied four foliar sprays of K2SO4 using 0%, 0.5%, 1%, and 1.5% K2O (w/v). The study found that a higher concentration of K2SO4 (1%, and 1.5% K2O) profoundly increased cold hardiness, ABA, polyamine, soluble sugars, and phenolic compounds. In another study, foliar macronutrient, Potassium Dextrose-Lac (KDL; 24% K2O) was applied at a concentration of 0.24% K2O just before exposing the vines to either natural or artificial freezing conditions (Centinari et al., Citation2016, Citation2018). The results of those studies showed a reduction in shoot mortality among single-bud cuttings but no effect on whole vines (Centinari et al., Citation2016).
In our study, we used a comparatively low concentration (0.38% K2O per application) and observed a significant increase in TSS, berry K levels, and cold hardiness, but a deficiency in K in the petioles. The fertilizer application showed no significant effect on yield or the other fruit quality traits measured. Additional studies are needed to determine if ReaXTM has an effect on protein, hormone, and secondary metabolite quantity, as was reported in other studies (Karimi, Citation2017; Sarikhani et al., Citation2014), or if increasing the concentration > 1% K2O per application could further affect fruit quality and cold hardiness.
Potassium is the most abundant cation in plant tissue and plays a critical role in overall plant health (Mpelasoka et al., Citation2003). Potassium is necessary for cellular osmoregulation, electrochemical processes, enzyme activation, cell division, photosynthesis, and protein and carbohydrate synthesis and transportation (Centinari, Citation2016; Karimi et al., Citation2014). Inadequate amounts of potassium can lead to reduced shoot, root, and fruit growth; thus, it is vital for growers to monitor potassium (Centinari, Citation2016). Demand for potassium is highest from fruit set through ripening (Keller, Citation2015; Mpelasoka et al., Citation2003), therefore, foliar application was applied during these stages of development (Rogiers et al., Citation2017). In our study, although we observed a 15% increase in potassium levels in K-treated petioles, the difference between K-treated vines and controls was not significant and all the vines showed deficient petiole K levels (0.73 and 0.84%), even after multiple foliar applications. For the Eastern US, petiole concentrations of potassium less than 1% are considered inadequate, whereas the normal range for potassium concentration in grapevine petioles is 1.5–2.5% (Dami et al., Citation2005; Wolf, Citation2008). When vines are K deficient, soil application, instead of foliar application, of the fertilizer is warranted due to the large amount of K needed (Wolf, Citation2008). The root system of grapevines has a larger surface area for absorbing nutrients and K that is not immediately taken up can remain in the soil to be absorbed as needed. In addition, higher concentrations of K fertilizer can be used since the risk of phototoxicity is low when soil fertilizers are applied at proper concentrations.
According to Mpelasoka et al. (Citation2003), the relation between petiole K and berry K content is still questionable. Similar results for N levels were found by Davenport et al. (Citation2012) where petiole N levels were below the normal range, even though the crop produced was of high quality for premium wine production. In our study, even though the nutrient analysis showed K deficiency in petioles of K-treated and control vines, the K levels in the berries were around or above the optimum level (>1000 mg/L), with the berries from K-treated vines having greater K levels than controls. K within the plants was likely transferred from their source locations to the strongest sinks, i.e., developing and ripening berries, leaving other tissues, such as the petioles, deficient. The K applied to the leaves of the treatment plants was also moved from source to sink, resulting in the higher K detected in the K-treated berries compared to controls. Buds are also nutrient sinks and K could be deposited within the buds, as well, enhancing the cold hardiness, though testing the potassium levels of the bud tissue between K-treated and control vines is needed to validate this hypothesis. Further investigation of K fertilizer application timing (e.g. post-harvest) and the transportation and sequestration of potassium to different plant tissues can better determine its value as a management practice for increasing cold hardiness.
It is also noted that the accumulation of sodium was approximately 3.5 times higher in K-treated vines than in the controls. We hypothesize that this could be due to a gradient effect of potassium-sodium exchange. Potassium was in higher demand in the sink tissues, i.e., the berries, therefore, sodium could have substituted for potassium as an osmoticum in the petioles. The concentration of sodium ions increases in berry pulp and skin along with potassium ions, but at a lower level. Under a limited K supply, sodium, along with magnesium and calcium, can take the role of an inorganic osmoticum replacing potassium ions (Keller, Citation2015). This could be the reason why we observed higher sodium levels in the petioles. In our study, Mg and Ca accumulation was lower in K-treated vines as compared to the controls, which could have further elevated the sodium levels to raise the water potential of the cell.
Although the cluster weight of K-treated vines was significantly lower in the first year, in general, the application of potassium fertilizer did not affect the yield components of Chambourcin. This finding is consistent with that of Centinari et al. (Citation2018) where they found no significant effect of KDL foliar spray on yield of Vitis vinifera cultivars Lemberger and Riesling and interspecific hybrid cultivars Noiret and Traminette. Potassium silicate (K2SiO3) foliar application did not affect berry weight or yield in Touriga National and Touriga Franca grapes, either (Singh et al., Citation2020). However, Karimi (Citation2017):did find an increase in cluster weight and crop weight per vine in “Sultana” grapevine (Vitis vinifera L.) with increasing concentration of foliar application of K2SO4. Additionally, all concentrations of K2SO4 increased individual berry weight over the controls (Karimi, Citation2017). We did observe a significant effect on cluster weight in 2021, but this did not translate to crop wt. or yield, which was not significantly different between the controls and K-treated vines. The discrepancy observed between K treatments and crop weight across studies may be due to variations in the concentration of K applied or could be a cultivar-dependent response.
However, foliar application of K increased sugar accumulation in berries. This finding is consistent with previous studies (Karimi, Citation2017; Sarikhani et al., Citation2014). Sarikhani et al. (Citation2014) suggested that K+ ions could be involved in inducing the enzymes for carbohydrate metabolism. This has been shown in other crop studies such as potato, where potassium increased the activity of sucrose synthase (Liu et al., Citation2013). An increase of K in grapevine could induce carbohydrate metabolism in the berries, which could lead to the higher sugar content observed. Additionally, high sugar levels in buds have been linked to enhanced cold hardiness, due to their role in osmoregulation (De Rosa et al., Citation2022). The positive correlation between K and sugar accumulation suggests that the K accumulated in buds (a sink tissue) protects against freezing injury by increasing the sugar levels, and therefore decreasing the freezing point of the cells (Cakmak, Citation2005).
Titratable Acidity is the ratio of free organic acids to organic acids neutralized by K+ (Villette et al., Citation2020). Among organic acids, tartaric acid is a stronger acid while malic acid is a weak acid and gets degenerated by malic enzyme (malic acid-degrading enzyme) (Lakso and Kliewer, Citation1975). K+ is known to decrease tartaric acid in juice and wine by combining with it to form potassium bitartrate precipitates, thus, lowering the tartrate-malate ratio (Mpelasoka et al., Citation2003). This could be the reason why we observed a decrease in fruit TA for K-treated vines, though it was only significantly lower in 2021. It is also noted that TA values were generally lower in 2021 than in 2022. This could be attributed to the warmer growing season of the former, since warmer temperatures can affect enzyme activity. The activity of the malic acid-degrading enzyme continuously increases with an increase in temperature from 10°C to 46°C (Lakso and Kliewer, Citation1975), therefore, due to higher temperatures, malic acid gets used as a respiratory substrate resulting in lower TA (Famiani et al., Citation2016; Villette et al., Citation2020).
In our study, K content in berries was 1017 mg/L in K-treated vines and 925 mg/L in controls in 2021. In 2022, the values were significantly higher in fruits of K-treated vines (1450 mg/L) than in the controls (1212 mg/L). Previous studies state that K levels within the range 860–1279 mg/L are considered “normal” (Somers, Citation1977) and values higher than 1056–2776 mg/L are considered high levels of fruit potassium (Mpelasoka et al., Citation2003; Somers, Citation1975). Although the berry levels were within the optimum range, the values were still low as compared to the K levels found in other red grape cultivars (1700–1800 mg L−) (ETS Laboratories, Citation2022). High concentrations of potassium in the fruit can cause high pH (>3.70) of fruit juice and wine and create changes in the physical, chemical, and microbial stability (Mpelasoka et al., Citation2003), color stability, and oxidative potential of the wine (Gardner, Citation2016). In our study, even though K concentration in the berries of K-treated vines significantly increased in year 2, as compared to the controls, foliar application of K did not affect the juice pH as reported in the literature, which is positive, since a high pH is deleterious for wine quality.
The effect of K on increasing cold hardiness was observed during both dormant seasons. In year 1, LT50 values of the K-treated buds were significantly lower than controls during mid-winter, but not during the acclimation (fall) and deacclimation (late winter-early spring) stages. In year 2, since the DTA test did not work due to bud injury, we relied on determining cold hardiness by tissue browning. Like year 1, K was most effective in increasing cold hardiness during mid-winter in year 2.
During this study, another observation regarding cold hardiness status was worth noting. In general, the cold hardiness of Chambourcin was higher during the 2021–2022 season than that during 2022–2023. Even though the lowest temperature recorded in both dormant seasons was similar (−21 ℃), injury after the second-year freezing event (December 23, 2022) was much higher than the first one (January 27, 2022). Since the coldest temperature in the second dormant season occurred in late December, 2022, the vines likely had not completely acclimated for maximum cold hardiness yet. Plants undergo cold acclimation under low non-freezing temperatures which leads to physiological changes in plants including membrane fluidity, sugar accumulations and cryoprotein synthesis, thus improving their ability to survive freezing conditions (Londo et al., Citation2018). Mild fall/winters, like seen in fall/winter 2022, were not conducive for maximum cold hardiness (Zabadal et al., Citation2007), whereas temperatures below freezing enhance cold hardiness of grapevines (Hamman et al., Citation1996; Zabadal et al., Citation2007). The mean temperature in January, 2022 was below the 30-year average, whereas the mean temperature in December 2022 was above the 30-year average (Supplementary Figure S1). Furthermore, the mean temperature was below 0°C for 7 consecutive days prior to the freeze event of January 27, 2022, while the mean temperature was not below 0°C in the days leading up to the freeze event of December 23, 2022 (). The warmer temperatures proceeding the extreme cold event in December 2022 likely impacted the cold acclimation of the vines prior to the major freeze event that season. Because of this, the majority of primary buds were dead, but our bud injury assessment did find that primary and secondary bud injury in K-treated vines was less than control vines (77% K-treated vs 91% control primary buds injured, and 55% K-treated vs 73% control secondary buds injured) (). These results suggest the K treatment did have a significant effect on preventing bud injury, especially in mid-winter. More studies are needed to understand K transportation in different plant tissues and how it protects dormant buds and canes from cold stress.
Supplemental Material
Download JPEG Image (128.9 KB){kind=link}
Supplemental Material
Download JPEG Image (129.9 KB){kind=link}
Acknowledgments
We would like to acknowledge the assistance provided by Diane Kinney and Megan Soehnlen for maintaining the vines in the vineyard and assisting in lab experiments. We would like to thank Cullen Dixon and Rachel Combs-Giroir for their in-depth review of the manuscript.
Disclosure Statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/15538362.2024.2344558
Additional information
Funding
References
- Cakmak, I. 2005. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 168(4):521–530. doi: 10.1002/jpln.200420485.
- Centinari, M. 2016. Assessing and managing potassium concentration in the Vineyard. Penn State Extension Wine & Grapes U. https://psuwineandgrapes.wordpress.com/2016/09/16/assessing-and-managing-potassium-concentration-in-the-vineyard/.
- Centinari, M., D.M. Gardner, D.E. Smith, and M.S. Smith. 2018. Impact of amigo oil and KDL on grapevine postbudburst freeze damage, yield components, and fruit and wine composition. Am. J. Enol. Vitic. 69(1):77–88. doi: 10.5344/ajev.2017.17030.
- Centinari, M., M.S. Smith, and J.P. Londo. 2016. Assessment of freeze injury of grapevine green tissues in response to cultivars and a cryoprotectant product. HortScience. 51(7):856–860. doi: 10.21273/HORTSCI.51.7.856.
- Dami, I. 2022. Spring frost injury of grapevines and protection methods. OSU Extension Publications. https://extensionpubs.osu.edu/spring-frost-injury-of-grapevines-and-protection-methods/
- Dami, M. 2005. Midwest grape production guide. https://extensionpubs.osu.edu/midwest-grape-production-guide/
- Dami, I., and D. Lewis 2014. 2014 Grape winter damage survey report. https://kb.osu.edu/handle/1811/60447
- Davenport, J., J. Lunden, and T. Winkler. 2012. Wine grape tissue nutrient concentrations in the Inland Pacific Northwest. Commun. Soil Sci. Plan. 43(1–2):21–27. doi: 10.1080/00103624.2012.631395.
- De Rosa, V., R. Falchi, E. Moret, and G. Vizzotto. 2022. Insight into carbohydrate metabolism and signaling in grapevine buds during dormancy progression. Plants 11(8):1027. doi: 10.3390/plants11081027.
- Dry, P., and B. Coombe. 2004. Revised version of “Grapevine growth stages—The modified E-L system” Viticulture 1 – Resources. 2nd ed. Winetitles.
- ETS Laboratories. 2022. Harvest Guide. https://issuu.com/etslabs/docs/harvestguide2022digital
- Famiani, F., D. Farinelli, T. Frioni, A. Palliotti, A. Battistelli, S. Moscatello, and R.P. Walker. 2016. Malate as substrate for catabolism and gluconeogenesis during ripening in the pericarp of different grape cultivars. Biol. Plant. 60(1):155–162. doi: 10.1007/s10535-015-0574-2.
- Gardner, D. 2016, September 23. Making [red] wine from fruit high in potassium. Penn State Extension Wine & Grapes U. https://psuwineandgrapes.wordpress.com/2016/09/23/making-red-wine-from-fruit-high-in-potassium/
- Hamman, R.A., I. Dami, T.M. Walsh, and C. Stushnoff. 1996. Seasonal carbohydrate changes and cold hardiness of chardonnay and riesling grapevines. Am. J. Enol. Vitic. 47(1):31–36. doi: 10.5344/ajev.1996.47.1.31.
- Hanson, D., and D.A. Horneck. 1997. Determination of potassium and sodium by flame emission spectrophotometry. https://api.semanticscholar.org/CorpusID:99517948
- Karimi, R. 2017. Potassium-induced freezing tolerance is associated with endogenous abscisic acid, polyamines and soluble sugars changes in Grapevine. Sci. Hortic. 215:184–194. doi: 10.1016/j.scienta.2016.12.018.
- Karimi, R., A. Ershadi, and M. Esna-Ashari. 2014. Effects of late- season nitrogen and potassium spray on dormant buds cold tolerance of ‘Bidaneh Sefid’ Grapevine. Iran. J. Hortic. Sci. Technol. 15:419–434. Scopus.
- Keller, M. 2015. The science of grapevines. 2nd edition. Elsevier Inc. doi: 10.1016/C2013-0-06797-7.
- Lakso, A.N., and W.M. Kliewer. 1975. The influence of temperature on malic acid metabolism in Grape Berries: I. Enzyme responses. Plant Physiol. 56(3):370–372. doi: 10.1104/pp.56.3.370.
- Lamichhane, J.R. 2021. Rising risks of late-spring frosts in a changing climate. Nat. Clim. Chang. 11(7):554–555. Article 7. doi: 10.1038/s41558-021-01090-x.
- Liu, H., Shi, C., Zhang, H., Wang, Z. and Chai, S. 2013. Effects of potassium on yield, photosynthate distribution, enzymes activity and ABA content in storage roots of sweet potato (Ipomoea batatas Lam.). Aust. J. Crop Sci. 7(6):735–743.
- Londo, J.P., A.P. Kovaleski, and J.A. Lillis. 2018. Divergence in the transcriptional landscape between low temperature and freeze shock in cultivated grapevine (vitis vinifera). Horticul. Res. 5(1):1–14. doi: 10.1038/s41438-018-0020-7.
- Miller, D.R.O., and D.D. Horneck. 2013. Soil, plant and water reference methods for the western region.
- Mpelasoka, B.S., D.P. Schachtman, M.T. Treeby, and M.R. Thomas. 2003. A review of potassium nutrition in grapevines with special emphasis on berry accumulation. Aust. J. Grape Wine Res. 9(3):154–168. doi: 10.1111/j.1755-0238.2003.tb00265.x.
- Pierquet, P., and C. Stushnoff. 1980. Relationship of low temperature exotherms to cold injury in Vitis riparia Michx. Am. J. Enol. Vitic. 31(1):1–6. doi: 10.5344/ajev.1980.31.1.1.
- Poni, S., P. Sabbatini, and A. Palliotti. 2022. Facing spring frost damage in Grapevine: Recent developments and the role of delayed winter pruning – a review. Am. J. Enol. Vitic. 73(4):211–226. doi: 10.5344/ajev.2022.22011.
- Rogiers, S.Y., Z.A. Coetzee, R.R. Walker, A. Deloire, and S.D. Tyerman. 2017. Potassium in the grape (Vitis vinifera L.) Berry: Transport and function. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01629.
- Sarikhani, H., H. Haghi, A. Ershadi, M. Esna-Ashari, and M. Pouya. 2014. Foliar application of potassium sulphate enhances the cold-hardiness of grapevine (Vitis vinifera L.). J. Hortic. Sci. Biotechnol. 89(2):141–146. doi: 10.1080/14620316.2014.11513060.
- Singh, R.K., J. Afonso, M. Nogueira, A.A. Oliveira, F. Cosme, and V. Falco. 2020. Silicates of potassium and aluminium (Kaolin); Comparative foliar mitigation treatments and biochemical insight on grape berry quality in Vitis vinifera L. (cv. Touriga National and Touriga Franca). Biology 9(3):58. doi: 10.3390/biology9030058.
- Somers, T.C. 1975. In search of quality for red wines. Food Technol. Aust. 27:49–56.
- Somers, T.C. 1977. A connection between potassium levels in the harvest and relative quality in Australian red wines. Aust. Wine Brew. Spirit Rev. 24:32–34.
- Villette, J., T. Cuéllar, J.-L. Verdeil, S. Delrot, and I. Gaillard. 2020. Grapevine potassium nutrition and fruit quality in the context of climate change. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00123.
- Wang, H., J. Blakeslee, M. Jones, L. Chapin, and I. Dami. 2020. Exogenous abscisic acid enhances physiological, metabolic, and transcriptional cold acclimation responses in Greenhouse-grown Grapevines. Plant Sci. 293. doi: 10.1016/j.plantsci.2020.110437.
- Wilson, S. 2001, August. Frost management in cool climate vineyards.
- Wolf, T.K. 2008. Wine grape production for Eastern North America. Ithaca, New York: Virginia Polytechnic Institute and State University, NRAES, Cooperative Extension.
- Wolf, T.K., and M.K. Cook. 1994. Cold hardiness of dormant buds of grape cultivars: Comparison of thermal analysis and field survival. HortScience. 29(12):1453–1455. doi: 10.21273/HORTSCI.29.12.1453.
- Zabadal, T., I. Dami, and M. Goffinet. 2007. Winter injury to grapevines and methods of protection. Ext. Bull. E. 2930:7–21.