
Abstract
Expanding diet breadth to previously unexplored resources was a key strategy in Neolithic adaptation through the Holocene. Most prominently, marine resources became substantial supplements in coastal and island regions worldwide. The Neolithic culture in Korea, known as the Chulmun, has been viewed as heavily based on a marine diet, as a large number of sites are shell middens with well-preserved marine resources. Terrestrial animal taxa are also documented in shell middens, but plant resources have been rarely reported because of the absence of archaeobotanical research. To help fill in this gap, we compared archaeobotanical and organic residue isotope data from four Neolithic shell middens, including Beombang, Bibongri, Sejukri, and Tongsamdong (7500 to 4000 cal BP). Results indicate that ca. Coastal inhabitants procured a broad-spectrum of resources from both marine and terrestrial taxa, while maintaining some difference in resource procurement across settlements from the Early Neolithic period. Particularly, the longer the occupation was, the wider the diet breadth became, including various plant resources and terrestrial animals. Our research demonstrates the utility of integrating archaeobotanical, grain impression, and isotopic methods, while building an additional robust case study demonstrating long-term niche construction and diverse resource seeking in coastal regions in the area.
Introduction
Subsistence has been one of the most published research topics in archaeology, as it is a key factor in human evolution and cultural development (Flannery Citation1969; Price and Bar-Yosef Citation2011; Smith Citation2016; Smith and Miller Citation2009; Winterhalder and Goland Citation1997). Studies on Korean Neolithic culture are no exceptions in this regard, but perspectives on the topic have for the most part been narrowly focused (see more discussion in Lee Citation2011). The Neolithic period of Korea (ca. 10,000–3500 cal BP), that is, Chulmun or Jeulmun, is named after the common incised geometric decoration on pottery. Coastal Neolithic subsistence has been regarded as a mixed economy with a focus on marine resources, especially prior to the Middle Neolithic period () (Lee Citation2002; Citation2011a). This view has been particularly dominant given the location of most Neolithic sites on coastlines and islands. The distributional pattern, however, is in part an artifact of survey, which has mostly taken place on coastlines and islands where sites were more visible and mass construction projects were conducted (Pai Citation2016). The dominance of shell middens in these locations is often used as evidence of a heavily marine-oriented economy. While well-preserved shellfish and fish remains are often intensively studied (e.g., Lee Citation2002; Kim Citation2011), in contrast, less than 1% of more than 870 Neolithic sites documented have been subjected to flotation or dry screening, contributing to the perceived invisibility of plant remains. This lack of data collection further reinforces the impression that “plant cultivation played, if any, [a] very limited role” (Kim Citation2006, 170), and is resonant in more recent publications (e.g., Bae, Bae, and Kim Citation2013). Although the Late Neolithic economy is thought to encompass a wide range of foraging strategies (Kim Citation2003, Citation2006; Lee Citation2001), a marine-oriented economy is still interpreted as the “typical coastal lifeway” (Kim Citation2010, 104).
Table 1. Neolithic chronology in the southern coastal regions of Korea.
In a paper published nearly 20 years ago, Crawford and Lee (Citation2003, 94) suggested that Middle and Late Chulmun cultures exhibited few, if any, significant alternations, suggesting that crops helped maintain Chulmun lifeways rather than alter them. This work was based on only two sites, however, as it was the beginning stage of systematic archaeobotanical application in Korea. Crawford and Lee (Citation2003) hypothesized neither a broad-spectrum nor specialized economy of Chulmun culture, but instead suggested that Chulmun farming emerged before the intensification of agriculture. As more data on plant resource use have accumulated in Korea since then (Lee Citation2013, 2016, Citation2017; Lee et al. Citation2011; Lee, Cho, and Obata Citation2019), it is worth revisiting previous notions of Neolithic lifeways in the region.
In order to address the lack of direct studies on Neolithic subsistence, several attempts have been made to trace diet through isotope/trace-element analysis (). Currently, remains of 23 human individuals have been analyzed from six different Neolithic shell middens (Choy, An, and Richards Citation2012; Lee Citation2011a; Shin et al. Citation2013) (). A few bulk isotope analyses have also been mostly confined to shell middens. A marine diet is reflected at all but one site at Daejukri on the west coast, which shows only a plant-based diet (An Citation2006a) (). Three more sites indicate diets based on plants or terrestrial animals in conjunction with marine resource signatures. For example, Gonamri (or Konamri), another west coast shell midden, shows a dietary pattern based on mixed consumption of C3 plants and terrestrial and marine protein (An Citation2006b). Other examples come from the southern coast. The Yeondaedo shell midden, for example, provides evidence of a plant-based diet as a sizable supplement to shellfish and fish during the Early Neolithic period (An Citation2009). Isotope signatures of 10 Early Neolithic individuals buried at the Janghang site on Gadeokdo Island, off Busan Harbor, reflect mainly a marine diet with a possible contribution from terrestrial animal protein and wild plants (Shin et al. Citation2013). The sites of Tongsamdong, Ando, and Daepo show mostly marine signatures (Choy and Richards Citation2010; Choy, An, and Richards Citation2012; Kim Citation2010). Such findings indicate the differentiation of resource use even among those with similar cultural traditions who reside in close proximity to each other.
Figure 1. The locations of archaeological sites mentioned in the text. (A) South coast, showing our sites in red, including Bibongri (1), Sejukri (2), Tongsamdong (3), and Beombang (4) as well as others in blue, including Janghang (5), Daepo (6), Yeondaedo (7), and Ando (8); (B) west coast, showing Dejukri (9) and Gonamri (10). The same scale for both maps. Satellite imagery credits: Esri, DigitalGlobe, GeoEye, Earthstar Geographics, CNES/Airbus DS, USDA, USGS, AeroGRID, IGN, and the GIS User Community.
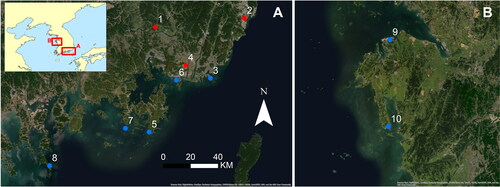
Table 2. Results from bulk isotope/trace-element analysis on Neolithic sites in Korea.
This study, therefore, aims to provide a more comprehensive view of subsistence practices in coastal Neolithic sites in southeastern Korea. Large shell middens at Tongsamdong (also known as Dongsam-dong) and Beombang (or Peombang) have been regarded as model cases of maritime Neolithic subsistence since the 1970s (Ahn Citation1988; Sample Citation1974). For this research, we revisited these two sites using systematic applications of archaeobotany and isotopic analysis on geochemical residues on pottery. We also studied two other shell midden sites, Bibongri and Sejukri, as they were long-term coastal settlements where previous research suggested there was a heavy marine diet (Kim Citation2011) (). A range of 14C dates from the studied sites indicate habitation from the Initial to the Final Neolithic period (7600–3600 cal BP) (, Supplementary Material 1).
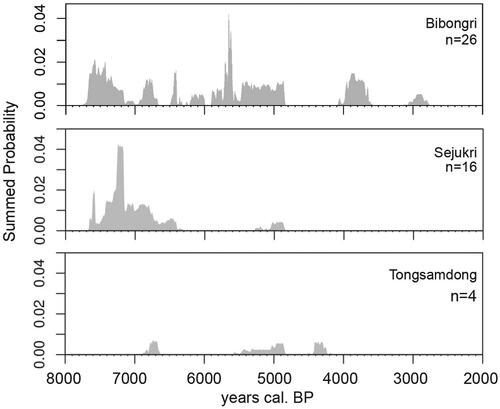
Figure 2. Summed Probability Distributions (SPDs) of 46 AMS dates on charred seeds from Bibongri, Sejukri, and Tongsamdong. Dates were calibrated using OxCal v.3.2 (Ramsey Citation2001), based on IntCal13 atmospheric curve (Reimer et al. Citation2013). SPDs were generated using ‘rcarbon’ R package (Bevan et al. Citation2018) to show the distribution of calibrated dates in each site.
A diachronically diverse setting is ideal to engage niche construction theory to reflect upon how Neolithic people utilized coastal environments to sustain themselves over three millennia. Niche construction is the process whereby organisms actively modify their own and each other’s evolutionary niches (Odling-Smee et al. Citation2013). This theory entails that niche construction be regarded as a fundamental evolutionary process in its own right (Laland, Matthews, and Feldman Citation2016). As it views the multigenerational transmission of learned behaviors and ecological inheritance as key mechanisms in evolution, niche construction theory has been recognized as a new conceptual framework that can integrate the biological and cultural realms of human evolution (Fuentes Citation2015; Laland and O’Brien Citation2010). Ecological inheritance and engineering, the main tenets of niche construction, have been already key concepts in archaeology under the guise of historical ecology (e.g., Bettinger Citation2002; Braje et al. Citation2017; Fitzpatrick and Erlandson Citation2009; Moss et al. Citation2016).
Niche construction theory has been embraced in archaeology since the 2010s. For examples, papers published in the Journal of Archaeological Method and Theory highlighted this trend with a special issue that addressed both hunter-gatherer (e.g., Bleed and Matsui Citation2010; Broughton et al., Citation2010; Riel-Salvatore Citation2010) and agricultural cases (e.g., Broughton et al., Citation2010; Crawford Citation2014; Riel-Salvatore Citation2010). Many other papers have followed suit, highlighting hunting-gathering subsistence (e.g., Bishop, Church, and Rowley-Conwy Citation2015; Zeder Citation2012), domestication, diffusion, and genetic consequences (e.g., Boivin et al. Citation2016; Laland and O’Brien Citation2010; Smith Citation2015, Citation2016; Zeder Citation2016), and agricultural practice (e.g., Aikens and Lee Citation2013; Crawford Citation2011; Lansing and Fox Citation2011; Rowley-Conwy and Layton Citation2011; Smith Citation2011, Citation2016). As these and other papers have noted, the phenomenon of niche construction has likely played a fundamental role throughout human evolution and prehistory (Broughton et al. Citation2010) regardless of the economic practice of individual societies. Island and coastal environments have also long been studied in the scope of historical ecology, which shares key concepts with niche construction theory (e.g., Fitzpatrick, Rick, and Erlandson Citation2015; Levin, Shelton, and Ayres Citation2017). In this paper, we attempt to add an additional case study while avoiding the artificial fault line between hunter-gatherers versus farmers and terrestrial versus marine-oriented economies. Earlier studies on Neolithic Jomon sites in Japan offer a cautionary tale given that the difference between foraging and agriculture is anything but a simplistic division (Crawford Citation2008). Similar to those case studies conducted in Jomon culture, our study evaluates the Chulmun management of landscapes and resources, a core practice of niche construction.
Materials and methods
Coastal, island, and inlet settings
All four sites studied here are situated along coastal areas of South Gyeongsang province, either along the eastern (Sejukri) or the southern (Beombang, Bibongri, and Tongsamdong) coasts (). The western and southern coastlines consist of complicated inlet-and-bay systems and numerous islands, while the eastern coastline is notably straight. Sites on both coasts went through dramatic landscape modifications throughout the Holocene. Tongsamdong is located on an islet of Busan Harbor on the Korean Strait. Beombang used to be located in the inner bay, called “Paleo Gimhae,” prior to dike construction in the 1930s that turned the area into inland terrain (Bokcheon Museum Citation2011). Bibongri currently lies near the tributary of the Nakdong River northeast of Beombang, but sat on the inner bay called “Paleo-Bibong” until the Middle Holocene (Gimhae National Museum Citation2008). Sejukri is right on the east coast, about 45 km north of Busan. Based on 14C dates and pottery types, Bibongri and Tongsamdong are regarded as long-term dwelling sites, spanning from the Initial to Final Neolithic periods (Supplementary Material 1). In contrast, Sejukri was occupied during the Initial and Early Neolithic periods, while Beombang yielded mostly Middle Neolithic seasonal camp materials. The summed probability distributions of 46 dates on terrestrial samples from Bibongri, Sejukri, and Tongsamdong sites indicate the association of our samples from the Initial through the Final Neolithic period (7600–3600 cal BP) ().
Tongsamdong (35° 04′ 15″ N 129° 04′ 46″ E), a designated National Cultural Heritage (Historical Site no. 266), is probably the best studied among all Neolithic sites in Korea. The site was found in the 1930s, but the first excavation was not implemented until three decades later in 1963 (Sample Citation1974). The National Museum of Korea has conducted several field seasons since then, producing five volumes of excavation reports and numerous journal articles throughout the 2000s. Tongsamdong was found on Yeongdo (13.97 km2), an islet currently connected to Busan Harbor by bridge, and is part of the complex system of well-developed inlets, delta plains, and islands. Tongsamdong is directly on the modern coastline (4 m asl), but this was farther away during the Neolithic. The change in sea level was demonstrated by the discovery of artifacts when breakwater construction drained the portion of the bay fronting Tongsamdong (Lee Citation2006). Tongsamdong and other sites in our study exhibit the same cultural tradition as inland sites in the southeastern region, sharing identical pottery styles, house structures, and tool kits.
Lying in the foothills of Mt. Bongraesan, Tongsamdong inhabitants probably had easy access to both terrestrial and marine resources. Despite intense development, Yeongdo still exhibits high biodiversity, accommodating at least 10 terrestrial mammals, 40 avian species, and more than 200 plant taxa (Busan Museum Citation2007). Many of those terrestrial species were also detected in faunal remains at the Tongsamdong site, indicating availability of diverse terrestrial resources throughout the Holocene. Several seasons of excavation during the 1970s revealed large shell middens, reflecting long-term intensive use of the area and the well-defined stratigraphy at Tongsamdong was used in establishing the Neolithic chronology of southern Korea (). In this study, we utilized data from the 1999 field season, which revealed the most comprehensive picture of long-term occupations (Bokcheon Museum Citation2011). Three pit houses and one jar burial were found near the shell midden, along with more than 1,500 shell ornaments, lithics, pottery, and animal bones (). The Tongsamdong site also yielded the first 14C date directly on charred millet grains in Korea (TO-8783), confirming the presence of two millet taxa, foxtail (Setaria italica ssp. italica) and broomcorn (Panicum miliaceum), by the Middle Neolithic period (Crawford and Lee Citation2003) (Supplementary Material 1).
Figure 3. (A) The stratigraphy and site plan of Tongsamdong in the field season in 1999, showing pit houses; (B) sediments for flotation were sampled from the floor of House no. 1 (arrow point) (Bokcheon Museum Citation2011).

Dike construction in the 1930s converted the Beombang wetland to farming fields at the estuary of the Nakdong River. The Neolithic inhabitants faced a quite different environment then, as Beombang was situated along the inner bay, called “Paleo Gimhae,” during the Middle Holocene Hypsithermal Period (Busan Museum Citation2009) (). Along the shoreline of Paleo Gimhae Bay, several Neolithic shell middens were identified in the Beombang District (35° 09′ 22″ N 128° 52′ 52″ E). Two seasons of excavation in 1991 and 2001 revealed an extensive shell midden, several hearths, and a pit grave (Busan Museum Citation2009; Ha Citation1997; Ha, Lee, and Lee Citation1993). Partial remains of one 11- to 14-year-old individual were preserved inside the pit grave. Thousands of artifacts were uncovered, including ornaments and fishing tools made of various shells and stones, pottery, and obsidian tools. Beombang was mostly visited during the Middle Neolithic period from 7000 to 4000 cal BP, as demonstrated by the abundance of Middle Neolithic pottery (Busan Museum Citation2009).
The Sejukri site (35° 27′ 57″ N 129° 21′ 11″ E) represents Initial and Early Neolithic adaptation to a large lagoon environment, dating back to 7500 to 6000 cal BP. The site is currently located on the edge of an inner bay along the east coast, but a lagoon was formed in front of the site by a long stretch of sandbar throughout the Early and Middle Holocene (Hwang and Yoon Citation2002). Among the more than 10 taxa of shellfish and fish identified at Sejukri, species of shallow waters and lagoon were dominant (Dongguk University Museum Citation2008). Various terrestrial mammals were preserved along with marine mammals, including wild boar and deer, as well as migratory birds. Although no dwelling structure was present, such numerous artifacts and bone remains indicate prolonged use of Sejukri.
The Bibongri site (35° 24′ 30″ N 128° 38′ 08″ E) represents long-term occupation from the Early to the Late Neolithic periods (Gimhae National Museum Citation2008) (). Bounded by steep hills (400 m asl) to the north and west, Bibongri is located in the foothills (3.7–3.8 m asl), overlooking the alluvial plain along the Chengdo-cheon River, a tributary of the Nakdong River, to the south. Although Bibongri is located inland today, it sat on a widened bay called “Paleo-Bibong,” where its settlers faced an inner bay environment throughout the Neolithic period. Similar to the case of Sejukri, abundant shellfish and brackish water fish reflect species composition that would be common within bay habitat (Gimhae National Museum Citation2008). Sometime after the abandonment, Bibongri was transformed to a low-lying wetland with the advance of the coastal margin and inundation from the Cheong-cheon. As a result, organic materials were well-preserved in multiple layers of silty mud, shells, and alluvial sediments, including a pine dugout canoe and a paddle, basketry, wooden artifacts, bones, and plants ().
Figure 4. (A) waterlogged, storage pit no. 1 at Bibongri; (B) acorn meat (Quercus sp.) from this storage pit. Scale = 10 mm.

A high pH level of the shell midden preserved faunal remains in excellent condition, including at least 35 taxa (Kaneko 2008). Deer (Cervus nippon hortulorum) and wild boar (Sus scrofa) are the most common taxon in mammal remains, as are in other Neolithic shell midden sites in Korea (Lee Citation2017). Other mammals, fish, and birds were also identified, including bear (Ursus arctos), raccoon dog (Nyctereutes procyonoides), wolf (Canis lupus ssp. lupus), and tiger (Panthera tigris ssp. tigris). Bones of a bovid may represent buffalo (Bubalus bubalis or B. arnee) (Kaneko 2003). Common bird species include water pheasant (Phasianus colchicus ssp. karpowi), duck (Anatidae sp.), redlip mullet (Liza haematocheilus or Planiliza haematocheilus), and Japanese seabass (Lateolabrax japonicus) (Kaneko 2008). Dog (Canis lupus ssp. familiaris) was the only domesticated animal taxon found at Bibongri. Structures at Sejukri, including storage pits, pit houses, and outdoor hearths, indicate that the site was a long-term settlement rather than a seasonal task locality. Below, we describe the recovery and analysis of plant materials found at these sites.
Plant analysis
This paper compares two types of plant data—macroscopic remains and grain impressions on pottery surfaces. To recover plant remains, either column or point sampling was applied to all three sites, strategies for which are described in Pearsall (Citation2015). At the Bibongri and Sejukri sites, bulk samples were collected in well-stratified shell layers and pits, while only a pit house fill was collected at Tongsamdong (). Sediments were soaked with water and floated materials skimmed with a standard geological sieve of 0.212 mm mesh (US Standard no. 70); floated remains were then dried for observation and photographed with a Nikon AZ 100 multi-zoom microscope and a digital DS-U2 camera.
Grain impressions on pottery have been recognized for their potential for tracing plant resources in Korea since the 1970s, but Korean archaeologists have only recently examined these systematically. The most common method is to replicate impressions by injecting silicone into a cavity on the surface of pottery for scanning electron microscopic observation (Obata et al. Citation2011). Our paper reexamines the data on grain impressions previously reported (Obata Citation2013; Obata and Manabe Citation2012) to compare with plant remains recovered from flotation samples.
Isotope analysis
Stable isotope analysis can be an efficient tool for documenting dietary patterns of animals, as the isotopic composition of living organisms depends upon the food they eat (Ambrose and Katzenberg Citation2002). In East Asia, isotope analysis has been applied primarily to human bones with a specific goal to distinguish diets based on rice (C3) from millet (C4) (Barton et al. Citation2009; Bentley et al. Citation2007). As human remains are scarce in low soil pH conditions in Korea, organic residues absorbed within the fabric of clay matrix can be used instead to improve our understanding of human diet (Eerkens Citation2005; Evershed Citation2008a). Lipids—fatty acids, waxes, and sterols—are naturally occurring compounds in organisms and can be preserved inside the porous matrix of fired clay (Reber and Evershed Citation2004). Stable carbon isotope ratios (hereafter δ13C) can be measured from major fatty acids such as C16:0 (palmitic acid) and C18:0 (stearic acid) that are ubiquitous in ceramics (Mottram et al. Citation1999). Fatty acids are carboxylic acids with a carbon chain of various numbers, indicating carbon atoms. C16:0 and C18:0 are two of the most common fatty acids found in nature. They are present in a wide range of food products, including butter, cheese, milk, meat, and sunflower oil.
To control the chronological sequence of our samples, we examined only the type-marker pottery from well-defined strata at Beombang (n = 29) and Tongsamdong (n = 32) where 14C dates are available (Supplementary Material 2). Following Correa-Ascencio and Evershed’s protocol (Correa-Ascencio and Evershed Citation2014), fatty acids were extracted from rims or upper body sherds, and the δ13C vPDB values of C16:0 and C18:0 were measured. Approximately a 5 g portion of each potsherd was sampled, and its surface was then cleaned using a drill (Dremel 3000) to remove any external contaminants, such as those originating from soil or fingers due to handling during the excavation/curation process. No other solvent extraction was conducted other than Correa-Ascencio and Evershed’s protocol. Before initiating isotope analysis, we examined lipids with gas chromatography-mass spectrometry (GC-MS) to separate and identify specific organic compounds within the mixture. Such biomarkers are often used to reconstruct prehistoric diets (Evershed Citation2008a, Citation2008b; Heron and Evershed Citation1993). The prepared fatty acid methyl esters (FAMEs) were measured with the Agilent Technologies 6890 N Network gas chromatography system with a 5979 mass selective detector. The GC was equipped with a fused silica capillary column (J&W; DB5-MS; 60 m × 0.32 mm; 0.25 µm film thickness) and the interface was maintained at 110 °C. The mass spectrometer was operated in full scan mode. Helium was the carrier gas, and the GC oven was programed in the following manner: 2 min isothermal at 50 °C is followed by an increase to 350 °C at a rate of 10 °C min-1, and then the temperature is held at 350 °C for 10 minutes. The peaks were identified based on their mass spectral characteristics and GC retention times and by comparison with the NIST mass spectral library. Isotope analysis was performed using a Thermo GC/C-IRMS system composed of a Trace GC Ultra gas chromatograph (Thermo Electron Corp., Milan, Italy) coupled to a Delta V Advantage isotope ratio mass spectrometer (IRMS) through a GC/C-III interface (Thermo Electron Corp., Bremen, Germany). FAMEs dissolved in hexane were injected in the splitless mode and separated on a Varian factor FOUR VF-5ms column (30 m × 0.25 mm ID, 0.25 micron film thickness). Once separated, FAMEs were quantitatively converted to CO2 in an oxidation reactor at 950 °C. Following water removal through a nafion dryer, CO2 entered the IRMS. Each sample was measured 10 times. Replicate measurements of each sample and FAMEs with δ13C values traceable to international standards (5-α Cholestane) were used to determine the instrument precision (<0.5‰) (Colonese et al. Citation2015).
Results and discussion
Diversity of plant resources
Our data set provides the composition of plants left in dwelling features throughout the Neolithic period in Korea. Floated sediments from Bibongri, Sejukri, and Tongsamdong yielded seeds, fruits, and nuts of 27 taxa in addition to several unidentified types (). Most of the identified taxa are known to provide edible nuts/fruits, medicinal ingredients, or lumber, including oak (Quercus sp.), pine (Pinus sp.), Manchurian walnut (Juglans manchuria), dogwood (Cornus sp.), hop (Humulus sp.), spicebush (Lindera sp.), and Japanese snowbell (Styrax japonicus). In earlier Neolithic contexts, acorn and other edible nuts seem to be a critical seasonal resource. Sejukri and Bibongri shell middens show exceptionally well-preserved storage pits filled with acorns (). At Sejukri, several pits lie slightly below the current sea level at −1.5 to −0.5 m asl. (Dongguk University Museum Citation2008). At Bibongri, a total of 17 pits yielded acorns, wild walnuts, and other plant remains (Lee Citation2008b). Frequent hybridizations of oak species prevent identification down to species level, but distinctive shapes of nutmeat and scales of nutshells may indicate at least three varieties, including Q. aliena, Q. serrata, and Q. variata.
Table 3. Plant taxa identified in Bibongri, Beombang, Sejukri, and Tongsamdong sites.
Inhabitants at Bibongri and Sejukri used brine for soaking tannins out of acorns by storing them in pits dug below the high tide level during the Initial and Early Neolithic periods. Manchurian walnut is another edible nut species found in such pits. Palynological studies indicate an increase in nut-bearing, deciduous broad-leaf trees from the Early Holocene in the region (Hwang and Yoon Citation2005). This resource-rich landscape may have been attractive to early settlers at Bibongri and Sejukri. The lack of nuts in the later Neolithic periods probably resulted from a preservation problem rather than an actual abandonment of nut harvesting practice. That is, the later period samples were recovered from open features, while all the earlier samples with nut remains were from waterlogged pits. Even in the earlier contexts, nuts were not present in open features ().
Table 4. Counts of plant taxa found in the sites studied.
Fifty native species of edible tubers grow in Korea, some of which were likely part of the Neolithic diet (Kim Citation2014). Archaeological evidence of tubers is rare, partially due to poor preservation of parenchyma tissues and the lack of attempts to recover starch and phytolith remains. The only known archaeological case is wild onion bulb encrustation on Early Neolithic pottery at the Bibongri site (Lee Citation2011).
Another critical finding in our study is charred foxtail millet grains systematically recovered from the Early Neolithic context at Bibongri. Through soil flotation, two foxtail millet grains were identified from Shell Layer 1 and Outdoor Hearth no. 10 (). Although a direct AMS date is not possible on such a small amount of carbon in a few millet grains (less than 1 mg), a study on grain impression provides supporting evidence of domesticated millets in this early context. Four foxtail and three broomcorn millet impressions were identified from typical Early Neolithic pottery at Bibongri (Obata Citation2013) (). All vessels with millet impressions were recovered from Shell Layer 1 except for one (BBR 0001) from House no. 2. Three AMS dates on charcoal are available from Shell Layer 1 (Supplementary Material 1). Among them, SNU05-343 (5330 ± 40 uncal BP) was dated on charcoal recovered during the first field season in 2004 (Gimhae National Museum Citation2008), while the other two were collected during the second field season in 2010 (SNU10-1098, 5530 ± 50 uncal BP and SNU10-1099, 5270 ± 50 uncal BP) (Gimhae National Museum Citation2020). All dates indicate the Early Neolithic period, ranging from 6400 to 6290 cal BP (SNU10-1098) to 6180 to 5950 cal BP (SNU10-1099, 1 sigma ranges) (Supplementary Material 1).
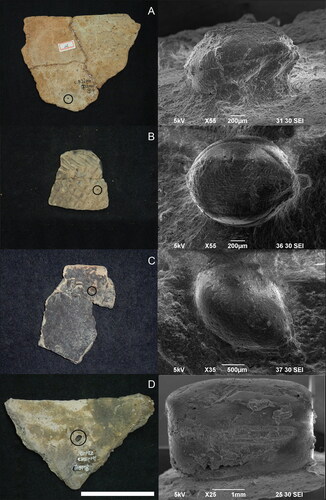
Figure 5. Charred seed remains of edible, anthropogenic taxa. (A to C) Foxtail millet from Bibongri (A, B) and Tongsamdong; (D, E) knotweed; (F) wild kiwi; (G) bramble; (H, I) wild grape. All except millets were retrieved from Sejukri. Scale, 1 mm for foxtail millet, 2 mm for knotweed, and 4 mm for others.

Figure 6. Millet impressions on Early Neolithic pottery recovered from Bibongri (BBR, 6A to 6D). All Bibongri potsherds were collected from Shell Layer 1 except for ‘A’ from House no. 2. Circles on potsherds indicate where each grain was detected. Scale = 5 cm for A, B, and C and 10 cm for D. (A) Foxtail millet impressions on rim sherd, BBR 0001; (B) foxtail millet impression on body sherd, BBR 0017; (C) Broomcorn millet impression on rim sherd, BBR 0018; (D) Azuki impression on rim sherd, BBR 0002.
Early evidence of both millet taxa was also reported from Tongsamdong and Beombang (Obata Citation2013; Obata and Manabe Citation2012). A broomcorn millet impression was reported from the Yunggimun pottery, a typical Initial Neolithic pottery (Obata Citation2013), but Kim (Citation2016) raised the question of whether this pottery is actually Middle Neolithic pottery of the Sugari type. Uncontroversial evidence of foxtail millet came from the Early Neolithic Yeongseondong type pottery from Layer V at Tongsamdong (Obata Citation2013). A total of 18 foxtail millet impressions were detected from four sherds (). Plain pottery at Beombang of the Initial and Early Neolithic layers show impressions of both foxtail and broomcorn millets (Obata Citation2013) (). Samples examined at Beombang, however, are body sherds without diagnostic decoration, so we cannot confirm their Initial or Early Neolithic association until the direct dates on sampled potsherds are available. In brief, two data sets of charred and impression studies from Bibongri and Tongsamdong indicate that millet was used for food and pottery temper at least from the Early Neolithic period possibly as early as 6400 to 5950 cal BP.
Figure 7. Foxtail millet impressions (no. 1–18) on three Early Neolithic rim sherds and one body sherd from Layer 5 at Tongsamdong (DSD0014). Circles on potsherds indicate where each grain was detected and match the numbers on SEM images. Scale = 5 cm.

A more substantive number of charred millets were identified in the dwelling context at Tongsamdong (, ) (Crawford and Lee Citation2003). More than 100 charred foxtail and broomcorn millet grains were recovered from a burned area inside the Middle Neolithic pit House no. 1 (). A direct AMS date on foxtail millet in this feature shows its Middle Neolithic association, dating to 5460 to 5050 cal BP (TO-8383) (Supplementary Material 1). This finding confirms that domesticated millets became a vital resource by the Middle Neolithic period (Lee Citation2011, Citation2016). By 5500 to 5000 cal BP, foxtail and broomcorn millets were widely spread in Korea (Lee Citation2011). All sites subject to archaeobotanical inquiry had examples of either one or both domesticated millet taxa (Lee Citation2017). Raised fields were constructed at the Munamri site along the east coast (National Research Institute of Cultural Heritage Citation2013), indicating the construction of a farming niche (Aikens and Lee Citation2013). Both charred seeds and grain impression on pottery at Munamri indicate widespread use of domesticated millets by the Middle Neolithic period across the Korean Peninsula (Lee, Cho, and Obata Citation2019). This earlier study also provides clear evidence for millet farming in three other Middle Neolithic sites in close proximity to Munamri, including Osanri C, Jikyeongri, and Songjeonri. One grain each of broomcorn millet and rice were recovered from Outdoor Hearth no. 10 at Munamri (Lee, Cho, and Obata Citation2019). No diagnostic pottery was recovered from this hearth, but two AMS dates on charcoal from this hearth indicate its association with the Early Neolithic period, ranging from 6100 to 5940 cal BP (SNU12R001, 5220 ± 50 uncal BP) to 5980 to 5870 cal BP (SNU12R200, 5170 ± 50 uncal BP). Domesticated millets were detected in all open conditions in our study area, either as charred seeds or grain impressions, from the Early to the Final Neolithic periods (). This outcome further confirms that millet has been an integral part of subsistence since the Early Neolithic period along the south coast.
Beyond millets, two managed plant taxa were detected in the Early Neolithic context at Bibongri (Obata Citation2013). Azuki impressions were identified on two typical Early Neolithic type pottery specimens, that is, plain vessels and those with a top-notch decoration from House no. 2 (). Morphological study on charred azuki grains indicates that small sized azuki in Neolithic contexts may represent managed species (Lee Citation2016). Further north along the east coast, a possibility of even earlier azuki in the Initial Neolithic context was also suggested based on a grain impression study (Lee, Cho, and Obata Citation2019).
Preservation issues
Our data show differential preservation of taxa (). Several taxa, including wetland species, tuber fragments, and fruits/nuts of herbaceous shrub and tree species, were detected only in wetland samples. Many of these species are economic taxa either for consumption or medicinal use (Lee Citation2014). This pattern indicates that some wild plant resources are probably underrepresented in open sites, which compose the majority of Neolithic sites in Korea. The absence of plant remains often leads to the impression that plant procurement during the Neolithic period did not contribute to subsistence in any meaningful way.
In both preservation conditions, when rigorous soil sampling and flotation procedures were applied, upland herbaceous plants and shrubs were frequently found. They include ruderal species—chenopod (Chenopodium sp.), knotweed (Polygonum sp.), millet tribe taxa (Paniceae)—and edible wild fruits such as kiwi (Actinidia sp.), grape (Vitis sp.), and bramble (Rubus sp.). These taxa seem to take advantage of human-induced disturbance, particularly the creation of open canopies and waste-driven increases in soil nutrients. They are common near settlements and farming fields even today and have been traditionally used as dietary supplements or medicine (Lee Citation2014) and were probably part of Neolithic plant resource procurement in an anthropogenic environment. By the Middle Neolithic period, millet was established as a key local resource in several regions (Lee Citation2011). The abundance of ruderal species at Tongsamdong seems to represent disturbance from longer occupation and millet cultivation. In wetland conditions, Initial Neolithic samples from Sejukri yielded abundant upland weeds, comparable to Tongsamdong.
Interestingly, Sejukri samples show two distinctive morphologies of knotweed achenes (), including tubercle-shaped and smooth-surface ones, similar to findings from the Late Archaic to Woodland periods in the American Bottom, Illinois, Mississippi, and Ohio Valleys (Mueller Citation2017). As we have only a few specimens, we cannot yet conclude that a smooth morphology indicates human management. Along with knotweed, other weedy taxa in our samples reflect plant resource procurement in an anthropogenic environment.
Differences in taxa composition may have been affected partially by preservation bias (wetland versus open conditions) and different analytical applications. The absence of crops at some sites may stem from a lack of rigorous soil sampling and flotation. For example, foxtail millet grains were found only in samples subjected to flotation at Bibongri and Tongsamdong. Small seeds were absent or rare from waterlogged features at Bibongri, probably because only visible large plant parts (tree nuts and shrub/tree fruits) were hand-picked without soil sampling. In contrast to the rarity of smaller seeds at Bibongri, wetland features at Sejukri yielded numerous tiny upland weedy seeds through systematic soil sampling and flotation. It is also premature to conclude that there were no crops at Sejukri without observation of grain impressions on pottery.
Neolithic waterlogged samples contained four woody taxa that were also present in the Iron Age Shingchangdong wetland site on the west coast (ca. 1800 BP) (Lee Citation2003). Among them, the Japanese hop grows in riverbanks and floodplains, while the remaining three are upland taxa (). This indicates that settlers in lowlands intentionally brought upland resources to residential areas. For example, the fruit of the Japanese snowbell is known to be used for fishing, as it contains a chemical compound that can paralyze fish (Lee Citation2014). Both the Japanese hop and the Japanese dogwood are known for the medicinal properties of their fruits and leaves (Yi Citation2014). This combination of taxa, visible only in wetland sites, represents purposeful harvest of upland plants rather than accidental inclusion of nearby lowland vegetations. In brief, our archaeobotanical data reflect that Neolithic dwellers utilized anthropogenic flora from both disturbed uplands and lowlands.
What was cooked in pots?
Our isotope analysis provides us a glimpse of what Neolithic people cooked in their ceramic vessels. Out of 29 sherds examined at Tongsamdong, 13 contained lipid residues with a mean of 25,740 ng/g (). In the case of Beombang, eight out of 32 samples contained lipids to be further tested (mean of 55,150 ng/g). Some of the samples had to be omitted mainly due to a low concentration of lipids.
Table 5. The concentrations of lipids in pottery samples analyzed for this study.
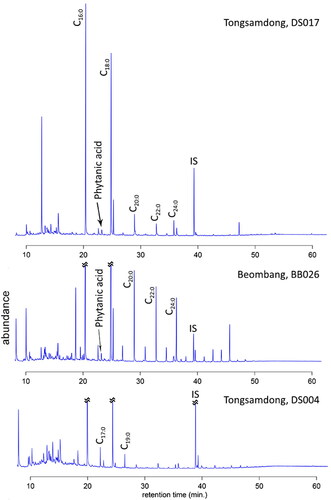
We were able to detect major odd- and even-numbered saturated fatty acids from the GC-MS analysis (). Peaks in the GC-MS chromatogram indicate organic compounds, including fatty acids, with a different retention time along the horizontal axis. The lipid distributions of our samples were dominated by palmitic (C16:0) and stearic (C18:0) fatty acids (Supplementary Material 3). Other organic compounds from the original contents of the vessels were probably degraded significantly during the post-depositional process, leaving these two prevalent acids in organisms more visible in our samples (see of Evershed Citation2008a). A high concentration of C18:0 fatty acid indicates an animal source (Copley et al. Citation2005; Enser Citation1991; Evershed et al. Citation2002). In addition to that, three samples (DS004, 025, 029), all from Tongsamdong, displayed a higher abundance of odd carbon number fatty acids (e.g., C15:0, C17:0, C19:0) (). Previous research indicates that these odd carbon number fatty acids tend to be more enriched in ruminant animals than others (Evershed et al. Citation2002). Due to many uncertainties, however, we do not believe that this alone is a suitable indicator. For example, differential degradation of compounds during the post-depositional process can alter the original compositions (Evershed Citation2008a). Marine resource use is reflected in the presence of aquatic biomarkers in the lipid analysis, including phytanic acid (3,7,11,15-tetramethylhexadecanoic acid), 4, 8, 12-trimethyltridecanoic acid (TMTD), and thermally produced long-chain ω-o-alkylphenyl alkanoic acids (e.g., Craig et al. Citation2011, Citation2013). All but four samples yielded phytanic acid (). Phytanic acid alone cannot be a reliable indicator of marine diets without other aquatic biomarkers, as this can be traced in the tissues of terrestrial animals, including ruminants (Heron and Craig Citation2015). Recently, Lucquin, Colonese et al. (Citation2016) traced the origin of phytanic acid by measuring the ratio of 3S,7R,11R,15-phytanic acid (SRR) and 3 R,7R,11R,15-phytanic acid (RRR), although this method was not applied in our study.
Figure 8. Chromatograms results from the GC-MS analysis. 5-α Cholestane was added as an internal standard (1232 ng). *IS: internal standard. (A) Tongsamdong sample, DS017; (B) Beombang sample, BB026 from Beombang; (C) Tongsamdong sample, DS004.
For the interpretation of isotope analysis, we adopt the approach that utilizes the Δ13C (δ13C18:0 - δ13C16:0) proxy (Salque et al. Citation2013). This method removes exogenous factors related to the local environment and thus highlights only the metabolic and biosynthetic characteristics of the animal fat sources themselves. As a result, it is possible to discern non-ruminant fats (both terrestrial and marine animals), ruminant fats, and ruminant milk fats (Dunne et al. Citation2012; Salque et al. Citation2013). Based on this method (Salque et al. Citation2013), we were able to trace three ruminant fats out of 13 samples from Tongsamdong (DS004, 025, 029) (). All of the other 10 samples show non-ruminant animal fats (DS001, 003, 008, 013, 015, 017, 018, 019, 022, 028).
Figure 9. Results of isotope analysis of Tongsamdong (TSD) and Beombang (BB) samples, using the method described in Salque et al. (Citation2013). Y axis indicates Δ13C18:0 = δ13C18:0 - δ13C16:0.
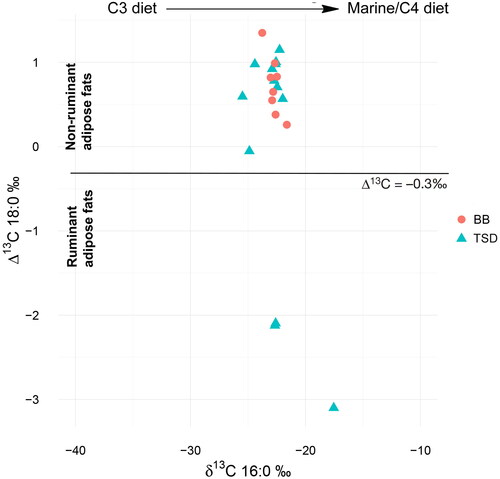
During the 1999 field season, a total 13,185 identifiable animal bones were recovered from Tongsamdong. A considerable representation of terrestrial mammal bones implies that Neolithic settlers consumed terrestrial game on a regular basis (Nishimoto, Kim, and Niimi Citation2011). Among them, approximately 38% were terrestrial animals and most of them were identified as either Sika deer (Cervus nippon) or Siberian roe deer (Capreolus pygargus). Wild boar (Sus scrofa) was also frequently detected. A similar composition has been observed from other analyzed Neolithic shell midden sites in Korea, including Bibongri and Sejukri (Choy, An, and Richards Citation2012; Lee Citation2011b; Shin et al. Citation2013).
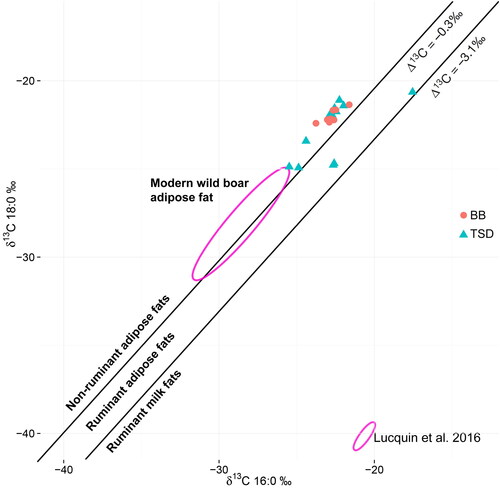
Considering the abundance of wild boar bone remains at Tongsamdong, we compared non-ruminant fats from 10 Tongsamdong samples to the ellipse of adipose fat from archaeological and modern Japanese wild boar (Lucquin, Gibbs et al. Citation2016). At least two (DS013, 015) were close to the range of this boar species (). As a diet based on local vegetations can result in different δ13C values even in the same consumer species, the direct analogy may lead to an incorrect identification of the source. Although it is known that C3 plants are dominant, especially in northeastern Japan (Still et al. Citation2003; Takeda and Hakoyama Citation1985; Takeda et al. Citation1985), it remains a question whether we can identify the δ13C values of archaeological samples from Korea solely based on Japanese references. For that reason, identification of wild boar fat in our samples remains conjectural. The remaining eight non-ruminant samples (DS001, 003, 008, 017, 018, 019, 022, 028) show relatively high δ13C C16:0 values ().
Figure 10. The distribution of δ13C18:0 and δ13C16:0 values of Tongsamdong (TSD) and Beombang (BB) samples, indicating a difference in subsistence strategy.
Ruminant adipose fats traced from three Tongsamdong samples (DS004, 025, 029) probably originated from Sika deer or Siberian roe deer. All three samples show high δ13C C16:0 values. This result contrasts with low δ13C C16:0 values in the ruminant animals from inland Bronze Age settlements with a clear signature of C3 plant consumption (Kwak, Kim, and Lee Citation2017; Takeda and Hakoyama Citation1985). Generally, an increase in the δ13C C16:0 values implies an increase in either C4 plants or marine resource consumption (Salque et al. Citation2013). The contribution of marine plants to ruminant diets has also been observed in other coastal regions. For example, the diet of sheep in Orkney in Great Britain is dominated by seaweed with only a small quantity of seasonal terrestrial grass (Schulting et al. Citation2017). As a result, the bulk δ13C values in their bone collagen (−13%) are close to those of total marine consumers (Schulting et al. Citation2017; Ambers Citation1990). In this respect, high δ13C (C16:0) values in ruminant fats at Tongsamdong may reflect the consumption of marine plants.
The coastal location of Tongsamdong gives strength to the interpretation that non-ruminant animal fats with relatively high δ13C C16:0 values reflect marine resources. Two cases have been published. From two coastal sites at Sejukri and Jukbeyonri, Shoda et al. (Citation2017) traced animal fats with high δ13C C16:0 values from Neolithic pottery, which they suggested was the result of marine consumption. Bulk isotope analysis on dog at Bibongri provides another example of high δ13C values that was interpreted as evidence of a marine diet (Kim Citation2012). Thus, eight non-ruminant Tongsamdong samples with high δ13C C16:0 values may indicate marine resource utilization.
This interpretation, however, may not be straightforward in agricultural settings, such as Tongsamdong. Our study confirms that millet farming was integrated into local subsistence at Tongsamdong by the Middle Neolithic period after millet adoption by the Early Neolithic. Farming villages often feed their livestock with crops or crop byproducts. For example, in northern Chinese Neolithic sites, high δ13C C16:0 values in pig have been interpreted as millet consumption (Pechenkina et al. Citation2005). In such farming communities, not just livestock but also wild terrestrial animals are often adapted to grazing crops as rural pests. In fact, deer and wild boar are recognized as one of the greatest pest threats for modern-day farmers as they consume and damage crops (Davis et al. Citation2016; Lee and Lee Citation2019). As Tongsamdong provides convincing evidence of firmly established millet farming, we raise the possibility that at least some of the ruminant and non-ruminant samples with high δ13C C16:0 values could be related to grazing millet.
In conclusion, both faunal and isotope data consistently reflect the regular consumption of terrestrial mammals in addition to marine resources. Roughly 38% of our Tongsamdong samples (DS004, 025, 029, 013, 015) indicate terrestrial mammal fats. Since some of the non-ruminant samples with high δ13C (C16:0) values may reflect terrestrial animals that consumed C4 plants, the proportion of terrestrial mammal fats could be even higher than 38%. Tongsamdong samples provide evidence of cooking terrestrial mammals, such as Sika deer, Siberian roe deer, and possibly wild boar. Millet farming may have attracted these mammals, which is reflected in their high δ13C values. In contrast to Tongsamdong, Beombang samples indicate non-ruminant animal fats with far less variation in δ13C values ( and ). Beombang samples most likely represent marine resources.
Conclusions
Interpretations emphasizing the marine diet in Neolithic culture have long been mainstream in Korean archaeology. This argument is based on the distributional patterns of most Neolithic sites in coastal regions and abundance of marine remains there. This study explored the multiple lines of data to gain a more nuanced view on diet in the southeastern coastal region without an assumption based on which is immediately visible at the sites. To do this, we applied archaeobotanical methods on charred remains and grain impressions on pottery along with isotope analysis on lipids preserved in the clay matrix of pottery.
Our study sites reflect Neolithic settlements and lifeway choices. All sites were located on seaside landscapes, including an inner bay, an estuary, a lagoon, and an island coast during the Neolithic period. For example, Tongsamdong inhabitants faced a more open sea environment, while Sejukri people inhabited the protected environment of an inner bay and a lagoon. Similarly, Bibongri people settled at the end of a long inner bay, avoiding the direct influence of ocean storms. Beombang settlers were able to utilize diverse estuary resources. Hills or mountains are accessible from all sites and probably provided a good vantage point to access terrestrial resources. Indeed, shell middens show diverse terrestrial mammals and birds of 20 taxa along with fish and shellfish (Lee Citation2011). Our isotope results on Beombang samples provide evidence of reliance on marine resources. On the other hand, Tongsamdong indicates consumption of terrestrial animal fats on a regular basis, most likely those of Sika deer, Siberian roe deer, and possibly wild boar. Neolithic inhabitants built settlements at Tongsamdong, in contrast to Beombang which was only visited seasonally. Differences in isotope composition between the two sites reflect the greater investment of Tongsamdong inhabitants in diverse local resources in order to prolong the duration of settlements. Deer and wild boars are the two dominant taxa among animal bones in this study area (Nishimoto, Kim, and Niimi Citation2011). Our isotope analysis on Tongsamdong pottery also supports this observation.
Moreover, our archaeobotanical work indicates the sustaining importance of terrestrial resources at Tongsamdong and other sites from the Early Neolithic onward. The presence of foxtail millet in floated samples from the Early Neolithic context at Bibongri is consistent with the finding from grain impression analysis there and from Tongsamdong. A total of 25 grains of both foxtail and broomcorn millets were impressed on the Early Neolithic pottery from Tongsamdong and Bibongri. Millet impressions were found in Shell Layer 1 at Bibongri with 14C dates ranging from 6400 to 6290 cal BP to 6180 to 5950 cal BP (SNU05-343, SNU10-1098, SNU 10-1099) (Supplementary Material 1). By the Middle Neolithic period, domesticated millets became vital foodstuffs at Tongsamdong (). Increasing millet cultivation by the Middle Neolithic is also well confirmed in other regions, including the east coast (Lee, Cho, and Obata Citation2019) and central inland regions (Lee 2016). The existence of non-native domesticated millets demonstrates the long-distance network by the beginning of the Early Neolithic period (6500/6000 cal BP). High δ13C C16:0 values in non-ruminant fats at Tongsamdong may reflect either marine resources or C4 plants consumed by terrestrial animals. In coastal regions, the former interpretation is more common, but the presence of millet at Tongsamdong indicates a possibility of the latter case. Neolithic millet fields may have also attracted wild animals, functioning as “hunting gardens,” similar to traditional subsistence farming in Panama (Linares Citation1976; Smith Citation2005). This early farming niche would have facilitated bringing both carbohydrate and protein sources to or near the settlements. Our archaeobotanical study also detected that weedy species common in agricultural settings became prevalent as early as the Initial Neolithic period. Diverse resource use and management of terrestrial plants and animals were an integral part of subsistence through the Neolithic period, even before the adoption of millet farming. For example, wild economic herbs, shrubs, and trees are well detected in our flotation samples from the Initial and Early Sejurki and Bibongri sites. By the Early Neolithic period, domesticated millet was part of subsistence and craft production. This fits into the global trend of niche building based on procurement of broad-spectrum resources from the onset of the Holocene (Zeder Citation2012). Distinctive seasonal environments in Korea provide highly seasonal resources, and our data show that Neolithic people sought after diverse resources throughout the year. Coastal inhabitants built up the cultural niche based on seasonally available terrestrial and marine resources as well as millets from the Early Neolithic period. Neolithic people constructed a long-term niche in coastal environments that facilitated seasonal, broad-range resources through resilient mixed strategies. During this long tradition, millet farming became integral to subsistence, possibly starting as early as the Early Neolithic period by 6400 to 5950 cal BP and becoming firmly established by the Middle Neolithic period. The long-term sustainability of broad-spectrum resource use in Korean Neolithic cases shows a contrast to the vulnerability of the highly specialized subsistence strategy employed in the Early-Middle Jomon culture in northeast Japan (Habu Citation2015).
Our study also reaffirms the importance of multi-line approaches to diet reconstruction and the importance of applying rigorous recovery efforts to samples that are often not easily visible without the use of microscopic techniques. In addition, research on Neolithic subsistence in Korea has been impaired without data on economic wild plant resources. With rigorous soil sampling and flotation using fine mesh, this study was able to provide a glimpse into the utilization of a much greater diversity of grain, fruit, and nut taxa. Prior to our earlier studies on later prehistoric sites (Kwak and Marwick Citation2015; Kwak, Kim, and Lee Citation2017) and two recent studies by Heron et al. (Citation2016) and Shoda et al. (Citation2017), isotope analyses had only been applied only to human remains in Korea. Our integrative approach opens the door to using the most common artifacts of the Korean Neolithic, pottery, to better understand the evolution of human diets and subsistence strategies in this region.
uica_a_1776427_sm6557.zip
Download Zip (1.6 MB)Acknowledgements
We appreciate the field staff members at the Dongkuk University Museum, Gimhae National Museum, and Busan Museum who assisted us while collecting data, including Dr. Insoo Ha, Dr. Jaeho Ahn, Dr. Hakjong Lim, and Dr. Jeongguen Lee. Dr. Sangtaek Lim (Pusan National University) and Dr. Bosik Hong (Bokcheon Municipal Museum) also helped us collect pottery samples and shares their knowledge on sites. A special thanks to Dr. Richard Evershed (University of Bristol), Dr. Julie Dunne (University of Bristol), and Dr. Marisol Correa-Ascencio for sharing their knowledge related to the lipid extraction and isotope analysis. Dr. Rory Walsh (University of Michigan), Dr. Maureece Levin (Stanford University), Dr. Habeom Kim (University of Oregon), and Hyunsoo Lee (University of Oregon) provided comments on an earlier version of this paper. Dr. Kim prepared and . We appreciate the anonymous reviewers and the editor for their comments on the earlier draft.
Additional information
Funding
References
- Ahn, S. M. 1988. The Neolithic age. Journal of Korean Archaeological Society 21:35–62.
- Aikens, C. M., and G.-A. Lee. 2013. Postglacial inception and growth of anthropogenic landscapes in China, Korea, Japan, and the Russian Far East. Anthropocene 4:46–56. doi:https://doi.org/10.1016/j.ancene.2014.02.005
- Ambers, J. C. 1990. Identification of the use of marine plant material as animal fodder by stable isotope ratios. PACT: Journal of the European Study Group on Physical, Chemical and Mathematical Techniques Applied to Archaeology 29:251–8.
- Ambrose, S. H., and M. A. Katzenberg. 2002. Biogeochemical approaches to paleodietary analysis. Advances in Archaeological and Museum Science. New York: Springer.
- An, D. I. 2006a. Dietary reconstruction at the Neolithic Konam-ri and Daejuk-ri sites by trace element analysis. Journal of the Korean Ancient Historical Society 53:23–41.
- An, D. I. 2006b. Dietary reconstruction by stable isotopic analysis: The Konam-ri shell midden in Korea. Journal of the Korean Ancient Historical Society 54:5–20. doi:https://doi.org/10.18040/sgs.2017.95.5
- An, D. I. 2009. Dietary reconstruction through trace element (Ba, Sr, Zn) analysis of human bones from the Neolithic Yondae-do site. Journal of the Korean Ancient Historical Society 66:5–26.
- Bae, K., C. J. Bae, and J. C. Kim. 2013. Reconstructing human subsistence strategies during the Korean Neolithic: contributions from zooarchaeology, geosciences, and radiocarbon dating. Radiocarbon 55 (3):1350–7. doi:https://doi.org/10.1017/S0033822200048268
- Barton, L., S. D. Newsome, F.-H. Chen, H. Wang, T. P. Guilderson, and R. L. Bettinger. 2009. Agricultural origins and the isotopic identity of domestication in northern China. Proceedings of the National Academy of Sciences of the United States of America 106 (14):5523–8. doi:https://doi.org/10.1073/pnas.0809960106
- Bettinger, R. L. 2002. Cultural, human and historical ecology in the Great Basin: Fifty years of ideas abut then thousand years of prehistory. In Advances in historical ecology, ed. W. Balée, 169–89. Columbia: Columbia University Press.
- Bevan, A., E. Crema, and F. Silva. 2018. Rcarbon v1.1.3: Methods for calibrating and analysing radiocarbon dates. https://CRAN.R-project.org/package=rcarbon
- Bentley, R. A., N. Tayles, C. Higham, C. Macpherson, and T. Atkinson. 2007. Shifting Gender Relations at Khok Phanom Di, Thailand: Isotopic evidence from the skeletons. Current Anthropology 48 (2):301–14. doi:https://doi.org/10.1086/512987
- Bishop, R. R., M. J. Church, and P. A. Rowley-Conwy. 2015. Firewood, food and human niche construction: The potential role of Mesolithic hunter–gatherers in actively structuring Scotland’s woodlands. Quaternary Science Reviews 108:51–75. doi:https://doi.org/10.1016/j.quascirev.2014.11.004
- Bleed, P., and A. Matsui. 2010. Why didn’t agriculture develop in Japan? A consideration of Jomon ecological style, niche construction, and the origins of domestication. Journal of Archaeological Method and Theory 17 (4):356–70. doi:https://doi.org/10.1007/s10816-010-9094-8
- Boivin, N. L., M. A. Zeder, D. Q. Fuller, A. Crowther, G. Larson, J. M. Erlandson, T. Denham, and M. D. Petraglia. 2016. Ecological consequences of human niche construction: Examining long-term anthropogenic shaping of global species distributions. Proceedings of the National Academy of Sciences of the United States of America 113 (23):6388–96. doi:https://doi.org/10.1073/pnas.1525200113
- Bokcheon Museum, ed. 2011. Dongsam-Dong shell midden site. Busan: Bokcheon Museum Press.
- Braje, T., T. Leppard, S. Fitzpatrick, and J. Erlandson. 2017. Archaeology, historical ecology and anthropogenic island ecosystems. Environmental Conservation 44 (3):286–97. doi:https://doi.org/10.1017/S0376892917000261
- Broughton, J. M. M. D. Cannon, and E. J. Bartelink. 2010. Evolutionary ecology, resource depression, and Niche construction theory: Applications to central california hunter-gatherers and mimbres-mogollon agriculturalists. Journal of Archaeological Method and Theory 17 (4):371–421. doi:https://doi.org/10.1007/s10816-010-9095-7.
- Busan Museum, ed. 2007. Dongsam-dong Shell Midden Site: Reconstruction Site Report. Busan: Busan Museum Press.
- Busan Museum, ed. 2009. Beombang Site. Busan: Busan Museum Press.
- Choy, K., D. I. An, and M. P. Richards. 2012. Stable isotopic analysis of human and faunal remains from the Incipient Chulmun (Neolithic) shell midden site of Ando Island. Journal of Archaeological Science 39 (7):2091–7. doi:https://doi.org/10.1016/j.jas.2012.03.005
- Choy, K., and M. P. Richards. 2010. Isotopic evidence for diet in the Middle Chulmun period: A case study from the Tongsamdong shell midden. Archaeological and Anthropological Sciences 2 (1):1–10. doi:https://doi.org/10.1016/j.jas.2012.03.005
- Colonese, A. C., T. Farrell, A. Lucquin, D. Firth, S. Charlton, H. K. Robson, M. Alexander, and O. E. Craig. 2015. Archaeological bone lipids as palaeodietary markers. Rapid Communications in Mass Spectrometry: RCM 29 (7):611–8. doi:10/1002/rcm.7144.
- Copley, M. S., Bland, H. A. P. Rose, P. M. Horton, and R. P. Evershed. 2005. Gas chromatographic, mass spectrometric and stable carbon isotopic investigations of organic residues of plant oils and animal fats employed as illuminants in archaeological lamps from Egypt. The Analyst 130 (6):860–71. doi:10/1039/B500403A.
- Correa-Ascencio, M., and R. P. Evershed. 2014. High throughput screening of organic residues in archaeological potsherds using direct acidified methanol extraction. Analytical Methods 6 (5):1330–40. doi:10.1-/1039/C3AY41678J
- Craig, O. E., H. Saul, A. Lucquin, Y. Nishida, K. Taché, L. Clarke, A. Thompson, D. T. Altoft, J. Uchiyama, M. Ajimoto, et al. 2013. Earliest evidence for the use of pottery. Nature 496 (7445):351–4. doi:10/1039/nature12109.
- Craig, O. E., V. J. Steele, A. Fischer, S. Anders, S. H. Hartz, P. Andersen, A. Donohoe, A. Glykou, H. Saul, M. Jones, et al. 2011. Ancient lipids reveal continuity in culinary practices across the transition to agriculture in northern Europe. Proceedings of the National Academy of Sciences of the United States of America 108 (44):17910–5. doi:https://doi.org/10.1073/pnas.1107202108
- Crawford, G. W. 2008. The Jomon in early agriculture discourse: Issues arising from Matsui, Kanehara and Pearson. World Archaeology 40 (4):445–65. doi:https://doi.org/10.1080/00438240802451181
- Crawford, G. W. 2011. Advances in understanding early agriculture in Japan. Current Anthropology 52 (S4):331–45. doi:https://doi.org/10.1086/658369
- Crawford, G. W. 2014. Food production and niche construction in pre-contact southern ontario. Midcontinental Journal of Archaeology. MCJA Occasional Papers No. 1, 135–60. https://www.midwestarchaeology.org/files/MAC-Occasional-Papers-No-1-Revised.pdf.
- Crawford, G. W., and G.-A. Lee. 2003. Agricultural origins in the Korean Peninsula. Antiquity 77 (295):87–95. doi:https://doi.org/10.107/S0003598X00061378
- Davis, N. E., A. Bennett, D. M. Forsyth, D. M. J. Bowman, E. C. Lefroy, S. W. Wood, A. P. Woolnough, P. West, J. O. Hampton, and C. N. Johnson. 2016. A systematic review of the impacts and management of introduced deer (family Cervidae) in Australia. Wildlife Research 43 (6):515–32. doi:https://doi.org/10.1071/WR16148
- Dongguk University Museum. 2008. The Sejukri site report. Gyeongju: Dongguk University Museum Press.
- Dunne, J., R. P. Evershed, M. Salque, L. Cramp, S. Bruni, K. Ryan, S. Biagetti, and S. di Lernia. 2012. First dairying in green Saharan Africa in the fifth millennium BC. Nature 486 (7403):390–4. doi:10/1038/nature/11186.
- Eerkens, J. W. 2005. GC–MS analysis and fatty acid ratios of archaeological potsherds from the western great basin of north America. Archaeometry 47 (1):83–102. doi:https://doi.org/10.1111/j.1475-4754.2995.00189.x
- Enser, M. 1991. Animal carcass fats and fish oils. In Analysis of oilseeds, fats and fatty foods, ed. J. B. Rossel, and J. L. R. Pritchard, 329–94. London: Elsevier.
- Evershed, R. P. 2008a. Organic residue analysis in archaeology: the archaeological biomarker revolution*. Archaeometry 50 (6):895–924. doi:https://doi.org/10.1111/j.1475-4754.2005.00189.x
- Evershed, R. P. 2008b. Experimental approaches to the interpretation of absorbed organic residues in archaeological ceramics. World Archaeology 40:26–47.
- Evershed, R. P., Dudd, S. N. M. S. Copley, R. Berstan, Robert, A. W. Stott, H. Mottram, S. A. Buckley, and Z. Crossman. 2002. Chemistry of archaeological animal fats. Accounts of Chemical Research 35 (8):660–8. doi:https://doi.org/10.1021/ar000200f
- Fitzpatrick, S. M., and J. M. Erlandson. 2009. Islands, zooarchaeology, and historical ecology. The Journal of Island and Coastal Archaeology 4 (2):139–40. Routledge: doi:https://doi.org/10.1080/15564890903203719
- Fitzpatrick, S. M., T. C. Rick, and J. M. Erlandson. 2015. Recent progress, trends, and developments in island and coastal archaeology. The Journal of Island and Coastal Archaeology 10 (1):3–27. doi:https://doi.org/10.1080/15564894.2015.1013647
- Flannery, K. V. 1969. Origins and ecological effects of early domestication in Iran and the Near East. In The domestication and exploitation of plants and animals, ed. P. J. Ucko, and G. W. Dimbleby, 73–100. Chicago: Aldine.
- Fuentes, A. 2015. Integrative anthropology and the human niche: Toward a contemporary approach to human evolution. American Anthropologist 117 (2):302–15. doi:https://doi.org/10.1111/aman.12248
- Gimhae National Museum. 2020. Bibongri Site AMS. Gimhae: Gimhae National Museum. https://gimhae.museum.go.kr/kr/html/sub05/0504.html?mode=V&mng_no=54
- Gimhae National Museum. 2008. The Bibong-ri site. Report on the research of antiquities of the Gimhae National Museum. Changnyeong: Gimhae National Museum Press.
- Ha, I. S. 1997. Bumbang Shell Midden II. Busan: Busan Museum Press.
- Ha, I. S., H. Y. Lee, and H. S. Lee. 1993. Bumbang Shell Midden I. Busan: Busan Museum Press.
- Habu, J. 2015. Mechanisms of long-term culture change and human impacts on the environment: A perspective from historical ecology, with special reference to the Early and Middle Jomon periods of prehistoric Japan. The Quaternary Research (Daiyonki-Kenkyu) 54 (5):299–310. doi:https://doi.org/10.4116/jaqua.54.299
- Heron, C., and O. E. Craig. 2015. Aquatic resources in food crusts: Identification and implication. Radiocarbon 57 (4):707–19. doi:https://doi.org/10.2458/azu_rc.57.18454
- Heron, C., and R. P. Evershed. 1993. The analysis of organic residues and the study of pottery use. Archaeological Method and Theory 5:247–84.
- Heron, C., S. Shoda, A. Breu Barcons, J. Czebreszuk, Y. Eley, M. Gorton, W. Kirleis, J. Kneisel, A. Lucquin, J. Müller, et al. 2016. First molecular and isotopic evidence of millet processing in prehistoric pottery vessels. Scientific Reports 6:38767. doi:https://doi.org/10.1038/srep38767
- Hwang, S., and S-O Yoon. 2002. The environmental change and human activities at Sejuk, Hwangseong-Dong, Ulsan City, southeastern coast of Korea during the middle Holocene. Journal of Korean Archaeological Society 48:35–57.
- Hwang, S., and S-O Yoon. 2005. The change of vegetation and climate during Late Quaternary in Korea. Research Review of Geography 24:154–80.
- Kim, E. Y. 2016. The clay band pottery of the Sugari culture. Seeds Contact 3:18–9.
- Kim, H. S. 2010. The age and dietary concerned about human remain of Daepo shell midden. Journal of Korean Neolithic Society 20:89–111.
- Kim, H. S. 2012. Isotope analysis on the do-g bone from Bibong-ri site. In Bibong-ri Site II, ed. Gimhae Nation Museum, 430–7. Changyeong: Gimhae National Museum Press.
- Kim, J. 2003. Land-use conflict and the rate of the transition to agricultural economy: A comparative study of southern Scandinavia and central-western. Journal of Archaeological Method and Theory 10 (3):277–321. doi:https://doi.org/10.1023/A:1026087723164
- Kim, J. 2006. Resource patch sharing among foragers. In Beyond affluent-foragers rethinking hunter-gatherer complexity, ed. C. Grier and J. Uchiyamam, 168–91. Oxford: Oxbow Books.
- Kim, K. S. 2011. Food culture. In Overview of the Korean Neolithic culture, ed. Jungang Cultural Heritage Institute, 369–86. Daejeon: Junang Cultural Heritage Institute Press.
- Kim, M. 2014. Wild edible plants. In Bronze age archaeology 1: Humans and environment, ed. H. Lee and H. Lee, 103–32. Seoul: Seokyung Press.
- Kwak, S., G. Kim, and G.-A. Lee. 2017. Beyond rice farming: Evidence from central Korea reveals wide resource utilization in the Songgukri culture during the late-Holocene. The Holocene 27 (8):1092–102. doi:https://doi.org/10.1177/0959683616683259
- Kwak, S., and B. Marwick. 2015. What did they cook? A preliminary investigation into culinary practices and pottery use in the central part of the Korean Peninsuladuring the mid to late Holocene. Journal of Indo-Pacific Archaeology 37:25–43. doi:https://doi.org/10.7152/jipa.v37i0.14708
- Laland, K., B. Matthews, and M. W. Feldman. 2016. An introduction to niche construction theory. Evolutionary Ecology 30:191–202. doi:https://doi.org/10.1007/s10682-016-9821-z
- Laland, K. N., and M. J. O’Brien. 2010. Niche construction theory and archaeology. Journal of Archaeological Method and Theory 17 (4):303–25. doi:https://doi.org/10.1177/0959683616689
- Lansing, J. S., and K. N. Fox. 2011. Niche construction on Bali: The Gods of the countryside. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 366 (1566):927–34. doi:https://doi.org/10.1098/rstb.2010.0308
- Lee, D. J. 2006. Review of the environmental change vestiges newly discovered in Dongsam-dong relic. Journal of Honam Archaeological Society 24:5–31.
- Lee, G.-A. 2003. Changes in subsistence patterns from the Chulmun to Mumun periods: Archaeobotanical investigation. Toronto: University of Toronto.
- Lee, G.-A. 2008a. Plant remains recovered from the Sejukri Early Neolithic shell midden. In The Bibong-ri Site, ed. Dongguk University Museum. Gyeongju: Dongguk University Museum Press.
- Lee, G.-A. 2008b. Plant remains recovered from the Bibong-ri shell midden Neolithic site. In The Bibong-ri Site, ed. Gimhae National Museum, 391–8. Gimhae: Gimhae National Museum Press.
- Lee, G.-A. 2011. The transition from foraging to farming in prehistoric Korea. Current Anthropology 52 (S4):307–29. doi:https://doi.org/10.1086/658488
- Lee, G.-A. 2013. Archaeological perspectives on origins of Azuki (Vigna angularis). The Holocene 23 (3):453–9. doi:https://doi.org/10.1177/0959683612460788
- Lee, G.-A. 2016. The spread of domesticated plant resources in prehistoric northeast Asia. In Routledge handbook of archaeology and globalization, ed. T. Hodos, 394–4. London: Routledge Taylor & Francis Group.
- Lee, G.-A. 2017. The Chulmun period of Korea: Current findings and discourse on the Korean Neolithic culture. In Handbook of East and Southeast Asian archaeology, ed. J. Habu, P. V. Lape, and J. W. Olsen, 451–82. New York: Springer-Verlag.
- Lee, G.-A., M.-S. Cho, and H. Obata. 2019. Coastal farmers during the Early and Middle Holocene: Archaeobotanical and grain impression studies on the east coast Korea. Quaternary International 529:57–74. doi:https://doi.org/10.1016/j.quaint.2019.10.013
- Lee, G.-A., G. W. Crawford, L. Liu, Y. Sasaki, and C. Xuexiang. 2011. Archaeological soybean (Glycine max) in East Asia: Does size matter? PLoS One 6 (11):e26720. doi:https://doi.org/10.1371/journal.pone.0026720
- Lee, J.-J. 2001. From shellfish gathering to agriculture in prehistoric Korea: The Chulmun to Mumun transition. Madison: University of Wisconsin-Madison.
- Lee, J.-J. 2002. Changing subsistence strategies during the Chulmun Period at Southeastern Korea. Journal of Korean Archaeological Society 79:59–92.
- Lee, J.-J. 2011a. Intensification of millet and rice agriculture in Korea: Evidence from stable isotopes. Journal of the Korean Ancient Historical Society 73:31–66.
- Lee, J.-J. 2011b. Domesticated pig in Korea: Its socioeconomic and symbolic context. Journal of Korean Archaeological Society 79:131–74.
- Lee, S.-M., and E.-J. Lee. 2019. Diet of the wild boar (Sus scrofa): Implications for management in forest-agricultural and urban environments in South Korea. PEER J. 7:e7835. doi:https://doi.org/10.7717/peerj.7835
- Lee, T. 2014. Korean flora. 5th ed. Seoul: Hyangmunsa.
- Lee, Y. D. 2011. Subsistence patterns. In Introduction to Korean Neolithic period, ed. Central Institute of Cultural Heritage, 335–68. Seoul: Seokyung Press.
- Levin, M. J., M. Shelton, and W. S. Ayres. 2017. Swiddening practices in the Micronesian past: Macrocharcoal quantification from a gardening site in Phonpei. The Journal of Island and Coastal Archaeology 12 (3):451–7. doi:https://doi.org/10.1080/15564894.2016.1271066
- Linares, O. F. 1976. “Garden hunting” in the American tropics. Human Ecology 4 (4):331–49. doi:https://doi.org/10.1007/BF01557917
- Lucquin, A., A. C. Colonese, T. F. G. Farrell, and O. E. Craig. 2016. Utilizing phytanic acid diastereomers for the characterization of archaeological lipid residues in pottery samples. Tetrahedron Letters 57 (6):703–7. doi:https://doi.org/10.1016/j.tetlet.2016.01.011
- Lucquin, A., K. Gibbs, J. Uchiyama, H. Saul, M. Mayumi, Y. Eley, A. Radini, C. P. Heron, S. Shoda, N. Yastami, et al. 2016. Ancient lipids document continuity in the use of early hunter-gatherer pottery through 9,000 years of Japanese prehistory. Proceedings of the National Academy of Sciences of the United States of America 113 (15):3991–6. doi:https://doi.org/10.1073/pnas.1522908113
- Moss, M. L., A. T. Rodrigues, C. F. Speller, and D. Y. Yang. 2016. The historical ecology of Pacific herring: Tracing Alaska Native use of a forage fish. Journal of Archaeological Science: Reports 8:504–12. doi:https://doi.org/10.1016/j.jasrep.2015.10.005
- Mottram, H. R., S. N. Dudd, G. J. Lawrence, A. W. Stott, and R. P. Evershed. 1999. New chromatographic, mass spectrometric and stable isotope approaches to the classification of degraded animal fats preserved in archaeological pottery. Journal of Chromatography A 833 (2):209–21. doi:https://doi.org/10.1016/S0021-9673(98)01041-3
- Mueller, N. G. 2017. Documenting domestication in a lost crop (Polygonum erectum L.): Evolutionary bet-hedgers under cultivation. Vegetation History and Archaeobotany 26 (3):313–27. doi:https://doi.org/10.1007/s00334-016-0592-9
- National Research Institute of Cultural Heritage. 2013. Munam-ri at Goseong Vol.1. Daejeon: National Research Institute of Cultural Heritage Press.
- Nishimoto, T., H. S. Kim, and M. Niimi. 2011. Analysis on the faunal remains discovered from Dongsam-dong shellmidden Neolithic site. In Dongsam-dong Shell Midden Site: Faunal remains, ed. Bokcheon Museum, 35–87. Busan: Bokcheon Museum Press.
- Obata, H. 2013. Descriptions on the diagnostic characteristics for identification of millet impressions from Dongsam-dong shellmidden and Pibongni site. Journal of Korean Neolithic Society 7:105–55.
- Obata, H., and A. Manabe. 2012. Changnyeong Bibongri pottery impression study. In Bibongri II, ed. Gimhae National Museum, 267–91. Gimhae: Gimhae National Museum Press.
- Obata, H., A. Manabe, N. Nakamura, T. Onishi, and Y. Senba. 2011. A new light on the evolution and propagation of prehistoric grain pests: the world's oldest maize weevils found in Jomon Potteries, Japan. Plos One 6 (3):e14785. doi:https://doi.org/10.1371/journal.pone.0014785
- Odling-Smee, J., D. H. Erwin, E. P. Palkovacs, M. W. Feldman, and K. N. Laland. 2013. Niche construction theory: A practical guide for ecologists. The Quarterly Review of Biology 88 (1):3–28. doi:https://doi.org/10.1086/669266
- Pai, H. I. 2016. Archaeologizing the Korean heritage cultural properties management and state tourist development. In Handbook of East and Southeast Asian archaeology, J. Habu, Junko, J. W. Olsen, and P. V. Lape, 27–37. New York: Springer Press.
- Pearsall, D. M. 2015. Paleoethnobotany: A handbook of procedures. 3rd ed. London: Routledge.
- Pechenkina, E. A., S. H. Ambrose, X. Ma, and R. A. Benfer. 2005. Reconstructing northern Chinese Neolithic subsistence practices by isotopic analysis. Journal of Archaeological Science 332:1176–89. doi:https://doi.org/10.1016/j.jas.2005.02.015
- Price, T. D., and O. Bar-Yosef. 2011. The origins of agriculture: New data, new ideas: An introduction to Supplement 4. Current Anthropology 52 (S4):S163–S74. doi:https://doi.org/10.1086/659964
- Ramsey, C. B. 2001. Development of the radiocarbon calibration program. Radiocarbon 43:355–63. doi:https://doi.org/10.1017/S0033822200038212
- Reber, E. A., and R. P. Evershed. 2004. How did Mississippians prepare maize? The application of compound-specific carbon isotope analysis to absorbed pottery residues from several Mississippi valley sites. Archaeometry 46 (1):19–33. doi:https://doi.org/10.1111/j.1475-4754.2004.00141.x
- Reimer, P. J., E. Bard, A. Bayliss, J. W. Beck, P. G. Blackwell, C. B. Ramsey, C. E. Buck, H. Cheng, R. L. Edwards, M. Friedrich, et al. 2013. IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55 (4):1869–87. doi:https://doi.org/10.2458/azu_js_rc.55.16947
- Riel-Salvatore, J. 2010. A niche construction perspective on the Middle–Upper Paleolithic transition in Italy. Journal of Archaeological Method and Theory 17 (4):323–55. doi:https://doi.org/10.1007/s10816-010-9093-9
- Rowley-Conwy, P., and R. Layton. 2011. Foraging and farming as niche construction: Stable and unstable adaptations. Philosophical Transactions of the Royal Society B: Biological Science 366 (1566):849–62. doi:https://doi.org/10.1098/rstb.2010.0307
- Rural Development Administration. 1988. Technical report on Korea Soil. Seoul: Rural Development Administration (RDA) of Republic of Korea.
- Salque, M., P. I. Bogucki, J. Pyzel, I. Sobkowiak-Tabaka, R. Grygiel, M. Szmyt, and R. P. Evershed. 2013. Earliest evidence for cheese making in the sixth millennium BC in northern Europe. Nature 493 (7433):522–5. doi:https://doi.org/10.1038/nature11698
- Sample, L. L. 1974. Tongsamdong: A contribution to Korean Neolithic culture History. Arctic Anthropology 11:1–125.
- Schulting, R. J., P. Vaiglova, R. Crozier, P. J. Reimer, and J. Paula. 2017. Further isotopic evidence for seaweed-eating sheep from Neolithic Orkney. Journal of Archaeological Science: Reports 11:463–70. doi:https://doi.org/10.1016/j.jasrep.2016.12.017
- Shin, J. Y., D. Y. Kang, S. H. Kim, and E. D. Jung. 2013. Isotopic dietary history of Neolithic people from Janghang site at Gadeok Island. Busan. Analytical Science and Technology 26 (6):387–94. doi:https://doi.org/10.5806/AST.2013.26.6.387
- Shoda, S., A. Lucquin, J.-H. Ahn, C.-J. Hwang, and O. E. Craig. 2017. Pottery use by early Holocene hunter-gatherers of the Korean peninsula closely linked with the exploitation of marine resources. Quaternary Science Reviews 170:164–73. doi:https://doi.org/10.1016/j.quascirev.2017.06.032
- Smith, A., and N. F. Miller. 2009. Integrating plant and animal data: Delving deeper into subsistence (Introduction to the Special Section). Current Anthropology 50 (6):883–4. doi:https://doi.org/10.1086/605867
- Smith, B. D. 2011. The cultural context of plant domestication in eastern North America. Current Anthropology 52 (S4):S471–S84. doi:https://doi.org/10.1086/659645
- Smith, B. D. 2015. A comparison of niche construction theory and diet breadth models as explanatory frameworks for the initial domestication of plants and animals. Journal of Archaeological Research 23:215–62. doi:https://doi.org/10.1007/s10814-015-9081-4
- Smith, B. D. 2016. Neo-Darwinism. Niche construction theory, and the initial domestication of plants and animals. Evolutionary Ecology 30:307–24. doi:https://doi.org/10.1007/s10682-015-9797-0
- Smith, D. A. 2005. Garden game: Shifting cultivation, indigenous hunting and wildlife ecology in Western Panama. Human Ecology 33 (4):505–37. doi:https://doi.org/10.1007/s10745-005-5157-Y
- Still, C. J., J. Berry, G. J. Collartz, and R. S. Defries. 2003. Global distribution of C3 and C4 vegetation: Carbon cycle implications. Global Biochemical Cycles 17:1006. doi:https://doi.org/10.1029/2001GB001807
- Takeda, T., and S. Hakoyama. 1985. Studies on the ecology and geographical distribution of C3 and C4 grasses: II. Geographical distribution of C3 and C4 grasses in far East and South East Asia. Japanese Journal of Crop Science 54 (1):65–71. doi:https://doi.org/10.1626/jcs.54.65
- Takeda, T., T. Tanlkawa, W. Agata, and S. Hakoyama. 1985. Studies on the ecology and geographical distribution of C3 and C4 grasses : I. Taxonomic and geographical distribution of C3 and C4 grasses in Japan with special reference to climatic conditions. Japanese Journal of Crop Science 54 (1):54–64. doi:https://doi.org/10.1626/jcs.54.54
- Winterhalder, B., and C. Goland. 1997. An evolutionary ecology perspective on diet choice, risk, and plant domestication. In Plants, people, and landscapes: Studies in paleoethnobotany, ed. K. Gremillion, 123–60. Tuscaloosa: University of Alabama Press.
- Yi, T.-B. 2014. The flora of Korea. Seoul: Hyangmun-sa.
- Zeder, M. A. 2012. The broad spectrum revolution at 40: Resource diversity, intensification, and an alternative to optimal foraging explanations. Journal of Anthropological Archaeology 31 (3):241–64. doi:https://doi.org/10.1016/j.jaa.2012.03.003
- Zeder, M. A. 2016. Domestication as a model system for niche construction theory. Evolutionary Ecology 30:325–48. doi:https://doi.org/10.1007/s10682-015-9801-8