
ABSTRACT
Seed dormancy and germination are the most important adaptive traits of seed plants, which control the germination in a proper space and time. Internal genetic factors together with environmental cues govern seed dormancy and germination. Abscisic acid (ABA), a key phytohormone induces seed dormancy and inhibits seed germination through its molecular genetic signaling network responding the seed inherent physiological and environmental factors. Recently, auxin has been shown to be another phytohormone that induces seed dormancy. We have recently shown that germonstatin (GS), a small synthetic molecule identified by high through-put chemical genetic screenings, inhibits seed germination through up-regulating auxin signaling and inducing auxin biosynthesis. GERMOSTATIN RESISTANCE LOCUS 1 (GSR1) encodes a plant homeodomain (PHD) finger protein and is responsible for GS seed germination inhibition. Its knockdown mutant gsr1 displays decreased dormancy. In this report, we show that GS is not an ABA analog and provided 2 other GS-resistant mutants related to the chemical's function in seed germination inhibition other than gsr1, suggesting that GS may have pleiotropic effects through targeting different pathway governing seed germination.
It has long been known that the levels of phytohormones, gibberellins (GAs) and ABA, determine dormancy and germination in plants.Citation1,2 GAs, the essential germination activators, break seed dormancy, while ABA induces seed dormancy. The ABA/GA ratio regulates the transition from dormancy to germination.Citation3 Recently, auxin has been identified as another dormancy regulatory phytohormone, which induces and maintains seed dormancy.Citation4 Auxin maintains seed dormancy through AUXIN RESPONSE FACTOR (ARF) 10/16-mediated expression of ABI3, a key regulator in the ABA-mediated seed dormancy pathway.Citation4 What's more, auxin also acts as a positive regulator of gemma dormancy in liverwort.Citation5 GS, the sythetic small molecule reported in our recent study, inhibits seed germination through up-regulating auxin response and enhancing auxin biosynthesis.Citation6 GSR1 encodes a PHD finger protein with H3K4 binding activity; and its knockdown mutant gsr1 shows resistance to GS; GSR1 involves in seed germination by interacting with IAA 17 and ARF10/16. We here reported the function difference between GS and ABA and the pleiotropic biological effects of GS.Citation6
Due to the fact that GS and ABA both inhibit seed germination and several ABA function analogs have been reported,Citation7-10 we therefore examined whether GS is also an ABA function analog. Our results showed the seeds would germinate when they were moved out of the growing plate with 10 µM ABA for 36 hours, indicating that ABA's effect on seed germination could be erased possibly through its biochemistry modification in seeds (). ABA, the stressed induced phytohormone, can be deactivated by ABA-8’-hydroxylase when plants were transferred to more suitable growth conditions.Citation11 Unlike ABA, the seed inhibitory effect of GS remained when the seeds were move out from their GS-containing growing MS plates (), indicating that GS possibly would not be biochemically modified in plant or the effects of GS on its target/targets could be perment and irreversible. However, hypostatin, another synthesis small molecule, could be glycoactivated by a UDP glycosyltransferase and affected plant growth irreversible upon the chemical treatment for a certain long period.Citation12 ABI3 and ABA INSENSITIVE 4 (ABI4) are 2 important downstream transcription factors of ABA core signaling and their loss of function mutants are hyposensitive to ABA treatment.Citation13 However, GS inhibits seed germination of abi3-8 and abi4-1, the 2 ABA sensitive mutants, suggesting that GS may not function through ABA signaling. Taken together, GS, like ABA, displays seed inhibitory activity but may function in pathway independent from ABA.
Figure 1. The pleiotropic effects of germostatin on Arabidopsis seed germination. (A) The different effects of germostatin (GS) and ABA on seed germiantion. Seeds of Col-0 were sowed on 1/2 MS medium with 10 µM ABA (left) and 15µM GS (right) for 36 hours in dark. After 36-hour incubation the seeds were moved to 1/2 MS medium for 3-day incubation in dark. (B) ABA insensitive mutants are not resistant to germostatin (GS). Wild-type (Col-0), abi3-8 and abi4-1 were grown on 1/2 MS with 13 µM germostatin (GS) for 36 hours in dark. (C) Multiple GS resistant mutants were acquired from mutant screenings. gsr1, gsr2, and gsr3 were GS resistant mutants. The seeds of Col-0 (WT), gsr1, gsr2, and gsr3 were sowed on 13 µM germostatin (GS) and incubated for 3 d in dark.
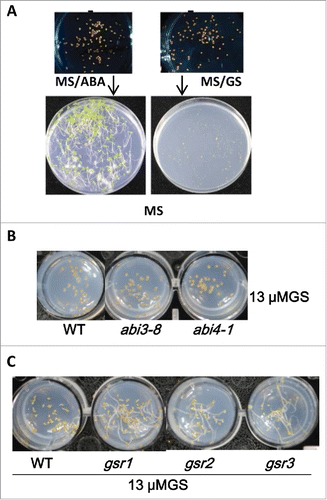
Exotic active small molecule may act as an agonist or antagonist of certain active indigenous molecule such as hormone, substrate analog, as well as regulatory ligand to perturb the function of a protein.Citation14 The central questions of a chemical genetics research are what the chemical's target is and how the chemical perturbs certain biological process through its acting on a certain protein. The targets of pyrabactin, a synthetic small molecule, have been identified by the chemical resistant mutant screenings.Citation8 We have identified GS resistant mutant gsr1 and GSR1 encodes a PHD finger protein with function involved in auxin signaling.Citation6 GS inhibits seed germination possibly through auxin signaling. However, auxin inhibits seed germination at a higher concentration when compared with GS. Reasonably, such higher activity of GS on seed germination inhibition could be due to its pleiotropic effects on seed germination. Besides gsr1, we have identified 2 other GS resistant mutants, gsr2 and gsr3, with different mutation locus, which possibly act simultaneously or synergistically with gsr1 to respond GS to inhibit seed germination (). At present, we have cloned GSR2, which encodes a RNA polymerase (the results will be published elsewhere). Characterizing the action modes of GS on seed germination will require firstly identification these responsible genetic loci, which may provide plant science community a research bonanza to explore the molecular genetic mechanism of seed germination in future.
These additional data shed light on the role of GS in the study of seed germination. Further research is needed to better understand the interaction linking GS to its other resistant mutants to reveal the GS target(s).
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgment
We thank Ziying Gong (Shanghai Institute of Plant Physiology and Ecology) for the preliminary work to this study.
Funding
This work was funded by grants from the National Natural Science Foundation of China (31171293 and 31371361) and the One Hundred Talents Program of the Chinese Academy of Sciences.
References
- Seo M, Hanada A, Kuwahara A, Endo A, Okamoto M, Yamauchi Y, North H, Marion-Poll A, Sun TP, Koshiba T, et al. Regulation of hormone metabolism in Arabidopsis seeds: phytochrome regulation of abscisic acid metabolism and abscisic acid regulation of gibberellin metabolism. Plant J 2006; 48:354–66; PMID:17010113; http://dx.doi.org/ 10.1111/j.1365-313X.2006.02881.x
- Finkelstein R, Reeves W, Ariizumi T, Steber C. Molecular aspects of seed dormancy. Annu Rev Plant Biol 2008; 59:387–415; PMID:18257711; http://dx.doi.org/10.1146/annurev.arplant.59.032607.092740
- Bentsink L, and Koornneef M. Seed dormancy and germination. The Arabidopsis Book 2008; 6:e0119; PMID:22303244; http://dx.doi.org/10.1199/tab.0119
- Liu X, Zhang H, Zhao Y, Feng Z, Li Q, Yang HQ, Luan S, Li J, and He ZH. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc Natl Acad Sci USA 2013; 110:15485–90; PMID:23986496; http://dx.doi.org/10.1073/pnas.1304651110
- Eklund DM, Ishizaki K, Flores-Sandoval E, Kikuchi S, Takebayashi Y, Tsukamoto S, Hirakawa Y, Nonomura M, Kato H, Kouno M, et al. Auxin produced by the indole-3-pyruvic acid pathway regulates development and gemmae dormancy in the liverwort Marchantia polymorpha. Plant cell 2015; 27:1650–69; PMID:26036256; http://dx.doi.org/10.1105/tpc.15.00065
- Ye Y, Gong Z, Lu X, Miao D, Shi J, Lu J, Zhao Y. GermoStatin Resistance locus 1 encodes a PHD domain protein involved in auxin-mediated seed dormancy and germination. Plant J 2015; 85:3-15; PMID:26611158; http://dx.doi.org/10.1111/tpj.13086.
- Park SY, Peterson FC, Mosquna A, Yao J, Volkman BF, Cutler SR. Agrochemical control of plant water use using engineered abscisic acid receptors. Nature 2015; 520:545–8; PMID:25652827; http://dx.doi.org/10.1038/nature14123
- Park SY, Fung P, Nishimura N, Jensen DR, Fujii H, Zhao Y, Lumba S, Santiago J, Rodrigues A, Chow TF, et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009; 324:1068–71; PMID:19407142; http://dx.doi.org/10.1126/science.1173041
- Cao M, Liu X, Zhang Y, Xue X, Zhou XE, Melcher K, Gao P, Wang F, Zeng L, Zhao Y, et al. An ABA-mimicking ligand that reduces water loss and promotes drought resistance in plants. Cell Res 2013; 23:1043–54; PMID:23835477; http://dx.doi.org/10.1038/cr.2013.95
- Okamoto M, Peterson FC, Defries A, Park SY, Endo A, Nambara E, Volkman BF, Cutler SR. Activation of dimeric ABA receptors elicits guard cell closure, ABA-regulated gene expression, and drought tolerance. Proc Natl Acad Sci USA 2013; 110:12132–7; PMID:23818638 http://dx.doi.org/10.1073/pnas.1305919110
- Yang SH, Zeevaart JA. Expression of ABA 8'-hydroxylases in relation to leaf water relations and seed development in bean. Plant J 2006; 47:675–86; PMID:16856981; http://dx.doi.org/10.1111/j.1365-313X.2006.02815.x
- Zhao Y, Chow TF, Puckrin RS, Alfred SE, Korir AK, Larive CK, Cutler SR. Chemical genetic interrogation of natural variation uncovers a molecule that is glycoactivated. Nat Chem Biol 2007; 3:716–21; PMID:17891152; http://dx.doi.org/10.1038/nchembio.2007.32
- Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR. Abscisic acid: emergence of a core signaling network. Annu Rev Plant Biol 2010; 61:651–79; PMID:20192755; http://dx.doi.org/10.1146/annurev-arplant-042809-112122
- Toth R, van der Hoorn, RA. Emerging principles in plant chemical genetics. Trends Plant Sci 2010; 15:81–8; PMID:20036182; http://dx.doi.org/10.1016/j.tplants.2009.11.005