
ABSTRACT
As an apoplast signal molecule, extracellular ATP (eATP) is involved in the growth regulation of Arabidopsis thaliana seedlings. Recently, RRFT1 was revealed to be involved in eATP- regulated seedling growth. To further verify the role of RRTF1 in seedlings' eATP response, expression of 20 eATP-responsive genes in wild type (Col-0) and RRTF1 null mutant (rrtf1-1) seedlings were investigated by using realtime quantitative PCR. After 0.5 mM ATP stimulation, the response of these genes' expression in rrtf1-1 seedlings was significantly different from that in Col-0 seedlings. Proteins which are encoded by these genes include transcription factors, plasma membrane receptors like kinases, ion influx/efflux transporters and hormone signaling components. The results indicated that RRTF1 may be involved in eATP regulated physiological responses via regulating the expression of some functional genes.
As an apoplast signal molecule, extracellular ATP (eATP) is involved in the growth regulation of Arabidopsis thaliana seedlings. Recently, RRFT1 was revealed to be involved in eATP- regulated seedling growth. To further verify the role of RRTF1 in seedlings’ eATP response, expression of 20 eATP-responsive genes in wild type (Col-0) and RRTF1 null mutant (rrtf1-1) seedlings were investigated by using realtime quantitative PCR. After 0.5 mM ATP stimulation, the response of these genes’ expression in rrtf1-1 seedlings was significantly different from that in Col-0 seedlings. Proteins which are encoded by these genes include transcription factors, plasma membrane receptor-like kinases, ion influx/efflux transporters, and hormone signaling components. The results indicated that RRTF1 may be involved in eATP regulated physiological responses via regulating the expression of some functional genes.
Extracellulr ATP, which is secreted from the cytoplasm, acts as a signaling molecule in the extracellular matrix. eATP is involved in regulating vegetative growth, reproductive processes, and stress responses.Citation1–Citation4 Functional gene expression and protein synthesis may be a central event in these eATP-stimulated responses.Citation5,Citation6 Hence, transcription factors may play a key role in eATP-regulated physiological processes as well. Some ethylene-responsive factors were reported to be induced by eATP and involved in eATP signaling.Citation7–Citation9 Recently, RRTF1, an ethylene-responsive transcription factor, was revealed to participate in eATP-regulated root & hypocotyl growth via regulating auxin transport and functional genes’ expression.Citation10 In 2017, in an DNA microarray assay, we found some ATP responsive genes which may possibly be involved in eATP-regulated root growth.Citation9 To further clarify the role of RRTF1 in eATP signaling, expression of these genes in eATP-treated wild type (Col-0) and RRTF1 null mutant (rrtf1-1) seedlings were detected by using realtime quantitative PCR.
Arabidopsis thaliana seeds were surface-sterilized and sown on the surface of solid 1/2 MS medium (containing 0.8% phytagel) in square culture dishes. After 2 days vernalization at 4°C, culture dishes were vertically placed in a growth chamber which was running at 22°C, 130 μmol/m2·s illumination and 16/8 h light/dark ratio. 4-d-old seedlings were then transplanted onto 0.5 mM ATP-containing medium which was made according to Zhu et al.Citation10 1/2 MS medium was pre-adjusted to maintain the final pH value around 5.8. 2 or 3 h after transplantation, seedlings were harvested and rapidly frozen in liquid nitrogen for RNA extraction. Total RNA was isolated from 0.1 g root tissue using the RNAiso plus reagent, and 2 μg of total RNA was reverse-transcribed to cDNA using M-MLV reverse transcriptase. Realtime quantitative PCR was performed using an ABI7500 system, ACTIN2 was used as the constitutive reference gene. The primers’ sequences are listed in Table S1.
The results showed that, after ATP stimulation, 20 genes’ expression pattern were markedly different in Col-0 and rrtf1-1 seedlings. The responsiveness of these genes’ expression can be sorted into three categories: (1) 14 genes’ expression were remarkably up-regulated in Col-0 while not up-regulated or even down-regulated in rrtf1-1, e.g. ABCG39, ABCG40, XTH33, GH3.4, etc.; (2) 3 genes’ expression were not or only slightly up-regulated in Col-0 while remarkably down-regulated in rrtf1-1, e.g. LECRK13, XTH22, and CRK5; (3) 3 genes’ expression were remarkably down-regulated or not changed in Col-0 while markedly up-regulated in rrtf1-1, e.g., ACA13, SAUR68, and At3g28580 (). The results indicated that RRTF1 may be involved in eATP regulated gene expression as a positive or negative regulator.
Figure 1. The expression of 20 eATP responsive genes in Col-0 and rrtf1-1 seedlings. Expression of gene was detected by using realtime quantitative PCR, which Actin2 was used as a reference gene. The primers of these genes are listed in Table 2. Four-d seedlings were transplanted onto 0.5 mM ATP-containing medium and cultured under light for 2 or 3 h. Then, total RNA was extracted and expression of each gene was detected. Data from three biological replicates (each include three technical replicates) were used to calculate the mean ± SD and statistically analyzed. Student’s t-test p-values: * p < .05, ** p < .01.
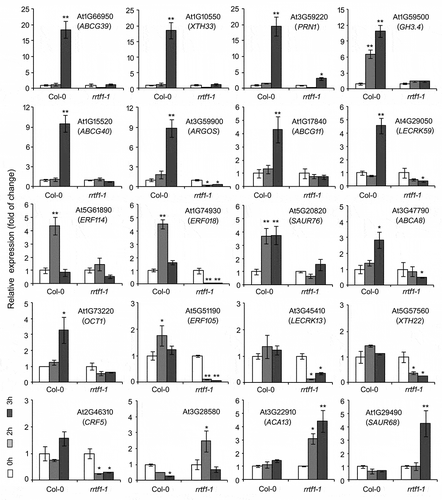
Based on the data from the NCBI (National Center for Biotechnology Information) and UniProt (Universal Protein) website, proteins which are encoded by these genes can be categorized into five groups: (1) Transcription factors, 3 ethylene-responsive transcription factors (ERF018, ERF105, and ERF114) are included; (2) Cell wall construction involved enzymes, XTH22, and XTH33 are included; (3) Plasma membrane lectin-domain containing receptor kinases, LECRK13, and LECRK59 are included; (4) Plasma membrane transporters, 4 ABC (ATP Binding Cassette) transporters (ABCA8, ABCG11, ABCG39, and ABCG40), an organic cation transporter (OCT1) and a Ca2+ ATPase (ACA13) are included; (5) Hormone metabolism and signaling related proteins, an IAA-amido synthase (GH3.4), 3 auxin-responsive proteins (SAUR68, SAUR76, and ARGOS), 2 ABA signaling related proteins (PRN1 and At3G28580) and 1 cytokinin responsive factor (CRK5) are included ().
Table 1. Function analysis of RRTF1-regulated eATP-responsive genes. Description of the genes’ function is based on information from website of NCBI (The National Center for Biotechnology Information) and UniProt (Universal Protein).
These proteins are involved in multiple physiological processes, including cell division & differentiation, and stress (or defense) responses. eATP-regulated expression of these genes may be involved in eATP-regulated seedling growth; hence, the different responses of Col-0 and rrtf1-1 seedlings to eATP stimulationCitation10 may partially result from the differential expression pattern of these genes in seedlings. Recently, by using chromatin immunoprecipitation (CHIP) and high throughput sequencing analysis, 48 genes were screened out to be ATP responsive genes which may be directly regulated by RRTF1.Citation10 The results here provided additional evidence that RRTF1 may be involved in eATP signaling. Nevertheless, the 20 genes we detected here were not enriched in CHIP assay, so RRTF1 may possibly regulate the expression of these genes indirectly.
During the past 2 decades, it was revealed that seedlings responded to the high concentration of eATP as a suppressed growth rate and changed the growth direction of roots or hypocotyls.Citation7–Citation12 Plant hormone (auxin, jasmonic acid, ethylene) signaling components were revealed to be involved in eATP regulated seedling growth.Citation6–Citation14 The 20 proteins had been proved or predicted to be involved in stimuli-regulated vegetative organs growth. Results here provided further evidence that these proteins may play roles in eATP signaling as well. RRTF1 has been revealed to be involved in plant growth and stress responses.Citation15,Citation16 Results here provided new evidence to elucidate the role of RRTF1 in Arabidopsis seedlings’ response to eATP. It is of good help for understanding the role of ethylene-responsive transcription factors in the signaling of apoplast messengers. Recently, it was revealed that MYC and CAMTA3 transcription factors play essential roles in regulating eATP responsive genes’ expression.Citation6 It is expected that the discovery of more transcription factors in eATP signaling will provide valuable information to elucidate the mechanism of eATP- regulated gene expression.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Supplemental Material
Download MS Word (15.7 KB)Supplementary material
Supplemental data for this article can be accessed on the publisher’s website.
Additional information
Funding
References
- Tanaka K, Gilroy S, Jones AM, Stacey G. Extracellular ATP signaling in plants. Trends Cell Bio. 2010;20:1–4. PMID:20817461. doi:10.1016/j.tcb.2010.07.005.
- Clark G, Roux S. Apyrases, extracellular ATP and the regulation of growth. Curr Opin Plant Biol. 2011;14:700–706. PMID:21855397. doi:10.1016/j.pbi.2011.07.013.
- Cao Y, Tanaka K, Nguyen C, Stacey G. Extracellular ATP is a central signaling molecule in plant stress responses. Curr Opin Plant Biol. 2014;20:82–87. PMID:24865948. doi:10.1016/j.pbi.2014.04.009.
- Clark G, Morgan R, Fernandez M, Salmi M, Roux S. Breakthroughs spotlighting roles for extracellular nucleotides and apyrases in stress responses and growth and development. Plant Sci. 2014;225:107–116. PMID:25017166. doi:10.1016/j.plantsci.2014.06.002.
- Chivasa S, Tome D, Hamilton J, Slabas A. Proteomic analysis of extracellular ATP-regulated proteins identifies ATP synthase β-subunit as a novel plant cell death regulator. Mol Cell Proteomics. 2011;10:M110.003905. PMID:21156838. doi:10.1074/mcp.M110.003905.
- Jewell J, Sowders J, He R, Willis M, Gang D, Tanaka K. Extracellular ATP shapes a defense-related transcriptome both independently and along with other defense signaling pathways. Plant Physiol. 2019;179:1144–1158. PMID:30630869. doi:10.1104/pp.18.01301.
- Liu X, Wu J, Clark G, Lundy S, Lim M, Arnold D, Chan J, Tang W, Muday GK, Gardner G, Roux SJ. Role for apyrases in polar auxin transport in Arabidopsis. Plant Physiol. 2012;160:1985–1995. PMID:23071251. doi:10.1104/pp.112.202887.
- Lim M, Wu J, Yao J, Gallardo I, Dugger J, Webb L, Huang J, Salmi M, Song J, Clark G, Roux S. Apyrase suppression raises extracellular ATP levels and induces gene expression and cell wall changes characteristic of stress responses. Plant Physiol. 2014;164:2054–2067. PMID:24550243. doi:10.1104/pp.113.233429.
- Zhu R, Dong X, Hao W, Gao W, Zhang W, Xia S, Liu T, Shang Z. Heterotrimeric G protein-regulated Ca2+ influx and PIN2 asymmetric distribution are involved in Arabidopsis thaliana roots’ avoidance response to extracellular ATP. Front Plant Sci. 2017;8:1522. PMID:28919907. doi:10.3389/fpls.2017.01522.
- Zhu R, Dong X, Xue Y, Xu J, Zhang A, Feng M, Zhao Q, Xia S, Yin Y, He S, Li Y, Liu T, Kang E, Shang Z. REDOX RESPONSIVE TRANSCRIPTION FACTOR1 (RRFT1) is involved in extracellular ATP regulated Arabidopsis thaliana seedling growth. Plant Cell Physiol. 2020;pii:pcaa014. PMID:32049334. doi:10.1093/pcp/pcaa014.
- Tang W, Brady S, Sun Y, Muday G, Roux S. Extracellular ATP inhibits root gravitropism at concentrations that inhibit polar auxin transport. Plant Physiol. 2003;131:147–154. PMID:12529523. doi:10.1104/pp.013672.
- Tonón C, Cecilia Terrile M, Jose Iglesias M, Lamattina L, Casalongue C. Extracellular ATP, nitric oxide and superoxide act coordinately to regulate hypocotyl growth in etiolated Arabidopsis seedlings. J Plant Physiol. 2010;167:540–546. PMID:19962212. doi:10.1016/j.jplph.2009.11.002.
- Tripathi D, Zhang T, Koo A, Stacey G, Tanaka K. Extracellular ATP acts on jasmonate signaling to reinforce plant defense. Plant Physiol. 2018;176:511–523. PMID:29180381. doi:10.1104/pp.17.01477.
- Yang X, Wang B, Farris B, Clark G, Roux S. Modulation of root skewing in Arabidopsis by apyrases and extracellular ATP. Plant Cell Physiol. 2015;56:2197–2206. PMID:26412783. doi:10.1093/pcp/pcv134.
- Matsuo M, Johnson J, Hieno A, Tokizawa M, Nomoto M, Tada Y, Godfrey R, Obokata J, Sherameti I, Yamamoto YY, Böhmer FD. High REDOX RESPONSIVE TRANSCRIPTION FACTOR1 levels result in accumulation of reactive oxygen species in Arabidopsis thaliana shoots and roots. Mol Plant. 2015;8:1253–1273. PMID:25882345. doi:10.1016/j.molp.2015.03.011.
- Wang M, Dai W, Du J, Ming R, Dahro B, Liu J. ERF109 of trifoliate orange (Poncirus trifoliata (L.) Raf.) contributes to cold tolerance by directly regulating expression of Prx1 involved in antioxidative process. Plant Biotechnol J. 2018;17:1316–1332. PMID:30575255. doi:10.1111/pbi.13056.