
Abstract
Despite decades of archaeological research on Jamaica, little is known about how settlers influenced landscape change on the island over time. Here, we examine the impact of human occupation through a multi-proxy approach using phytolith, charcoal, and stratigraphic analyses. White Marl was a continuously inhabited village settlement (ca. 1050–450 cal yrs BP) with large mounded midden areas, precolonial house structures, and human landscape management practices. We have shown that the local vegetation at White Marl was directly affected by human settlement through the use of agroforestry and burning, and suggest that fire was used to modify vegetation. Manioc phytoliths were found throughout human occupation and are broadly associated with increases in evidence for burning, suggesting fire was used to modify the landscape and clear vegetation for crop cultivation. The phytolith assemblages relate to three distinct temporal vegetation phases: (1) the earliest occupation dominated by arboreal vegetation (pre-ca. 870 cal yrs BP); (2) a transition to palm-dominated vegetation (ca. 870–670 cal yrs BP); and (3) the latest occupation representing European colonization associated with a more open, grass-dominated landscape (after ca. 670 cal yrs BP). These transitions occur independent of changes in paleoclimate records, suggesting humans were the dominant driver of vegetation change.
Introduction
Environmental archaeology in the Caribbean has a long history of multidisciplinary research, combining anthropological, biological, and environmental datasets (LeFebvre, Giovas, and Laffoon Citation2019). Faunal analysis has received particular attention since the 1940s (e.g., Chabanaud Citation1946) and increasingly since the late 1960s (e.g., Bochaton et al. Citation2015; Boudadi-Maligne et al. Citation2016; Giovas Citation2018; LeFebvre and DeFrance Citation2014; Pestle Citation2013; Steadman et al. Citation2017; Wing Citation1961, Citation1972). Comparatively, the recovery of plant remains by flotation and dry sieving in the Caribbean has been less common (Newsom and Wing Citation2004), but has steadily increased since the late 1980s, though it is geographically patchy (e.g., Bain et al. Citation2018; Hooghiemstra et al. Citation2018; Kennedy, Horn, and Orvis Citation2006; Newsom Citation1988; Pearsall Citation1985).
Botanical remains in archaeological sediments can provide important evidence for the exploitation of economically useful plant taxa and subsistence patterns and help to identify human influence on local environments. Based on existing studies throughout the Caribbean, human-induced environmental changes have varied (e.g., Berman and Pearsall Citation2020; Ciofalo, Sinelli, and Hofman Citation2019; Pagán-Jiménez et al. Citation2020; Siegel et al. Citation2018). It is widely known, however, that early inhabitants cleared and impacted landscapes using fire (Cochrane Citation2009; Fitzpatrick and Keegan Citation2007; Sponsel Citation1998), a strategy that seems to have been used for millennia in the Caribbean (Bain et al. Citation2018; Burney, Burney, and MacPhee Citation1994; Caffrey and Horn Citation2015; Richardson Citation2004; Siegel et al. Citation2015).
Both macro- and micro-botanical proxies are increasingly used to understand subsistence patterns, plant exploitation, and plant domestication (Castilla-Beltrán et al. Citation2018, Citation2020; Ciofalo, Sinelli, and Hofman Citation2020; Fitzpatrick Citation2015; Lane et al. Citation2009; Newsom and Wing Citation2004; Pagán-Jiménez Citation2007, Citation2013; Pagán-Jiménez, Rodríguez-Ramos, and Hofman Citation2019; Pagán-Jiménez et al. Citation2020; Siegel et al. Citation2015). Human–environment interactions are most effectively examined by analyzing phytoliths and a combination of other micro-proxies such as starch and pollen. Phytolith and pollen research conducted within the region have shown that anthropogenic modifications of the landscape were gradual and locally variable (e.g., Siegel et al. Citation2018).
For example, studies of plant remains in the Dominican Republic from multi-layered household middens, which are formed from a combination of natural and cultural processes, have targeted site-wide human–plant relationships and show the consolidation and configuration of local foodway systems ahead of European colonialism (Pagán-Jiménez et al. Citation2020). Furthermore, this analysis at two precolonial household mounds bearing Ostionoid/Meillacoid and Meillacoid/Chicoid ceramics (ca. 870–500 cal yrs BP) indicates that the inhabitants adopted a polyculture agricultural strategy (Pagán-Jiménez Citation2007; Pagán-Jiménez et al. Citation2020). Ciofalo, Sinelli, and Hofman (Citation2019) also compared sites in the Dominican Republic along with the Turks and Caicos Islands where contemporaneous sites revealed the remains of a range of food-based plants, including maize, manioc, and guáyiga/coontie/zamia. Micro-botanical analyses in The Bahamas have identified a package of domesticated plants, including Cucurbita (squash), Manihot esculenta (manioc), Calathea (leren or arrowroot), Zea mays (maize), and Capsicum sp. (chili) (Berman and Pearsall Citation2020). Other studies in the region include the identification of anthropogenic burning in the late Archaic Period (ca. 2700 cal yrs BP) (Sara and Aguilú Citation2003); investigations into the timing of human arrival (Burney, Burney, and MacPhee Citation1994) in Puerto Rico; the dispersal of (domesticated food) plants into and within the region (Berman and Pearsall Citation2020; Pagán-Jiménez et al. Citation2015), as well as local and regional variations in diet, feasting, and ceremonial activities (e.g., Mickleburgh and Pagán-Jiménez Citation2012).
Until recently, little was known about how different groups influenced or impacted landscape change in Jamaica due to a dearth of botanical studies. As such, the primary goal of the micro-botanical approach adopted here is to disentangle the long-term influence of anthropogenic disturbance regimes on Jamaica’s landscape by identifying changes in the vegetation composition utilizing botanical studies. We aim to investigate the legacy of land use around White Marl (ca. 1050–450 cal yrs BP) by examining how the initial and subsequent occupation impacted local vegetation at the site (locally to the sampled trench, ca. 5 m2 of the sampling location; see SI for more details). Second, we aim to assess new phytolith evidence from White Marl and place it within the context of local and regional paleoenvironmental studies to establish whether any landscape modifications resulted from human-induced disturbances or natural drivers of environmental change such as climate. A further aim is to identify evidence for dietary indicators and alterations in subsistence strategy.
In this paper, we combine independent lines of evidence to document human occupation, including phytoliths, macro-charcoal, and stratigraphic data from one of the excavation units at White Marl. Previous work in Puerto Rico documented human occupation and land management from archaeological deposits based exclusively on increases in charcoal frequencies (Burney Citation1997; Burney, Burney, and MacPhee Citation1994). However, utilizing independent lines of evidence which may relate to human-induced changes, and that combine charcoal analyses with phytoliths and stratigraphy, strengthens this type of investigation (Siegel et al. Citation2015). Phytoliths were used instead of pollen due to the dry nature of the deposits where pollen is unlikely to be well preserved. In addition, phytoliths represent an in situ vegetation signal, localized to the sampling area (Piperno Citation2006). Comparatively, pollen represents a much wider vegetation signal as many pollen morphotypes can travel hundreds of kilometers from the source (Faegri and Iversen Citation1989). Here we present the first extensive phytolith study in Jamaica based on sediments from one of the largest and most well-studied archaeological sites in Jamaica, the Taíno site of White Marl.
Archaeological context
The Caribbean islands were colonized in two major population dispersals possibly from both Central and South America beginning around 7,000 years ago. The first major migration was in the Archaic Age (ca. 7000–2500 BP) by semi-mobile groups with lithic and shell tools and some simple pottery. The early Archaic Age is thought to have been characterized by a hunter-gatherer-forager food economy and social organization without formalized social or political roles. In contrast to what was previously believed, research over the past decade has shown that the end of the Archaic Age was not a rapid technological, political, and societal shift as the result of a migration, but rather a gradual development of subsistence, material culture, and settlement patterns (Chanlatte Baik Citation1995; Keegan Citation1994, Citation2006; Newsom and Wing Citation2004; Pagán-Jiménez Citation2005; Rodríguez Suárez and Pagán-Jiménez Citation2008; Wilson Citation2007).
The second major migration originated in the Lower Orinoco of South America and brought along characteristic material culture, notably including a distinct and refined type of ceramics referred to as the Saladoid series (Petersen, Hofman, and Curet Citation2004; Rouse Citation1986, Citation1992). This marked the archaeological period known as the Early Ceramic Age in the region (ca. 2500–1200/1400 BP). The Early Ceramic Age is believed to have consolidated developments that started in the Archaic Age, including permanent settlements and a food economy based on horticulture and hunting-fishing (Boomert Citation1999; Siegel Citation1992). The precise manner and route of the dispersion of Saladoid material culture and whether this marks the spread of a distinct group of people (and if so, which cultural groups are associated with this), is still a matter of debate. Linguistically, evidence strongly suggests that these groups spoke variants of the Arawakan language family (Granberry and Vescelius Citation2004).
Some scholars have argued that Archaic Age groups in the Greater Antilles persisted for hundreds of years (particularly in Hispaniola and Cuba) after the arrival of Saladoid groups in Puerto Rico. Archaic Age groups are argued to have played a significant role through multifocal development of Archaic peoples and interaction with Saladoid peoples (Rodríguez Suárez and Pagán-Jiménez Citation2008). Others have argued that Archaic peoples were responsible for the later development of ceramic styles and technologies of the Ostionoid and Meillacoid (and eventually “Taíno” societies) (Keegan Citation2006). The Ostionoid and Meillacoid ceramic styles mark the beginning of the Late Ceramic Age in this region (1200/1400–500 BP). In the Lesser Antilles, the widespread Saladoid ceramics and other material culture of the Early Ceramic Age was similarly replaced by significant material culture diversification during the Late Ceramic Age, producing a variety of pottery styles including the Suazoid, Troumassoid, or Cayoid series (Bright Citation2011; Hofman and Hoogland Citation2004; Petersen, Hofman, and Curet Citation2004). In the Late Ceramic Age, social organization in the Greater Antilles and parts of the northern Lesser Antilles included ingrained hierarchical status differentiation and regional-scale political organization, thought to be characterized by “chiefdoms” (Curet Citation1992; Hoogland and Hofman Citation1999; Keegan Citation2000; Wilson Citation2007). However, the archaeological evidence of socio-political organization suggests that it was diverse across the islands. The assumed existence of “chiefdoms” rests largely on interpretations of early historical documents describing Spanish contacts with and ideas about the Indigenous peoples of these islands. It is clear from these records, and studies on material culture style and human and animal mobility, that the region consisted of interlocking regional networks (Mol Citation2014).
The island of Jamaica certainly was a part of this Pan-Caribbean system, but it appears to have been settled relatively late (ca. 1350–1300 BP) compared to the other islands. To date, no securely dated Archaic or Early Ceramic Age settlements have been found, although multiple large Late Ceramic Age settlements are known. These sites are associated with local varieties of Meillacoid ceramics, which are believed to represent two distinct migrations originating in the area of western Puerto Rico and eastern Dominican Republic around 1350 BP (Allsworth-Jones Citation2008; Atkinson Citation2006; Callaghan Citation2008; Keegan and Hofman Citation2017; Napolitano et al. Citation2019). Significantly, no Ostionan ceramics (redware) have been identified in the lower stratigraphic levels of Meillacoid sites like White Marl (Wesler Citation2013). While the apparent lack of earlier habitation of Jamaica is not well understood, it has been suggested that dangerous currents and prevailing winds discouraged traversing the seas between Jamaica and its nearest neighboring islands (Allsworth-Jones Citation2008; Atkinson Citation2006).
Archaeobotany in Jamaica
While environmental archaeological techniques are commonly used in other parts of the Caribbean to assess human–plant relationships, paleobotanical approaches are very limited in Jamaica. Jamaica remains one of the least studied islands in the Greater Antilles (Rampersad Citation2009). There is a relatively short human history in Jamaica that was punctuated by distinct vegetation shifts associated with fire, activity resulting from the earliest colonization of the island (Williams Citation2019). The human–environment disturbance regimes are associated with the initial arrival of early Meillacoid ceramic bearing populations beginning around 1050 cal yrs BP (Napolitano et al. Citation2019) and the European development of industrial sugar cultivation after 440 BP (Horst and Garner Citation2007). Any shifts in human occupation could potentially be associated with diminished plant taxa, eradicated or selectively nurtured as a result of active human land management (Siegel et al. Citation2015).
From an archaeological perspective, limited work has been done with regard to human–environment interactions and the reconstruction of the vegetation prior to European colonization in Jamaica in the late fifteenth century (Atkinson Citation2006, 97). Existing paleobotanical studies for Jamaica have predominantly been utilized at archaeological sites to identify plants of economic importance in targeted archaeological contexts only (Newsom and Wing Citation2004; Siegel et al. Citation2015, Citation2018), and extraction of micro-fossils from archaeological sediments is absent in existing studies. Micro-botanical evidence such as silica phytoliths, pollen, and starch, is currently limited to a study of the human diet using three dental calculus samples from White Marl (Mickleburgh et al. Citation2019). In Jamaica, large multi-proxy paleoenvironmental studies have been conducted on lake cores that incorporate pollen, charcoal, ostracod, and dung fungal spore data (Williams Citation2019); however, this present study is the first from Jamaica to analyze sediments for phytoliths and charcoal directly associated with human occupation. In this study, we investigate the potential influence of different land use systems on the local environment and human–plant interrelationships at the site of White Marl using a multi-proxy micro-botanical approach, following recommendations from recent micro-botanical research in the region (Pagán-Jiménez et al. Citation2020).
The White Marl site
White Marl is one of the largest and most archaeologically studied precolonial Indigenous settlements in Jamaica (Allsworth-Jones Citation2008; Clair Citation1970; Howard Citation1956, Citation1965; Silverberg, Vanderwal, and Wing Citation1972; Vanderwal Citation1968). The site, which has been dated to ca. 1050–450 cal yrs BP (Silverberg, Vanderwal, and Wing Citation1972) is located on the south coast of Jamaica around 2.8 miles east of the Spanish colonial capital of Villa de la Vega (now Spanish Town), and about 8 miles west of Kingston. Its location on a hilltop close to the Rio Cobre and Fresh River, as well its proximity to Kingston Harbor, provided access to both terrestrial and marine resources (Aarons Citation1983; Allsworth-Jones Citation2008, Howard Citation1956) (). Like contemporaneous communities across the Greater Antilles, White Marl is believed to have developed a Taíno chiefdom socio-political organization structure by ca. 750 cal yrs BP. The area was subsequently occupied by a series of British colonial sugar plantations and livestock pens, including the Caymanas Estate and Windsor Pen. The earliest records for the Caymanas Estate are from 210 to 200 BP (https://www.ucl.ac.uk/lbs/).
Figure 1. (A) Inset: Location of Jamaica. Main: Location of White Marl site; (B) White Marl recent excavation areas (Zone A and Zone B) separated by the Nelson Mandela Highway, location of Unit T2-11, control test pit and museum circled in red.
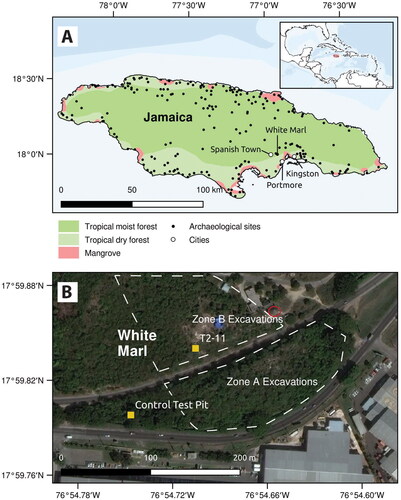
Excavations in the late 1950s and 1960s focused primarily on midden areas that were likely located in excavation area Zone B () based on comparisons with topography and early maps of the area. Significant finds included human burials inside large mounded middens, faunal assemblages, the first precolonial house structure identified on the island, green stone axes, decorated shell adornments, and a relatively homogenous and regionally significant ceramic tradition known as the White Marl ceramic style, which is one of three local variants of the Meillacoid series (see of panel) (Allsworth-Jones Citation2008; Atkinson Citation2006; Clair Citation1970; Keegan and Hofman Citation2017; Silverberg, Vanderwal, and Wing Citation1972; Wesler Citation2013; Wilson Citation2007). At contemporaneous sites with Meillacoid ceramics in the northern Dominican Republic and on other islands in the Greater Antilles, house and burial areas are not separated; middens form within the settlement area around the houses, and were used as burial areas and agricultural gardens/plots. The characteristic stratigraphy observed at these sites probably represents evidence of a “complex, engineered landscape” (Pagán-Jiménez et al. Citation2020, 3) with different episodes of surface leveling for household floors or other activities, deposition of alternating layers of food refuse with artifacts, and intentionally deposited ash and soil layers (Hofman and Hoogland Citation2015; Hofman et al. Citation2018; Pagán-Jiménez et al. Citation2020). Parallels to this can be observed at White Marl, including in the scale and extent of the intentional deposition of marl layers alternating with cultural strata (Allsworth-Jones Citation2008; Howard Citation1965), although the spatial organization of the site is less well understood.
Figure 2. (A) aerial view of White Marl 2019, showing locations of excavation trench including Unit T2-11 and control test pit (orthomosaic composite image made in DroneDeploy); (B) aerial view showing excavation of trench including Unit T2-11 in 2018; (C) aerial view showing extended excavation trench and Unit T2-11 in 2019, dashed line indicates the sampled profile; (D–E) ceramics from Unit T2-11 recovered during excavations and sampling, showing typical decorations including incisions, punctuations and molded clay protrusions; (F) bowl fragment from excavations at White Marl. All aerial photographs taken by drone (DJIMavic Pro) by Zachary J. M. Beier 2018–19.

Archaeobotanical studies of dental calculus from three of the White Marl burials from excavation Zone A (see Figure 1, ca. 360–680 cal yrs BP) identified Marentaceae (arrowroot), Arecaceae (palms), Cannaceae (achira), maize Z. mays, Fabaceae (wild beans), and possible identification of Theobroma cacao (chocolate) (Mickleburgh et al. Citation2019). Our investigations focused on one of the more extensive profiles in Zone B (western profile of T2-11), which commences at the bedrock and extends upwards ca. 2 m. Unit T2-11 is located within a large mounded midden/refuse area in the southern portion of the White Marl development corridor (). Excavations led by archaeologists from the Jamaica National Heritage Trust (JNHT) and the University of West Indies (UWI) Mona between 2018 and 2019 in this area recovered a large assemblage of artifacts within well preserved stratified deposits along with three human burials.
Previous radiocarbon dates on charcoal range from ca. 1400 to 670 cal yrs BP (Allsworth-Jones Citation2008; Silverberg, Vanderwal, and Wing Citation1972; recalibrated using IntCal20, Reimer et al. Citation2020, see SI), and ca. 610–390 cal yrs BP (Mickleburgh et al. Citation2019, recalibrated using IntCal20, Reimer et al. Citation2020, see SI). Two new radiocarbon dates from charcoal were obtained in 2019 from the section sampled for phytolith analysis used in this study from Unit T2-11 in Zone B. These dates range from ca. 870 to 670 cal yrs BP (, see SI for recalibration with IntCal20).
Table 1. Radiocarbon dates from charcoal collected from the western profile of Unit T2-11 in Zone B.
Materials and methods
Sampling, stratigraphic analysis, and recording
In July 2019 a total of 21 sediment samples and two charcoal samples for radiocarbon dating were taken from the western profile of the open trench, Unit T2-11 in Zone B ( and ). This trench represents a series of interleaved cultural, ashy, and marl layers in a midden where it is likely that extensive landscaping activities contributed to its formation (see and ). This trench initially measured 1 × 1 m and was excavated by the University of the West Indies (UWI) Mona Archaeology team in the summer of 2018 to approximately 1.5 m in depth. The JNHT expanded this excavation to a 2 × 2 m unit in 2019 and extended excavations to a depth of approximately 2.2 m. At the time of sampling, the profile represented a full sequence from the bedrock extending over 2 m to the modern ground surface. No human remains were recovered from this excavation unit, but three human burials were documented approximately 3–4 m to the east in neighboring units (U2-11-NW, U2-12-NE).
Figure 3. Archaeobotanical phytolith sampling, White Marl. (A) view of archaeological excavation in Sampling Unit T2-11; (B) recording and sampling stratigraphy in Unit T2-11; (C) stratigraphic profile of Unit T2-11 sampled for phytoliths (21 samples in total) and charcoal for C14 dating; (D) schematic of the 21 layers sampled for phytolith analysis (for description see ). Black boxes indicate phytolith sample locations and red boxes indicate charcoal samples taken for C14 dating.

Table 2. Details of samples taken for phytolith analysis; sample number, description, Munsell color, Munsell description, and sample depth.
The soil profile was cleaned in July 2019 and the visible stratigraphy was recorded and sampled for phytoliths using standard procedures (Piperno Citation2006) (). A description of the stratigraphic layers is detailed in . Samples were collected from each identified stratum using sterile protocols (Goldberg and Macphail Citation2006, 328) and the Munsell color recorded. For comparative purposes, a small trench was also excavated away from the archaeological units at White Marl, approximately 100 m from Unit T2-11 (see ), into a natural soil profile encapsulating the A and B horizons which were observed as sterile deposits. Five comparative control samples were taken at 10 cm intervals to a depth of 60 cm (see SI for details of control pit samples).
Phytoliths
Silica phytoliths are a micro-botanical proxy composed of opaline silica that forms in and around plant cells which are well preserved in archaeological sediments. They survive in most soil conditions, only degrading in extremely alkaline soils or when burned at very high temperatures (e.g., from 600 to 900 °C; Devos, Hodson, and Vrydaghs Citation2021). Phytoliths represent a localized vegetation signature because, unlike many pollen types, they have limited transportation dispersal by wind and water and therefore are usually assumed to occur in situ (i.e., within <5 m2 of original deposition location) (Shillito Citation2018).
Phytolith analysis has been successfully integrated in archaeological contexts with macro-charcoal, stratigraphic analyses, and radiocarbon dating, as well as integrating with pollen analysis from lake cores which helps reconstruct the environment. This multi-method integration of proxies is the recommended standard approach to paleoenvironmental investigations (Iriarte et al. Citation2020; Robinson et al. Citation2021). Much work has been conducted in South America on a wide range of domesticated crops including squash, maize, manioc, and rice (Ezell, Pearsall, and Zeidler Citation2006; Hilbert et al. Citation2017; Iriarte et al. Citation2020; Piperno and McMichael Citation2020), but has not yet become standard practice in the Caribbean.
Phytolith extraction
The phytoliths were extracted using a modified methodology combining Piperno (Citation2006) and Rosen (Citation1999), utilizing a combination of 24 h disaggregation and dry ashing. Approximately 25 g of each sample was sieved through a 500-micron mesh and disaggregated with sodium hexametaphosphate (5%) on an orbital shaker for 24 h prior to clay removal using a settling procedure at hourly intervals. Carbonates were then removed using 10% hydrochloric acid (HCl), this was followed by removal of organics using dry ashing in a muffle furnace for 3 h at 500 °C. The phytoliths were then separated using sodium polytungstate heavy liquid calibrated to 2.3 specific gravity, and then dried and 200 mg mounted onto glass slides using the mounting medium Entellan.
Phytolith identification, counting, and quantification
Phytoliths were counted and identified at ×400 magnification on a Meiji MT4300L transmitted light microscope and photographed using a Canon EOS 1100 D SLR and EOS Utility software. Phytoliths were compared to the University of Exeter’s guide and reference collection for identification, in addition to a range of published studies (Albert et al. Citation1999; Dickau et al. Citation2013; Ezell, Pearsall, and Zeidler Citation2006; Morcote-Ríos, Bernal, and Raz Citation2016; Ollendorf, Mulholland, and Rapp Citation1988; Piperno Citation2006; Twiss, Suess, and Smith Citation1969; Watling and Iriarte Citation2013; Watling et al. Citation2016). For each sample, a minimum of 200 identifiable phytoliths were recorded where possible; for some samples this minimum count could not be reached due to extremely low phytolith concentrations (this was not possible for two of the 24 samples). Not all phytoliths can be identified further than Class (i.e., monocotyledon or dicotyledon), but where possible, the phytoliths were assigned to family, sub-family, and genus and also grouped into four main categories for analysis: arboreal, palms, grasses/herbs, and crops. The results are presented as percentages of the total morphotypes identified and graphed using C2 software (Juggins Citation2007). In addition, degraded, melted, burned, fused, and unidentified phytoliths were recorded, as well as the presence of diatoms and sponge spicules.
We used the phytolith analysis to document landscape modifications and indicators of management, establish the drivers for these changes, and present evidence of crops incorporated into the diet, and furthermore examine any evidence that may indicate dietary or subsistence change over time. This is conducted by recognizing a range of staple crops that can be identified from their phytoliths (e.g., squash, manioc, maize, rice), which might be grown in the vicinity of the site and incorporated into the diet.
Charcoal extraction and quantification
Subsamples of 1 cm3 were taken using a 5 cm3 syringe and sorted into 15 ml test tubes. To disaggregate the charcoal particles from the sediment, the samples were placed in a hot water bath (80 °C) with 45 ml of 5% potassium hydroxide for 15 mins. The samples were washed in a 125 μm sieve until clean then transferred into petri dishes submerged in water for analysis. All charcoal particles >125 µm were counted in the analysis.
Results
This section presents the results of the archaeological samples from White Marl; for a summary of the control samples please see the SI. The concentration of phytoliths in 19 of the 21 archaeological samples analyzed was very high (see SI). However, the two deepest samples contained very few phytoliths in comparison to the samples in the upper stratigraphic layers of the soil profile, and a minimum number of phytoliths was therefore not obtained even when the whole slide was observed (see SI).
A total of 24 single phytolith morphotypes and six multi-cellular phytolith forms were identified in the samples from White Marl and the most common types are presented in . The results of the phytolith analyses for the White Marl archaeological profile T2-11 in Zone B (western profile) () can be divided into three main vegetation phases (), with Phase 1 representing the deepest and oldest samples (192–224 cm), Phase 2 representing the samples in the center of the profile (93–185 cm), and Phase 3 representing the newest deposits at the top of the profile (5–87 cm). Phase 1 and Phase 2 are dominated by canopy cover phytoliths (arboreal and palms combined) in the oldest levels. The canopy cover phytoliths represent at least 50% of the phytolith assemblage from the base of the profile until 78 cm below ground level (pre-ca. 670 cal yrs BP). In Phase 3, from ca. 670 cal BP, the canopy cover reduces to an average of 29% of the phytoliths originating from canopy vegetation (combined arboreal and palm).
Figure 4. Common and distinctive phytolith forms observed in White Marl archaeobotanical samples, morphotypes listed indicated by arrows: (A) arboreal globular granulate (S7, 160 cm); (B) Arecaceae globular echinates (S17, 52 cm); (C) Poaceae elongate psilate conjoined (S5, 185 cm); (D) Aristoideae bilobe (S21, 5 cm); (E) Bambusoideae spiked rondel (S13, 93 cm); (F) Chloridoideae saddles (S21, 5 cm); (G) Panicoideae bilobe (S17, 52 cm); (H) Pooideae rondel (S21, 5 cm); (I) Phragmites conjoined stacked bulliforms (S17, 52 cm); (J) Manihot sp. secretory cell phytoliths (S13, 93 cm); (K) Cucurbita (squash) scalloped sphere (S17, 52 cm); (L) Oryza (rice) double peaked glume (S20, 15 cm); (M) Calathea rhizome cylinder (S20, 15 cm); (N) Unidentified fused phytoliths (S9, 150 cm); (O) Burnt occluded carbon Poaceae keystone (S21, 5 cm).

Figure 5. Results of the phytolith analysis from Unit T2-11, White Marl. Results expressed as a percentage of the total phytolith morphotypes identified. Charcoal (in black) is presented as overall counts per 1 cm3 > 125 um. Radiocarbon dates at depths of 190 and 90 cm. Hashed lines indicate main vegetation phases. Overall data summary indicative of three main vegetation phases highlighted by gray box.
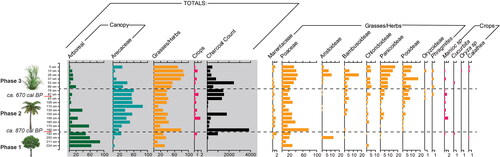
Vegetation phase 1: Arboreal-dominated canopy
In the deepest levels, the phytoliths extracted from the sediment samples suggest that the oldest vegetation in this area of the site was probably dominated by an arboreal canopy (224–192 cm, layers 18–21) with an average of 65% arboreal phytoliths recovered (and only 8% palm on average, pre-ca. 870 cal yrs BP). The phytolith remains in these layers represent the dominant deposition of arboreal plant remains in these midden contexts and as such, the phytoliths likely represent the vegetation at the time of the early occupation at White Marl.
Vegetation phase 2: Palm-dominated canopy
From ca. 870 cal yrs BP, there was a transition to a palm-dominated canopy (185–78 cm, layers 5–15) with an average of 48% palm phytoliths (and only 14% arboreal on average) which continues throughout Phase 2 until ca. 670 cal yrs BP. Palm species are not only useful industrially in construction, but many species are also edible. This increase in palms can be attributed to an increase in some of these useful species at the site which have only been identified from the second vegetation phase (185 cm upwards). In Phase 1, only low numbers of globular echinate palm phytoliths were identified, which are not attributed to edible species of palm. In Phase 2 the globular echinate palm phytoliths increase in addition to the identification of globular echinate-hat type phytoliths which are produced in nine edible species of the genus’ Astrocaryum and Bactris (Bozarth et al. Citation2009). In these samples the phytoliths probably originate from Bactris jamaicana which is native to Jamaica (Adams Citation1972); the three species of Bactris restricted to the Caribbean are Bactris plumeriana, Bactris jamaicana, and Bactris cubensis (Salzman and Judd Citation1995).
Vegetation phase 3: Open grassland-dominated landscape
The vegetation surrounding the site was further altered with a transition to an open grass-dominated landscape (69–5 cm, layer 16–21) from ca. 670 cal yrs BP onwards, with an average of 71% grass phytoliths (only 10% arboreal and 19% palm on average). While some of the typical surface disturbance indicators were not identified in the samples (e.g., Asteraceae) the reduction in canopy indicators is clear; the combined palm and arboreal canopy in these levels was only represented by an average of 29% canopy cover phytolith morphotypes in this final vegetation phase. This final shift in vegetation roughly coincides with the emergence of Taíno chiefdoms in the northern Caribbean around 750 cal yrs BP. All grass sub-families show an increase in Phase 3, but the phytolith assemblages are dominated by panicoideae grass phytoliths. Domesticated sugarcane, genus Saccharum is not known to produce distinctive phytoliths, but is a panicoid grass, and sugarcane is introduced only after 440 BP, and therefore could be responsible for the increase in panicoideae phytoliths in these upper layers in Phase 3. At ca. 670 cal yrs BP, Phragmites phytoliths also appear in the phytolith assemblages. There is a small peak in Arecaceae in the uppermost sample (5 cm), although the upper 15 cm of the profile is likely affected by plow disturbance and modern vegetation clearance for the archaeological investigations that have been on-going since the 1960s.
Crops
Phytoliths from four crops were recovered in our samples from profile T2-11 at White Marl: manioc, squash, Oryza sp. (rice), and leren/arrowroot (see ). Manioc was identified from 10 of the samples at White Marl, initially at 192 cm (pre-ca. 870 cal yrs BP). The majority of the manioc was recovered during the second vegetation phase dominated by palms ca. 870–60 cal yrs BP (manioc was recovered in 7/11 samples in this phase). Squash was identified from two samples at a depth of 192 cm, ca. 870 cal yrs BP, and at 52 cm after ca. 670 cal yrs BP. Rice was only identified in one sample after ca. 670 cal yrs BP (15 cm depth) which is within the plow zone and likely colonial in origin. Leren (arrowroot) was identified in three samples, before ca. 870 cal yrs BP (202 cm depth), and in two of the latest samples after ca. 670 cal yrs BP (5 cm and 15 cm, respectively). Maize phytoliths were not identified from these samples analyzed. Overall, the crops identified represent a small percentage of the phytolith assemblages; however, we know that some of the crops are extremely low producers of phytoliths (e.g., manioc, see Ezell, Pearsall, and Zeidler Citation2006) so may be underrepresented. In the earliest arboreal vegetation phase, the crops represented only 0.2% of the phytolith assemblage. The highest percentage of crops occurred in the second phase of vegetation dominated by a palm canopy (0.5% crops on average), with only manioc recovered during this phase. In the latest grass-dominated phase, the crops represented an average of 0.4% of the phytolith assemblage.
Burning indicators
There is a high number of unidentifiable “fused” phytoliths in the samples (), particularly in the second vegetation phase which was dominated by palms ( and ). These types were also observed ubiquitously in archaeological deposits at two contemporaneous sites in the Dominican Republic (see Pagán-Jiménez et al. Citation2020). These unidentifiable fused phytoliths, or pyrolyzed phytoliths, show traces of melting or burning probably relating to lower temperature burning. The pyrolytic process is a slow combustion carried out at low oxygen supply, between 300 and 700 °C (Boateng et al. Citation2015; Li and Delvaux Citation2019) . These “fused phytoliths” identified at White Marl have been interpreted as an indicator of past burning activities. During sampling, many layers were observed as visibly burned or ashy (). The number of fused phytoliths generally peak at the same depths as peaks in the soil charcoal counts (, with the exception of 52 cm). Occluded carbon burned phytoliths are an additional proxy for fire activity and were also observed throughout the profile from 13 of the 21 samples. The peaks in burned phytoliths do not directly relate to peaks in fused phytoliths and charcoal; however, these peaks are broadly stratigraphically related (always within 15 cm of the charcoal and fused phytolith peaks; see ). In the deepest stratigraphic layers, there were no fused phytoliths and minimal charcoal (five and six charcoal fragments in samples from 224 and 211 cm; ). The first peak in charcoal and fused phytoliths coincides with the vegetation transition from arboreal- to palm-dominated canopy and a concomitant increase in grasses (185 and 192 cm). It also coincides with the first visible appearance of ashy material as described in the field descriptions of the stratigraphy (see sample 5, , and ). These repetitive thick layers of ash, which often contain mollusks, pottery, and faunal remains (first occurrence at 185 cm; stratigraphic/sample 5, then at 170, 150, 93, 52 cm, see ) are separated by unconsolidated marl sediments sometimes containing small rocks or cobbles (at 160, 156, 130, 108, 69, and 37 cm; see ).
Figure 6. Grape Tree Pond climate record (Burn and Palmer Citation2015) (also indicating the Medieval Warm Period (Medieval Climate Anomaly, MCA) and Little Ice Age (LIA)) plotted against burning proxies (charcoal counts, fused unidentifiable phytoliths, burnt occluded carbon phytoliths) and vegetation summaries (arboreal, palms, grasses/herbs), with percentage of manioc and new White Marl radiocarbon dates (at 90 cm and 190 cm). The Grape Tree Pond record has been cropped between 650 and 500 cal yrs BP to reflect the age resolution of the soil samples. Hashed lines indicate transitions between vegetation phases 1, 2 and 3. Av = averages for each phase for burning indicators and manioc.
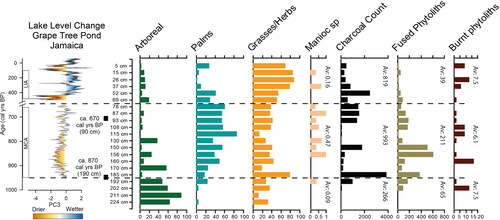
At White Marl within the palm-dominated vegetation phase (Phase 2), fire indicators from the phytoliths and charcoal peak in similar locations in the profile, but are not perfectly correlated. This may be due to post deposition taphonomy. There are some coincidental peaks in charcoal and peaks in fused phytoliths. At 150 cm these include: 509 fused phytoliths, and 1,806 charcoal fragments; and at 87–93 cm: 136–185 fused phytoliths and 1,358–1,545 charcoal fragments (). In these locations either ash or charcoal was noted in the field descriptions (see , samples 9, 13, 14). On average, the highest number of fused phytoliths and charcoal fragments occurred during the palm-dominated vegetation phase with 211 fused phytoliths and 993 charcoal fragments (see ).
In the final grass-dominated vegetation phase (Phase 3), there was a general trend toward reduced numbers of both fused phytoliths and charcoal. However, even though there is a reduction in burning proxies extracted from the sediment samples, charcoal was observed in the profile description in sample 17 (52 cm; see ), which corresponded to a peak in macroscopic charcoal (2,455 charcoal fragments at 52 cm; see ). Additionally, charcoal was observed in the profile description in sample 19 (26 cm) and there was a peak in burned phytoliths (n = 13 at 26 cm; ).
Vegetation changes and climate
The main vegetation phases and burning indicators from White Marl have been plotted against climatic data for lake level change at Grape Tree Pond (Burn and Palmer Citation2015, ) and the predominant climate patterns of the Medieval Climate Anomaly (MCA) and Little Ice Age (LIA) (). Currently, a direct comparison between the two records is difficult given the different temporal resolution of both datasets and that our sequence from White Marl is reliant on, and therefore constrained by, only two radiocarbon dates. However, in general, there appears to be some relationship between climatic variability and the main changes in dominant vegetation types identified between our phases in the phytolith data. The climatic record indicates a wetter environment in the LIA, where there is a concomitant increase in grass and decrease in palm phytoliths ().
The mosaic of grasses in Phase 3 include increasing C3 pooideae grasses, which prefer cold wet growing environments, and the continued low level of C4 chloridoideae grasses that prefer dry environments. Panicoid grasses also increase in Phase 3 (LIA; see and ), which grow in wet and warm conditions.
There is an emergence of Phragmites (reeds) at the transition from Phase 2 to 3 (at 87 cm; ) and persistence in Phase 3 (15 cm, 37 cm and 69 cm; ) may be a consequence of climatic change, with a general trend toward wetter conditions in the LIA.
Discussion
Despite a growing number of studies examining human–environment interactions in the Caribbean (Berman and Pearsall Citation2020; Castilla-Beltrán et al. Citation2018; Ciofalo, Sinelli, and Hofman Citation2020; Mickleburgh and Pagán-Jiménez Citation2012; Pagán-Jiménez et al. Citation2020; Piperno Citation2002; Sara and Aguilú Citation2003; Siegel et al. Citation2015), to date there is limited archaeobotanical evidence from Jamaica. The current assumption is that the Indigenous occupants removed and altered the natural vegetation in order to establish settlements and areas for cultivation, and to create useful productive subsistence landscapes (Atkinson Citation2006). On other Caribbean islands, the Indigenous populations in the Late Ceramic Age exploited diverse environments relying on agriculture and fishing with the settlements increasingly shifting away from coastal areas to hilltop and riverine locations (de Armas et al. Citation2015; Fitzpatrick and Keegan Citation2007; Hofman et al. Citation2018; Keegan and Hofman Citation2017; Pagán-Jiménez et al. Citation2020; Sonnemann, Ulloa Hung, and Hofman Citation2016). At White Marl, like most other sites on the island, the emphasis has traditionally been placed on the recovery of faunal remains, and while charcoal has previously been collected from the White Marl contexts for dating, no interpretations pertaining to past human–vegetation–fire linkages have been conducted. Three phases identified from the phytolith and charcoal analyses characterize the human-induced interactions with the environment at White Marl which include intentional burning (see ).
Figure 7. Diagrammatic representation of three vegetation phases at White Marl, as indicated by phytolith and charcoal results.
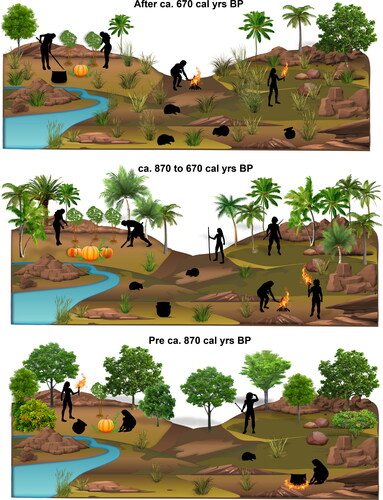
In this study, the earliest phytolith data in the deepest layers of the sampled profile (e.g., at 224 and 211 cm)—which were identified visually as the eroded natural geologic material (C horizon/eroded parent material)—suggest sparse vegetation deposition in this location. This is indicated by the lack of phytoliths (less than statistically viable for analyses), the absence of fused or burnt phytoliths, and extremely limited charcoal fragments, suggesting minimal fire activity at the site during this time. The phytoliths that were identified in the earliest samples in Phase 1 were dominated by arboreal forms with some input of grasses. This material is likely the first organic component within the soil matrix and represents an established forest community in this location, with similarities to other sites in the Caribbean (Pagán-Jiménez et al. Citation2020, 9). Together, these data indicate a signature of naturally occurring vegetation, with little to no anthropogenic influence in this location of the site in the earliest sampled levels, suggesting that these deposits, which sit directly on the bedrock, perhaps pre-date human occupation at White Marl that is thought to have taken place ca. 1050 cal yrs BP. Furthermore, this signal is supported by the absence of ceramic materials recovered in the deepest context in this unit (sample 1; ), the first ceramics being recovered at ca. 210 cm depth in the transition between stratigraphic units 2 and 3 (see ).
In the overlying strata, there is evidence for an actively managed landscape in the form of human modifications and management of the vegetation. This is represented in the phytolith assemblages by three main vegetation phases and their associated fire proxies. This repetitive sequence of marly sediment and interspersed ashy layers at White Marl have been observed at contemporaneous Meillacoid sites in the Dominican Republic where the “layers of fine ash” were separated by “whitish gravelly fill,” which has been interpreted as leveling material (Pagán-Jiménez et al. Citation2020). These sites in the Dominican Republic have similar complex engineered landscapes with cleared and leveled areas for house construction (Pagán-Jiménez et al. Citation2020, 4). The origin of the whitish gravelly sediment has been interpreted as deriving from leveling activities during house construction, and the ashy layers deposited either from the cooking huts, or developed in situ as active kitchen areas (Pagán-Jiménez et al. Citation2020, 4). However, these ash layers could have formed during localized tree clearance across these sites. Furthermore, in the Dominican Republic sites, burials were recovered from these deposits, which can also be seen at White Marl. The formation of these ash/midden deposits with burials incorporated into them appears to be a characteristic both at this site and across the region, which could suggest the importance of the interaction between humans and their surrounding environment (Allsworth-Jones Citation2008; Stokes Citation2002).
Initially an arboreal-dominated vegetation canopy around White Marl is indicated by the phytolith assemblage pre-ca. 870 cal yrs BP (Phase 1). The initial vegetation modification from ca. 870 cal yrs BP was associated with the expansion of the palm-dominated canopy (Phase 2), including the introduction of at least one species of edible palms (Bactris) at the expense of arboreal tree cover, in addition to increased manioc and peaks in fire proxies. In this palm-dominated phase the increase of fused phytoliths and charcoal suggests that during this time there was increased burning, or increased deposition of burned materials at the sampling location, particularly of palm vegetation. The use of palm trees as a fuel, as is represented by the phytolith remains in these deposits, suggests that high numbers of trees were being removed and burned, and probably therefore represent local vegetation. This shift in vegetation in Phase 2 to an increased palm canopy indicates probable anthropogenic changes in vegetation composition around White Marl. Palms are frequently utilized around the world in antiquity and modern times, both in rural construction and other human activities such as rituals (Jarzombek Citation2013, 99, 111, 562).
Crop phytoliths increase in Phase 2 also suggesting anthropogenic influence. As these are assumed to be consumed, and we see an increase through time, the hypothesis is that they are domesticated crops, although the phytoliths have only been identified to genus (with the exception of Calathea). There is a possibility, however, that these crops were collected as wild species from around the site.
The new phytolith data suggest that inhabitants of White Marl from ca. 870 cal yrs BP practiced agroforestry, increasing the concentration of useful palms and crops in and around the site. This is similar to contemporaneous sites in the northern Dominican Republic where similar interspersed ashy and marl layers are observed, and agroforestry was also interpreted from the phytoliths (Pagán-Jiménez et al. Citation2020). The phytolith data indicate clear instances of palm enrichment and predominance alongside crops (e.g., Zone FA-II, El Flaco). It was suggested that occupants at these sites in the Dominican Republic were consciously increasing food plant production through agroforestry practices and a clear cultural sustenance of palms (Pagán-Jiménez et al. Citation2020, 8–12). In contrast to the inhabitants at these two sites, the inhabitants of White Marl appear to have dramatically affected general forest and vegetation composition. Significantly, at White Marl, the final phase after ca. 670 cal yrs BP (Phase 3) represents a more open grassland-dominated vegetation with continued representation of crops.
The latest deposits in the sequence are probably affected by disturbance and therefore also represent colonial and modern-day occupation, a more open environment with lower levels of trees, but continued evidence for low-level burning. Burning continued at a reduced rate in Phase 3, suggesting that there was more limited burning or lower amounts of burned materials deposited in this final phase where there are fewer trees, with the phytoliths indicative of a more open grassland. The reduction in burned materials could result from availability of materials, a change in fuel practices or an alternative function/use of this area of the site. The dominant phytoliths in this phase (grasses) could reflect the surrounding natural vegetation. An increase in grasses and reduction in canopy would impact the availability of fuel and therefore intensity of burning. Ethnographic and ethnohistorical studies in both North and South America show sedentary agricultural communities relocating after around 15–20 years of occupation due to diminished fertility in agricultural soils and increased distance to firewood. These visual observations, and peaks in burning proxies in Phase 3 at White Marl, seem to represent some continuation of burning and potential kitchen activities throughout the final occupation phase at White Marl.
In addition to the three broader transitions in vegetation cover, the phytolith data from this study provide new insights into changes in past subsistence strategies at White Marl. Arecaceae (palm) phytoliths were identified in all samples, but edible palms are only represented with certainty from the second vegetation phase onwards (from ca. 870 cal yrs BP) indicating a human-driven change in the landscape which relies more on palms, including edible palms.
A range of plants that were edible and most likely consumed by the White Marl inhabitants were identified from the phytoliths at White Marl: manioc, squash, rice, and leren/arrowroot. This is unlikely to represent the full suite of edible species consumed at White Marl based on dental calculus evidence (Mickleburgh et al. Citation2019), evidence found at contemporary sites on other Caribbean islands (e.g., Pagán-Jiménez et al. Citation2020), and what is known about precolonial crop plant use in the region generally (Newsom Citation1993; Newsom and Wing Citation2004).
Manioc was the dominant crop identified in the phytolith profile at White Marl and was once assumed to be the dominant crop in the precolonial Caribbean (Berman and Pearsall Citation2020; Howard Citation1956; Parry Citation1955; Pope et al. Citation2001; Rouse Citation1992; Vanderwal Citation1968). Early assumptions of manioc cultivation and consumption at White Marl were inferred from ceramic artifacts which are identical in form to the griddles used in the Caribbean for baking manioc bread at the time of European conquest (Silverberg, Vanderwal, and Wing Citation1972). However, griddles have long been shown to have been used for a variety of starchy plants, and are no longer considered de facto indicators for manioc consumption (Berman and Pearsall Citation2020; Ciofalo et al. Citation2018; Ciofalo, Sinelli, and Hofman Citation2019; DeBoer Citation1975; Rodríguez Suárez and Pagán-Jiménez Citation2008). In addition, the analysis of stone tools from the insular Caribbean and French Guiana recovered manioc starch from only 10% of the stone tools analyzed (Pagán-Jiménez Citation2011). Furthermore, the region-wide study of starch grains extracted from human dental calculus samples spanning ca. 2300–350 cal yrs BP found that the existence of a sole staple food plant in the precolonial Caribbean is unsupported, refuting traditional narratives that manioc-dominated plant-based subsistence in the region (e.g., Mickleburgh and Pagán-Jiménez Citation2012; Pagán-Jiménez and Mickleburgh Citation2022).
In the past, manioc had proved to be elusive in the phytolith record; however, manioc produces low numbers of distinctive heart-shaped phytoliths (pectate forms) from the secretory cells in the root, leaf, and fruit of the genus Manihot (Ezell, Pearsall, and Zeidler Citation2006). The new phytolith results presented here have identified manioc in half of the layers from this excavation unit, suggesting that it may have been an important crop at White Marl. We assume that based on its importance as a food crop, and its recovery in an environment with significant anthropogenic influence, that manioc was indeed being consumed.
In the light of this finding, we emphasize the importance of integrating multi-proxy datasets such as dental calculus, stone tools, pottery residues, and macro-botanicals to develop a more holistic understanding of crop cultivation and consumption. For example, maize starches were identified in the dental calculus (Mickleburgh et al. Citation2019) but no maize phytoliths were identified from the sediment phytoliths, and vice versa for manioc. The small size and low concentration of distinctive manioc phytoliths could result in their misidentification or absence from other phytolith studies. The disparity in the results from a range of different proxies highlights the importance of adopting an interdisciplinary approach. The combined results indicate that manioc was grown and probably consumed at the site or close to the site, whereas the absence of maize in the phytolith profile suggests it may have been grown either in a different location within the site, or grown away from the site and brought in and consumed. Perhaps maize was grown in the lower flood plains closer to the Rio Cobre where the soil was better suited for growing this nutrient demanding crop (Crawford et al. Citation1998). Alternatively, its absence in these layers could have resulted from artificial landscape modifications and the movement and reworking of sediments at White Marl intentionally by the occupants (i.e., the deposits where maize was grown could have been removed and deposited elsewhere on site, in locations not sampled by this pilot study). Only one stratigraphic profile was analyzed and therefore further testing at the site might clarify the presence and absence of certain crops.
The absence of manioc in the dental calculus is unsurprising as the numbers of phytoliths and starch grains extracted from calculus are often small (often only dozens rather than hundreds; see Cummings, Yost, and Sołtysiak Citation2018; Piperno Citation2006, 163), and sometimes only single starch grains or phytoliths are recovered per tooth (e.g., Henry Citation2011). So, while the results can provide dietary insight, the remains do not fully represent the breadth of what is consumed or the proportional contribution of each plant. Cultivation of starchy staple crops could occur in isolated areas which have been recognized from similar sites as “home or kitchen gardens” (de Las Casas Citation1909; Newsom and Wing Citation2004; Pagán-Jiménez et al. Citation2020). However, the wider study of contexts from across the site is required to support this interpretation at White Marl.
The first appearance of manioc in the phytoliths and the subsequent peaks in manioc phytoliths broadly coincide with increases and peaks in charcoal and burned and fused phytoliths, suggesting fire was present and possibly used as a tool to modify the landscape and clear vegetation for crop cultivation. The area around White Marl is naturally dry and susceptible to fire as it is located in the rain shadow of the Blue Mountains, and it seems reasonable that peoples would select fire resilient crops. Manioc tubers are well-adapted to fire and are often referred to as a “disturbance adapted plant” (Pujol et al. Citation2002). The tuberous roots store underground reserves, enabling rapid regrowth after fire and other disturbances (Rival and McKey Citation2008). Studies have shown that when fields are cleared manioc seeds still remain. When these fields are subsequently burned it stimulates manioc germination approximately two weeks after the burning occurs (Pujol et al. Citation2002). Some research suggests that domesticated manioc could have inherited a germination biology that was “pre-adapted” to shifting cultivation and is particularly well suited to habitats in which fire is used, such as in slash and burn (Pujol et al. Citation2002, 367, 375). At White Marl, the contexts sampled and analyzed in this study represent midden areas and may have been affected by landscape reworking. Without further investigation, however, we cannot rule out whether there were discrete areas of cultivation within the site itself. The identification of manioc from the phytoliths throughout the record coinciding with a decline in arboreal taxa, combined with peaks in the fire indicators (charcoal, fused and burned phytoliths), suggests that the manioc–fire linkages were key components of the land use strategy and subsistence strategy at White Marl.
The variety of crops represented by the phytoliths increased after ca. 670 cal yrs BP during the third phase of vegetation which was dominated by open grassland; however, these assemblages are likely influenced by plowing and the colonial and modern-day occupation at White Marl, with manioc, squash, and leren (arrowroot), and European introduced rice, identified in all of the most recent levels.
Phytoliths from the Zingiberales order were also recovered, specifically from the family Marantaceae. The only genus within this family that was positively identified from the phytoliths extracted from the sediment samples was Calathea (arrowroot). The tubers of leren/arrowroot are edible and it is classed as a minor food crop in precolonial times (Iriarte et al. Citation2020). Arrowroot tubers, like manioc, are fire adapted and the selection of this crop seems logical. Starch has been recovered from dental calculus samples taken from other sites in the insular Caribbean (de Armas et al. Citation2015; Mickleburgh and Pagán-Jiménez Citation2012) and starch produced by the family Marantaceae and genus Calathea have been identified from Guadeloupe ca. 1500–600 cal yrs BP (Mickleburgh and Pagán-Jiménez Citation2012). Maize and wild beans (Fabaceae) were not identified from the sediment phytolith analysis at White Marl, although they were identified in the dental calculus from burials at White Marl, which evidenced the consumption of maize, wild beans, and possible Theobroma cacao (cocoa) (Mickleburgh et al. Citation2019).
The modifications in vegetation composition appear to correlate with climatic shifts () though the response is the opposite to what might be predicted. We would expect any increase in precipitation versus evaporation during the LIA to have increased the relative percentage of closed-canopy trees and palms versus grasses (Davis et al. Citation1997; Grossman, Iremonger, and Muchoney Citation1993). Indeed, wet conditions on the northeast coast of Jamaica today are, in part, responsible for the presence of contemporary continuous closed-canopy rainforest communities. Given that the response of the local vegetation contradicts what we would expect if climate were the driving force, we conclude that human activity was the cause of the observed changes in vegetation. Within the mosaic of increased grasses in Phase 3, C4 chloridoideae, C3 pooideae, and C4 panicoideae grasses were identified. The panicoid grasses grow in wet and warm conditions, and so while the overall vegetation composition is not what would be expected for these climatic conditions, the composition of the grass assemblage (more panicoid than pooideae grasses) may correlate with precipitation and climatic shifts.
The increase in panicoid grasses in Phase 3 could also be related to the introduction of sugarcane to the area after 450 BP. Domesticated sugarcane (genus Saccharum), is a panicoid grass which almost doubles at the beginning of Phase 3 and further increases during this phase, suggestive of sugar cultivation in the area. The slight drying conditions likely made this area suitable for sugarcane cultivation and livestock farming and colonizers’ exploitation of the area could have led to the clearance of the canopy for this purpose. This grass-dominated ecosystem persists to the present day, suggesting that the historic land use practices associated with industrial agriculture, alongside present-day denser habituation in the area, had a lasting legacy on the modern vegetation at White Marl.
The emergence of Phragmites, which is a wetland species that grows in year-round wet environments (at the end of Phase 2 and in Phase 3) could reflect a trend toward a wetter climate which is represented in the Grape Tree Pond climate record (Burn and Palmer Citation2015). Alternatively, the presence of these materials, which are well documented in many areas of the world as common materials in construction, textiles, and boat building (Jarzombek Citation2013, 285–6), may have resulted from human selection.
Conclusion
The results from micro-botanical analyses at White Marl provide evidence for distinctive signals of human–environment interactions in this area of the site. We have shown that the local vegetation at White Marl was directly impacted by human settlement. Overall, the new phytolith results indicate that there was a diverse polyculture subsistence strategy at White Marl which is in keeping with what is known from botanical studies at other Meillacoid sites in the region (e.g., Pagán-Jiménez et al. Citation2020).
Here we have shown evidence for temporal vegetation change driven by humans rather than the climate. The evidence from White Marl shows a transition from the pre-human floral environment encountered at initial colonization of pre-ca. 870 cal yrs BP, through subsequent European colonial occupation and sugar cultivation after ca. 670 cal yrs BP. In this study, we identified three clear phases of vegetation change which reflect human-driven environmental modifications. This includes the increasing importance of palms and first evidence for persistent manioc remains throughout occupation of White Marl. The evidence for the relationship between burning and manioc suggests that fire may have been used as a tool to modify the landscape, clear vegetation for crop cultivation, and likely to promote manioc germination. Based on the economic importance of this crop, we can assume it was incorporated into the diet alongside terrestrial protein as indicated by the White Marl faunal remains, which include hutia (Geocapromys brownii), land crab, and reptiles (Wing Citation1972), in addition to some marine resources likely sourced from Kingston harbor (Mickleburgh et al. Citation2019). More research is currently underway to explore the relationships between marine, terrestrial, and agricultural resource management, which includes stable isotope analysis on remains of the Jamaican Hutia or Coney (Shev, Laffoon, and Hofman Citation2021) and new lake core records from the island that can be compared with White Marl.
Later, polycrop cultivation was also identified at White Marl, including squash, leren, and eventually rice in the colonial occupation deposits. Our results show the consistent presence of manioc throughout the period of occupation at White Marl. Where manioc is absent in the starches extracted from dental calculus (Mickleburgh et al. Citation2019), it is deposited in sediments as evidenced by the phytolith remains. Various studies have shown that other starchy foods were equally or more important in the precolonial diet in the Greater Antilles (Berman and Pearsall Citation2020; Ciofalo et al. Citation2018; Ciofalo, Sinelli, and Hofman Citation2019; de Armas et al. Citation2015; Mickleburgh and Pagán-Jiménez Citation2012; Pagán-Jiménez et al. Citation2020; Pagán-Jiménez and Mickleburgh Citation2022). Increasingly, botanical evidence in the region points to no single staple plant or crop being utilized ubiquitously across contemporary sites, but rather the use of a range of crops which varied on a site by site basis (Pagán-Jiménez and Mickleburgh Citation2022). Further research at White Marl may reveal the role of manioc and other crops within the plant component of the diet.
The latest vegetation transition, dominated by increased canopy clearance and prevalence of grasses with peaks in fire proxies persisted to modern times, suggesting that the land clearance associated with industrial production of sugarcane (suggested by the increase in panicoid grasses), likely had the most significant impact on the landscape over a millennium, and was a persistent feature in modern vegetation. Agroforestry was initiated by increasing palms and crops and then eventually opening up the canopy where grasses increased prior to colonial occupation. The panicoid grasses (which include sugarcane) show a marked increase corresponding with the colonial plantation economy and industrial-scale land use that permanently transformed the local vegetation.
Supplemental Material
Download MS Word (430.6 KB)Acknowledgements
This is a joint program with the National Geographic Exploration Fund (PI Dr. Yoshi Maezumi), the Jamaica National Heritage Trust, and the Department of History and Archaeology at The University of the West Indies Mona. We would also like to thank and acknowledge Jonas Gregorio de Souza for producing , Harry Manley for assistance with , and Angus Mol for his input to the archaeological context of the Caribbean. Comments from several reviewers helped to improve various aspects of the manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Data availability statement
The authors confirm that the data supporting the findings of this study are available within the article and its supplementary materials (see SI online). This material has not been published elsewhere and has not been submitted simultaneously for publication elsewhere.
Additional information
Funding
References
- Aarons, G. A. 1983. Archaeological sites in the Hellshire area. Jamaica Journal 16 (1):76–87.
- Adams, C. D. 1972. Flowering plants of Jamaica. Mona, Jamaica: University of West Indies.
- Albert, R. M., O. Lavi, L. Estroff, S. Weiner, A. Tsatskin, A. Ronen, and S. Lev-Yadun. 1999. Mode of occupation of Tabun Cave, Mt Carmel, Israel during the Mousterian Period: A study of the sediments and phytoliths. Journal of Archaeological Science 26 (10):1249–60. doi:10.1006/jasc.1999.0355
- Allsworth-Jones, P. 2008. Pre-Columbian Jamaica. Tuscaloosa: University of Alabama Press.
- Atkinson, L. G. 2006. The earliest inhabitants: The dynamics of the Jamaican Taino. West Indies: University of West Indies Press.
- Bain, A., A.-M. Faucher, L. M. Kennedy, A. R. LeBlanc, M. J. Burn, R. Boger, and S. Perdikaris. 2018. Landscape transformation during ceramic age and colonial occupations of Barbuda, West Indies. Environmental Archaeology 23 (1):36–46. doi:10.1080/14614103.2017.1345115
- Berman, M. J., and D. M. Pearsall. 2020. Crop dispersal and Lucayan tool use: Investigating the creation of transported landscapes in the Central Bahamas through starch grain, phytolith, macrobotanical, and artifact studies. Journal of Field Archaeology 45 (5):355–71. doi:10.1080/00934690.2020.1740958
- Boateng, A. A., M. Garcia‐Perez, O. Mašek, R. Brown, and B. DelCampo. 2015. Biochar production technology. In Biochar for environmental management: Science and technology, ed. J. Lehmann and S. Joseph, 2nd ed., 64–8. London: Routedge, Eastscan Books Lts.
- Bochaton, C., S. Grouard, R. Cornette, I. Ineich, A. Lenoble, A. Tresset, and S. Bailon. 2015. Fossil and subfossil herpetofauna from Cadet 2 Cave (Marie-Galante, Guadeloupe Islands, FWI): Evolution of an insular herpetofauna since the Late Pleistocene. Comptes Rendus Palevol 14 (2):101–10. doi:10.1016/j.crpv.2014.10.005
- Boomert, A. 1999. Saladoid sociopolitical organization. In Proceedings of the XVIIIth International Congress for Caribbean Archaeology, 55–77. St George, Grenada: Association Internationale d’Archéologie de la Caraibe.
- Boudadi-Maligne, M., S. Bailon, C. Bochaton, F. Casagrande, S. Grouard, N. Serrand, and A. Lenoble. 2016. Evidence for historical human-induced extinctions of vertebrate species on La Désirade (French West Indies). Quaternary Research 85 (1):54–65. doi:10.1016/j.yqres.2015.11.001
- Bozarth, S., K. Price, W. Woods, E. Neves, and R. Rebellato. 2009. Phytoliths and terra preta: The Hatahara site example. In Amazonian Dark Earths: Wim Sombroek’s Vision, eds. W.I. Woods, W.G. Teixeira, J. Lehmann, C. Steiner, A. WinklerPrins, and L. Rebellato, 85–98. Berlin: Springer.
- Bright, A. J. 2011. Blood is thicker than water: Amerindian intra- and inter-insular relationships and social organization in the pre-Colonial Windward Islands. Leiden: Sidestone Press.
- Burn, M., and S. E. Palmer. 2015. Atlantic hurricane activity during the last millennium. Scientific Reports 5 (1):12838. doi:10.1038/srep12838
- Burney, D. A. 1997. Tropical islands as paleoecological laboratories: Gauging the consequences of human arrival. Human Ecology 25 (3):437–57. doi:10.1023/A:1021823610090
- Burney, D. A., L. P. Burney, and R. D. E. MacPhee. 1994. Holocene charcoal stratigraphy from Laguna Tortuguero, Puerto Rico, and the timing of human arrival on the Island. Journal of Archaeological Science 21 (2):273–81. doi:10.1006/jasc.1994.1027
- Caffrey, M. A., and S. P. Horn. 2015. Long-term fire trends in Hispaniola and Puerto Rico from sedimentary charcoal: A comparison of three records. The Professional Geographer 67 (2):229–41. doi:10.1080/00330124.2014.922017
- Callaghan, R. T. 2008. On the question of the absence of Archaic Age sites on Jamaica. The Journal of Island and Coastal Archaeology 3 (1):54–71. doi:10.1080/15564890801928615
- Castilla-Beltrán, A., H. Hooghiemstra, M. L. Hoogland, T. H. Donders, J. R. Pagán-Jiménez, C. N. McMichael, S. M. F. Rolefes, T. Olijhoek, E. Herrera-Malatesta, J. U. Hung, et al. 2020. Ecological responses to land use change in the face of European colonization of Haytí island. Quaternary Science Reviews 241:106407. doi:10.1016/j.quascirev.2020.106407
- Castilla-Beltrán, A., H. Hooghiemstra, M. L. Hoogland, J. Pagán-Jiménez, B. van Geel, M. H. Field, M. Prins, T. Donders, E. Herrera Malatesta, J. Ulloa Hung, et al. 2018. Columbus’ footprint in Hispaniola: A paleoenvironmental record of indigenous and colonial impacts on the landscape of the central Cibao Valley, northern Dominican Republic. Anthropocene 22:66–80. doi:10.1016/j.ancene.2018.05.003
- Chabanaud, P. 1946. Faune du gisement précolombien d'Anse-Belleville: Poissons et Oiseaux. Mémoires du Muséum 22 (2): 19–126.
- Chanlatte Baik, L. 1995. The archaic and the formative Antillean [Association Internationale Archeology of the Caribbean]. Proceedings of the International Congress for Caribbean Archaeology 16:267–74.
- Ciofalo, A. J., W. F. Keegan, M. P. Pateman, J. R. Pagán-Jiménez, and C. L. Hofman. 2018. Determining precolonial botanical foodways: Starch recovery and analysis, Long Island, The Bahamas. Journal of Archaeological Science: Reports 21:305–17. doi:10.1016/j.jasrep.2018.07.022
- Ciofalo, A. J., P. T. Sinelli, and C. L. Hofman. 2020. Starchy shells: Residue analysis of precolonial northern Caribbean culinary practices. Archaeometry 62 (2):362–80. doi:10.1111/arcm.12524
- Ciofalo, A. J., P. T. Sinelli, and C. L. Hofman. 2019. Late precolonial culinary practices: Starch analysis on griddles from the northern Caribbean. Journal of Archaeological Method and Theory 26 (4):1632–64. doi:10.1007/s10816-019-09421-1
- Clair, J. 1970. Problem orientated archaeology. Jamaica Journal 4 (1):7–10.
- Cochrane, M. A. 2009. Tropical fire ecology: Climate change, land use, and ecosystem dynamics. Chichester: Springer.
- Crawford, G. W., D. G. Smith, J. R. Desloges, and A. M. Davis. 1998. Floodplains and agricultural origins: A case study in south-central Ontario. Canada. Journal of Field Archaeology 25 (2):123–37.
- Cummings, L. S., C. Yost, and A. Sołtysiak. 2018. Plant microfossils in human dental calculus from Nemrik 9, a Pre-Pottery Neolithic site in Northern Iraq. Archaeological and Anthropological Sciences 10 (4):883–91. doi:10.1007/s12520-016-0411-3
- Curet, L. A. 1992. The development of chiefdoms in the Greater Antilles: A regional study of the Valley of Maunabo, Puerto Rico. PhD diss., Arizona State University.
- Davis, S. D., V. H. Heywood, O. Herrera-MacBryde, J. Villa-Lobos, and A. C. Hamilton. 1997. Centres of plant diversity: A guide and strategy for their conservation. Volume 3. The Americas. New York: The Worldwide Fund for Nature (WWF)/The World Conservation Union (IUCN).
- de Armas, Y. C., W. M. Buhay, R. R. Suárez, S. Bestel, D. Smith, S. D. Mowat, and M. Roksandic. 2015. Starch analysis and isotopic evidence of consumption of cultigens among fisher–gatherers in Cuba: The archaeological site of Canímar Abajo, Matanzas. Journal of Archaeological Science 58:121–32.
- de Las Casas, B. 1909. Apologética historia de las Indias: de Fr. Bartolome de las Casas [Apologetic history of the Indies: by Fr. Bartolome de las Casas]. Madrid: Bailly, Bailliere e hijos.
- DeBoer, W. R. 1975. The archaeological evidence for manioc cultivation: A cautionary note. American Antiquity 40 (4):419–33. doi:10.2307/279328
- Devos, Y., M. J. Hodson, and L. Vrydaghs. 2021. Auto-fluorescent phytoliths: A new method for detecting heating and fire. Environmental Archaeology 26 (4):388–405. doi:10.1080/14614103.2020.1777056
- Dickau, R., B. S. Whitney, J. Iriarte, F. E. Mayle, J. D. Soto, P. Metcalfe, F. A. Street-Perrott, N. J. Loader, K. J. Ficken, and T. J. Killeen. 2013. Differentiation of neotropical ecosystems by modern soil phytolith assemblages and its implications for palaeoenvironmental and archaeological reconstructions. Review of Palaeobotany and Palynology 193:15–37. doi:10.1016/j.revpalbo.2013.01.004
- Ezell, K. C., D. M. Pearsall, and J. A. Zeidler. 2006. Root and tuber phytoliths and starch grains document manioc (Manihot esculenta) arrowroot (Maranta arundinacea) and llerén (Calathea sp.) at the real alto site Ecuador. Economic Botany 60 (2):103–20. doi:10.1663/0013-0001(2006)60[103:RATPAS2.0.CO;2]
- Faegri, K., and J. Iversen. 1989. Textbook of pollen analysis. New York: John Wiley & Sons Ltd.
- Fitzpatrick, S. M. 2015. The pre-Columbian Caribbean: Colonization, population dispersal, and island adaptations. PaleoAmerica 1 (4):305–31. doi:10.1179/2055557115Y.0000000010
- Fitzpatrick, S. M., and W. F. Keegan. 2007. Human impacts and adaptations in the Caribbean Islands: An historical ecology approach. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 98 (1):29–45. doi:10.1017/S1755691007000096
- Giovas, C. M. 2018. Continental connections and insular distributions: Deer bone artifacts of the Precolumbian West Indies—A review and synthesis with new records. Latin American Antiquity 29 (1):27–43. doi:10.1017/laq.2017.57
- Goldberg, P., and R. I. Macphail. 2006. Practical and theoretical geoarchaeology. Oxford: Blackwell Science Ltd.
- Granberry, J., and G. Vescelius. 2004. Languages of the pre-Columbian Antilles. Tuscaloosa: University of Alabama Press.
- Grossman, D., S. Iremonger, and D. Muchoney. 1993. Jamaica: Map of natural communities and modified vegetation types. Jamaica: A rapid ecological assessment. Phase 1: An island-wide characterization of mapping of natural communities and modified vegetation types. Washington, DC: The Nature Conservancy.
- Henry, A. G. 2011. Plant foods and the dietary ecology of Neanderthals. American Journal of Physical Anthropology 144: 160–1.
- Hilbert, L., E. G. Neves, F. Pugliese, B. S. Whitney, M. Shock, E. Veasey, C. A. Zimpel, and J. Iriarte. 2017. Evidence for mid-Holocene rice domestication in the Americas. Nature Ecology & Evolution 1 (11):1693–8. doi:10.1038/s41559-017-0322-4
- Hofman, C. L., and M. L. Hoogland. 2015. Investigaciones arqueológicas en los sitios El Flaco (Loma de Guayacanes) y La Luperona (UNIJICA) [Archaeological investigations at the El Flaco (Loma de Guayacanes) and La Luperona (UNIJICA) sites]. Informe Preliminary. Boletín del Museo del Hombre Dominicano 46 (42):61–7.
- Hofman, C. L, and M. L. P. Hoogland. 2004. Social dynamics and change in the northern Lesser Antilles. In Late Ceramic Age societies in the eastern Caribbean, ed. A. Delpuech and C. L. Hofman, 1–22. Oxford: BAR International Series 1273. British Archaeological Reports.
- Hofman, C. L., J. U. Hung, E. H. Malatesta, J. S. Jean, T. Sonnemann, and M. L. Hoogland. 2018. Indigenous Caribbean perspectives: Archaeologies and legacies of the first colonised region in the New World. Antiquity 92 (361):200–16. doi:10.15184/aqy.2017.247
- Hooghiemstra, H., T. Olijhoek, M. Hoogland, M. Prins, B. van Geel, T. Donders, W. Gosling, and C. Hofman. 2018. Columbus' environmental impact in the New World: Land use change in the Yaque River valley, Dominican Republic. The Holocene 28 (11):1818–35. doi:10.1177/0959683618788732
- Hoogland, M. L., and C. L. Hofman. 1999. Expansion of the Taino cacicazgos towards the Lesser Antilles. Journal de la Société des Américanistes 85 (1):93–113. doi:10.3406/jsa.1999.1731
- Horst, H. A., and A. Garner. 2007. Jamaican Americans. New York: Chelsea House Publications.
- Howard, R. R. 1956. The archaeology of Jamaica: A preliminary survey. American Antiquity 22 (1):45–54. doi:10.2307/276166
- Howard, R. R. 1965. New perspectives in Jamaican archaeology. American Antiquity 31 (2Part1):250–5. doi:10.2307/2693993
- Iriarte, J., S. Elliott, S. Y. Maezumi, D. Alves, R. Gonda, M. Robinson, J. G. de Souza, J. Watling, and J. Handley. 2020. The origins of Amazonian landscapes: Plant cultivation, domestication and the spread of food production in tropical South America. Quaternary Science Reviews 248:106582. doi:10.1016/j.quascirev.2020.106582
- Jarzombek, M. M. 2013. Architecture of first societies: A global perspective. New Jersey: John Wiley & Sons.
- Juggins, S. 2007. C2: Software for ecological and palaeoecological data analysis and visualisation (user guide version 1.5). Newcastle upon Tyne: Newcastle University 77.
- Keegan, W. F. 1994. West Indian archaeology. 1. Overview and foragers. Journal of Archaeological Research 2 (3):255–84. doi:10.1007/BF02231434
- Keegan, W. F. 2000. West Indian archaeology. 3. ceramic age. Journal of Archaeological Research 8 (2):135–67. doi:10.1023/A:1009403127753
- Keegan, W. F. 2006. Archaic influences in the origins and development of Taino societies. Caribbean Journal of Science 42 (1):1.
- Keegan, W. F., and C. Hofman. 2017. The Caribbean before Columbus. New York: Oxford University Press.
- Kennedy, L. M., S. P. Horn, and K. H. Orvis. 2006. A 4000-year record of fire and forest history from Valle de Bao, Cordillera Central, Dominican Republic. Palaeogeography, Palaeoclimatology, Palaeoecology 231 (3–4):279–90. doi:10.1016/j.palaeo.2005.08.002
- Lane, C. S., S. P. Horn, C. I. Mora, and K. H. Orvis. 2009. Late-Holocene paleoenvironmental change at mid-elevation on the Caribbean slope of the Cordillera Central, Dominican Republic: A multi-site, multi-proxy analysis. Quaternary Science Reviews 28 (23–24):2239–60. doi:10.1016/j.quascirev.2009.04.013
- LeFebvre, M. J., and S. D. DeFrance. 2014. Guinea pigs in the pre-Columbian West Indies. The Journal of Island and Coastal Archaeology 9 (1):16–44. doi:10.1080/15564894.2013.861545
- LeFebvre, M. J., C. M. Giovas, and J. E. Laffoon. 2019. Advancing the study of Amerindian ecodynamics in the Caribbean: Current perspectives. Environmental Archaeology 24 (2):107–14.
- Li, Z., and B. Delvaux. 2019. Phytolith‐rich biochar: A potential Si fertilizer in desilicated soils. GCB Bioenergy 11 (11):1264–82. doi:10.1111/gcbb.12635
- Mickleburgh, H. L., J. E. Laffoon, J. R. Pagán-Jiménez, A. A. Mol, S. Walters, Z. J. Beier, and C. L. Hofman. 2019. Precolonial/early colonial human burials from the site of White Marl, Jamaica: New findings from recent rescue excavations. International Journal of Osteoarchaeology 29 (1):155–61. doi:10.1002/oa.2707
- Mickleburgh, H. L., and J. R. Pagán-Jiménez. 2012. New insights into the consumption of maize and other food plants in the pre-Columbian Caribbean from starch grains trapped in human dental calculus. Journal of Archaeological Science 39 (7):2468–78. doi:10.1016/j.jas.2012.02.020
- Mol, A. A. 2014. The connected Caribbean: A socio-material network approach to patterns of homogeneity and diversity in the pre-colonial period. Leiden: Sidestone Press.
- Morcote-Ríos, G., R. Bernal, and L. Raz. 2016. Phytoliths as a tool for archaeobotanical, palaeobotanical and palaeoecological studies in Amazonian palms. Botanical Journal of the Linnean Society 182 (2):348–60. doi:10.1111/boj.12438
- Napolitano, M. F., R. J. DiNapoli, J. H. Stone, M. J. Levin, N. P. Jew, B. G. Lane, J. T. O’Connor, and S. M. Fitzpatrick. 2019. Reevaluating human colonization of the Caribbean using chronometric hygiene and Bayesian modeling. Science Advances 5 (12):eaar7806. doi:10.1126/sciadv.aar7806
- Newsom, L. A. 1988. Paleoethnobotanical remains from Windover Site, Florida: Human abdominal samples. Journal of Wetland Archaeology 4:5–7.
- Newsom, L. A. 1993. Native West Indian plant use. PhD diss., University of Florida.
- Newsom, L. A., and E. S. Wing. 2004. On land and sea: Native American uses of biological resources in the West Indies. Tuscaloosa: University of Alabama Press.
- Ollendorf, A. L., S. C. Mulholland, and G. Rapp, Jr. 1988. Phytolith analysis as a means of plant identification: Arundo donax and Phragmites communis. Annals of Botany 61 (2):209–14. doi:10.1093/oxfordjournals.aob.a087544
- Pagán-Jiménez, J. R. 2005. En diálogo con José R. Oliver y Reniel Rodríguez Ramos. La emergencia de la temprana producción de vegetales en nuestros esquemas investigativos y algunos fundamentos metodológicos del estudio de almidones [In dialogue with José R. Oliver and Reniel Rodríguez Ramos. The emergence of early vegetable production in our research schemes and some methodological foundations of the study of starches]. Diálogo Antropológico 3 (10):49–55.
- Pagán-Jiménez, J. R. 2007. De antiguos pueblos y culturas botánicas en el Puerto Rico indígena: el archipiélago borincano y la llegada de los primeros pobladores agroceramistas [Of ancient towns and botanical cultures in the Port Rich indigenous: The Borincano archipelago and the arrival of the first agro potter villagers]. 18. Oxford: British Archaeological Reports.
- Pagán-Jiménez, J. R. 2011. Early phytocultural processes in the pre-Colonial Antilles. A Pan-Caribbean survey for an ongoing starch grain research. In Communities in contact: Essays in archaeology, ethnohistory, ethnography of the Amerindian circum-Caribbean, ed. C. L. Hofman and A. Van Duijvenbode, 87–116. Leiden: Sidestone Press.
- Pagán-Jiménez, J. R. 2013. Human–plant dynamics in the precolonial Antilles. In The Oxford handbook of Caribbean archaeology, ed. W. F. Keegan, C. L. Hofman, and R. R. Ramos, 391–406. New York: Oxford University Press.
- Pagán-Jiménez, J. R., Z. Ali, C. G. Santiago-Marrero, and C. L. Hofman. 2020. Plantscapes of dwelling: Precolonial household mounds, phytocultural dynamics and the ensuing human ecosystems at El Flaco and El Carril (cal. AD 990–1450), northern Dominican Republic. Review of Palaeobotany and Palynology 274:104160. doi:10.1016/j.revpalbo.2020.104160
- Pagán-Jiménez, J. R., and H. L. Mickleburgh. 2022. Caribbean deep-time culinary worlds revealed by ancient food starches: Beyond the dominant narratives. Journal of Archaeological Research 1–47. doi:10.1007/s10814-021-09171-3
- Pagán-Jiménez, J. R., R. Rodríguez-Ramos, and C. Hofman. 2019. On the way to the islands: The role of domestic plants in the initial peopling of the Antilles. In Early settlers of the insular Caribbean. Dearchaizing the Archaic, ed. C. L. Hofman and A. Antczak, 89–106. Leiden: Sidestone Press.
- Pagán-Jiménez, J. R., R. Rodríguez-Ramos, B. A. Reid, M. van den Bel, and C. L. Hofman. 2015. Early dispersals of maize and other food plants into the Southern Caribbean and Northeastern South America. Quaternary Science Reviews 123:231–46. doi:10.1016/j.quascirev.2015.07.005
- Parry, J. H. 1955. Plantation and provision ground: An historical sketch of the introduction of food crops into Jamaica. Revista de Historia de América 35:1–20.
- Pearsall, D. M. 1985. Analysis of soil phytoliths and botanical macroremains from El Bronce archaeological site, Ponce, Puerto Rico. Final Report, Phase 2. Puerto Rico: Archaeological data recovery at El Bronce.
- Pestle, W. J. 2013. Fishing down a prehistoric Caribbean marine food web: Isotopic evidence from Punta Candelero, Puerto Rico. The Journal of Island and Coastal Archaeology 8 (2):228–54. doi:10.1080/15564894.2013.797943
- Petersen, J. B., C. L. Hofman, and L. A. Curet. 2004. Time and culture: Chronology and taxonomy in the eastern Caribbean and the Guianas. In Late Ceramic Age Societies in the Eastern Caribbean Ed. A. Delpuech and C. L. Hofman, 17–33. Bar International Series 1273. Oxford: British Archaeological Reports.
- Piperno, D. R. 2002. Phytolithic remains from the Tutu site in Tutu Archaeological Village Site: A multidisciplinary case study in human adaptation, ed. E. Righter, 135–40. London: Routeledge.
- Piperno, D. R. 2006. Phytoliths: A comprehensive guide for archaeologists and paleoecologists. New York: AltaMira Press.
- Piperno, D. R., and C. McMichael. 2020. Phytoliths in modern plants from Amazonia and the Neotropics at large: Implications for vegetation history reconstruction. Quaternary International 565:54–74. doi:10.1016/j.quaint.2020.10.043
- Pope, K. O., M. E. Pohl, J. G. Jones, D. L. Lentz, C. V. Nagy, F. J. Vega, and I. R. Quitmyer. 2001. Origin and environmental setting of ancient agriculture in the lowlands of Mesoamerica. Science 292 (5520):1370–3. doi:10.1126/science.292.5520.1370
- Pujol, B., G. Gigot, G. Laurent, M. Pinheiro-Kluppel, M. Elias, M. Hossaert-McKey, and D. McKey. 2002. Germination ecology of cassava (Manihot esculenta Crantz, Euphorbiaceae) in traditional agroecosystems: Seed and seedling biology of a vegetatively propagated domesticated plant. Economic Botany 56 (4):366–79. doi:10.1663/0013-0001(2002)056[0366:GEOCME2.0.CO;2]
- Rampersad, S. R. 2009. Targeting the Jamaican Ostionoid: The Blue Marlin Archaeological Project. Caribbean Quarterly 55 (2):23–42. doi:10.1080/00086495.2009.11829757
- Reimer, P. J., W. E. N. Austin, E. Bard, A. Bayliss, P. G. Blackwell, C. Bronk Ramsey, M. Butzin, H. Cheng, R. L. Edwards, M. Friedrich, et al. 2020. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62 (4):725–57. doi:10.1017/RDC.2020.41
- Richardson, B. C. 2004. Igniting the Caribbean's past: Fire in British West Indian history. Chapel Hill: University of North Carolina Press.
- Rival, D. L., and D. McKey. 2008. Domestication and diversity in Manioc (Manihot esculenta Crantz ssp. Esculenta, Euphorbiaceae) Current Anthropology 49 (6):1119–28. doi:10.1086/593119
- Robinson, M., C. Jaimes‐Betancourt, S. Elliott, S. Y. Maezumi, L. Hilbert, D. Alves, J. G. Souza, and J. Iriarte. 2021. Anthropogenic soil and settlement organisation in the Bolivian Amazon. Geoarchaeology 36 (3):388–403. doi:10.1002/gea.21839
- Rodríguez Suárez, R, and J. R. Pagán-Jiménez. 2008. The Burén in precolonial Cuban archaeology: New information regarding the use of plants and ceramic griddles during the Late Ceramic Age of Eastern Cuba gathered through starch analysis. In Crossing the borders, new methods techniques in the study of archaeological materials from the Caribbean, ed. C. L. Hofman, M. L. P. Hoogland and A. Van Gijn, 159–69. Tuscaloosa: The University of Alabama Press.
- Rosen, A. M. 1999. Phytolith analysis in Near Eastern archaeology. In The practical impact of science on Aegean and Near Eastern Archaeology, ed. S. Pike and S. Gitin, 86–92. London: Archetype Press.
- Rouse, I. 1986. Migrations in prehistory: Inferring population movement from cultural remains. New Haven: Yale University Press.
- Rouse, I. 1992. The Tainos: Rise and decline of the people who greeted Columbus. New Haven: Yale University Press.
- Salzman, V. T., and W. S. Judd. 1995. A revision of the Greater Antillean species of Bactris (Bactridinae: Arecaceae). Brittonia 47 (4):345–71. doi:10.2307/2807563
- Sara, T. R., and J. J. O. Aguilú. 2003. Paleoenvironmental Investigations of Navy Lands on Vieques Island, Puerto Rico. Rapport non publié préparé pour Geo-Marine, Inc. au Department of The Navy, Atlantic Division, Naval Facilities Engineering Command. Number 280.
- Shev, G. T., J. E. Laffoon, and C. L. Hofman. 2021. Human and hutia (Isolobodon portoricensis) interactions in pre-Columbian Hispaniola: The isotopic and morphological evidence. Journal of Archaeological Science: Reports 37:102913. doi:10.1016/j.jasrep.2021.102913
- Shillito, L. M. 2018. Phytolith analysis. In The encyclopedia of archaeological sciences, ed. S. López Varela and J. Thomas, 1–3. Hoboken: John Wiley & Sons Ltd.
- Siegel, P. E. 1992. Ideology, power, and social complexity in prehistoric Puerto Rico. New York: State University of New York at Binghamton.
- Siegel, P. E., J. G. Jones, D. M. Pearsall, N. P. Dunning, P. Farrell, N. A. Duncan, J. H. Curtis, and S. K. Singh. 2015. Paleoenvironmental evidence for first human colonization of the eastern Caribbean. Quaternary Science Reviews 129:275–95. doi:10.1016/j.quascirev.2015.10.014
- Siegel, P. E., J. G. Jones, D. M. Pearsall, N. P. Dunning, P. Farrell, N. A. Duncan, J. H. Curtis, and S. K. Singh. 2018. Humanizing the landscapes of the Lesser Antilles during the Archaic Age. In The archaeology of Caribbean and Circum-Caribbean Farmers (6000 bc–ad 1500), ed. B. A. Reid, 55–70. New York: Routledge.
- Silverberg, J., R. Vanderwal, and E. Wing. 1972. The White Marl site in Jamaica: Report of the 1964 Robert R Howard excavations. Milwaukee: Department of Anthropology, University of Wisconsin.
- Sonnemann, T. F., J. Ulloa Hung, and C. L. Hofman. 2016. Mapping indigenous settlement topography in the Caribbean using drones. Remote Sensing 8 (10):791. doi:10.3390/rs8100791
- Sponsel, L. E. 1998. The historical ecology of Thailand: Increasing thresholds of human environmental impact from prehistory to the present. In Advances in historical ecology, ed. W. Balee, 376. New York: Columbia University Press.
- Steadman, D. W., H. M. Singleton, K. M. Delancy, N. A. Albury, J. A. Soto-Centeno, H. Gough, N. Duncan, J. Franklin, and W. F. Keegan. 2017. Late Holocene historical ecology: The timing of vertebrate extirpation on Crooked Island, Commonwealth of the Bahamas. The Journal of Island and Coastal Archaeology 12 (4):572–84. doi:10.1080/15564894.2017.1305469
- Stokes, B. J. 2002. Prehistoric occupation of Rio Nuevo Bay. Jamaican Historical Society Bulletin 11 (10):295–7.
- Twiss, P. C., E. Suess, and R. M. Smith. 1969. Morphological classification of grass Phytoliths 1. Soil Science Society of America Journal 33 (1):109–15. doi:10.2136/sssaj1969.03615995003300010030x
- Vanderwal, R. L. 1968. The prehistory of Jamaica: A ceramic study. Unpublished master's thes., University of Wisconsin-Milwaukee, Milwaukee, WI.
- Watling, J., and J. Iriarte. 2013. Phytoliths from the coastal savannas of French Guiana. Quaternary International 287:162–80. doi:10.1016/j.quaint.2012.10.030
- Watling, J., J. Iriarte, B. Whitney, E. Consuelo, F. Mayle, W. Castro, D. Schaan, and T. R. Feldpausch. 2016. Differentiation of neotropical ecosystems by modern soil phytolith assemblages and its implications for palaeoenvironmental and archaeological reconstructions II: Southwestern Amazonian forests. Review of Palaeobotany and Palynology 226:30–43. doi:10.1016/j.revpalbo.2015.12.002
- Wesler, K. W. 2013. Jamaica. In The Oxford handbook of Caribbean archaeology, ed. W. F. keegan, C. L. Hofman, and R. R Ramos, 250-63. New York: Oxford University Press.
- Williams, M. A. 2019. A multi-proxy paleoecological reconstruction of Holocene climate, vegetation, fire and human activity in Jamaica. West Indies: Master of Science, Ecology and Environmental Science, The University of Maine.
- Wilson, S. M. 2007. The archaeology of the Caribbean. Cambridge: Cambridge University Press.
- Wing, E. S. 1961. Animal remains excavated at the Spanish site of Nueva Cadiz on Cubagua Island, Venezuela. New West Indian Guide/Nieuwe West-Indische Gids 41 (1):162–5. doi:10.1163/22134360-90002346
- Wing, E. S. 1972. Identification and interpretation of faunal remains from the white marl site in Jamaica. In The White Marl site in Jamaica: Report of the 1964 Robert R. Howard Excavation, ed. J. R. L. Silverberg and R. L. Vanderwal, 18–35. Milwaukee, WI: University of Wisconsin-Milwaukee.