
ABSTRACT
Seed germination and seedling development in nearly all orchid species rely on a symbiotic relationship with mycorrhizal fungi; however, this is not the case with all mycorrhizal fungi. This study aims to provide an understanding about the important role of mycorrhiza in seed germination and growth of Paphiopedilum barbigerum. Therefore, we isolated and identified endophytic fungi from the roots of wild P. barbigerum. The beneficial mycorrhizal fungi Epulorhiza sp. FQXY019 and Tulasnella calospora FQXY017 were screened by seed symbiotic germination tests and found to promote seed germination. However, only the seeds inoculated with FQXY019 progressed from the seed germination to rooting stage. This shows that mycorrhizal fungi and P. barbigerum have a specific relation at different growth phases. In addition, we selected FQXY019 and inoculated it into MS medium, B5 medium, OMA medium, and PDA medium. The results showed that FQXY019 co-cultured on PDA significantly promoted the increase in seedling fresh weight, leaf length, and root length (p < .01). Furthermore, it significantly promoted the root number and leaf number of seedlings compared with those co-cultured on MS, B5, and OMA media and control (p < .05). Thus, this study demonstrated the promoting effect of Epulorhiza sp. FQXY019 on seed germination and seedling development, making it an alternative method for the artificial propagation of P. barbigerum.
Introduction
Under natural conditions, orchids rely on mycorrhizal fungi early in their life cycle.Citation1The seeds of the orchids are small, so a single fruit encloses millions of them.Citation2 Orchid seed lack a developed endosperm and reserves are limited to a small store of complex carbohydrates. Thus, nearly all orchid species rely on a symbiotic relationship with fungi for seed germination and protocorm development.Citation3 Depending on the presence of functional fungi in cortical cells, orchid fungi are classified as mycorrhizal and nonmycorrhizal fungi.Citation4 Mycorrhizal fungi (soil saprotrophs),Citation5 such as Ceratobasidiaceae, Tulasnellaceae, and Sebacinaceae, are collectively referred to as Rhizoctonia; they increase seed germination as well as the development rate of protocorm and seedlings of orchids.Citation6–12 However, not all mycorrhizal fungi can promote seed germination, seedling growth, and development of orchids.Citation13,Citation14 Therefore, it is essential to screen and identify beneficial mycorrhizal fungi in orchids.
Orchidaceae is a species-rich and diverse family of flowering plants comprising approximately 28,000 species in 880 genera.Citation15,Citation16 Paphiopedilum Pfitzer are famous worldwide because of their unique flower shape, attractive colors, and long flower lifespan. They are also known as slipper orchids; they have a high ornamental value and are one of the most popular genera of family Orchidaceae.Citation17,Citation18 Paphiopedilum barbigerum is native to southwest China and northern VietnamCitation19 and is one of China’s state-protected one-grade rare and endangered plants. It is an endangered species that is critically threatened by excessive harvesting, climate change, and severe habitat destruction.Citation20 The only wild population have been discovered in northern and western Guangxi and Guizhou provinces, China.Citation21 Therefore, we must understand the establishment and maintenance of the mutualistic interactions between this orchid species and its corresponding mycorrhizal fungi to conserve wild populations of the threatened species and reintroduce the plant into declining populations. Unfortunately, there are no reports on the isolation, identification, and screening of the mycorrhizal associates of P. barbigerum.
Artificial propagation technology can be used to realize the mass reproduction of Paphiopedilum,Citation22while the use of mycorrhizal fungi can help overcome the problems in the culture process, such as low survival rate, growing slowly after transplanting and poor resistance.This study aimed to isolate and identify mycorrhizal fungi from the roots of P. barbigerum. Additionally, the effects of the mycorrhizal fungi on seed germination and seedling growth of P. barbigerum were evaluated. The isolation, identification, and screening of mycorrhizal fungi are important to determine the specific and beneficial endophytic fungi species in P. barbigerum that will increase in vitro germination and seedling growth and provide fundamental guidance for the symbiotic techniques for P. barbigerum artificial propagation and conservation.
Materials and methods
As part of in vitro propagation studies, root samples of P. barbigerum were collected from their native habitat from a mountain near Fuquan County in the Guizhou Province, China, in February 2021 and the fungi were isolated from these samples.
Isolation of mycorrhizal fungi from the root of P. barbigerum: Zhu et al.’s methodCitation23
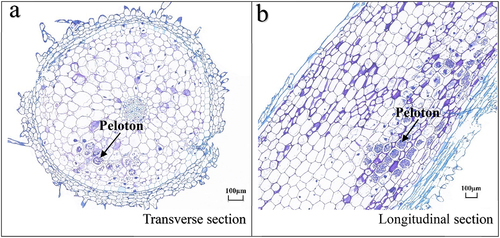
First, wild and healthy P. barbigerum root samples were chosen, rinsed under tap water for 4–6 h and cut into 4–6-cm long segments. The root segments were placed in a Petri dish filled with sterile distilled water, and root hairs, epidermis, velamen, and other attachments were gently scraped off using a scalpel on an ultra-clean workbench. Additionally, brownish zones containing pelotons were selected after microscopic examination, isolated, and rinsed three times with sterile distilled water. Then, the root segments were surface-sterilized using 70% ethanol for 30 s, 0.1% mercuric chloride for 4 min, and finally rinsed with sterile distilled water five times. The root segments with pelotons are cut 2-cm long segments. This released individual pelotons from the cortex cells, which were then placed in a 60-mm sterile Petri dish containing 10 ml sterile distilled water. Pelotons from all roots were teased three times from the exodermis to the endodermis. These were then incubated in the dark at 24°C and microscopically examined after 24 h.Second, the pelotons in sterile distilled water were placed under an optical microscope at low power. A 50-μl solution containing germinated mycelium was transferred to 1 cm2 PDA disks using a 1-ml Eppendorf micropipette and the samples were cultured at 24°C until hyphae growth was observed.
Finally, the germinated pelotons and emerging hyphae on agar disks are observed under an optical microscope. These are then cut out from the agar disks, transferred onto fresh PDA plates, and cultured at 24°C.This was repeated until pure cultures were obtained and stored at 4°C for further observations and identification.
Morphological and molecular identification of mycorrhizal fungi
The purified fungal strains were inoculated on PDA plates and incubated at 24°C. The colony characteristics of each strain were observed and recorded daily, including colony shape and color.
Purified genomic DNA of the fungal strains was extracted using the cetyltrimethylammonium bromide (CTAB) method. Take an appropriate amount of fungal mycelium and 800 µl CTAB was then added to the 2 ml centrifuge tube,ground until it was melted, incubated at 65°C for 30–40 min using a water bath.800 µl of Chloroform/Isoamyl alcohol (24:1) was added, homogenized, and centrifuged at 12,000 rpm for 10 min.Supernatant was taken and transferred to a tube, and 1 ml of 95% cold ethanol was added, shaked gently for 2 min, and incubated at −20°C for at least 30 min. It was centrifuged at 12,000 rpm for 10 min. The supernatant was removed, 1 ml of 70% ethanol was added and centrifuged at 12,000 rpm for 10 min. The supernatant was then discarded, and dried on the super clean workbench. After drying, the tube was then added 50 µl TE buffer.DNA integrity was analyzed by electrophoresis on 1% agarose gels.
The primers for ITS sequencing of the fungi were ITS1 (5′-TCCGTAGGTGAAC
CACCTGCGG −3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′). Polymerase chain reaction (PCR) was performed in a 30-μL reaction mixture solution containing 1 μL of DNA, 1 μL each of the downstream and upstream primers, 12 μL of double-distilled water, and 15 μL of 2× Power Taq PCR Master Mix (Beijing Baitaike Biotechnology Co., Ltd). The amplification program comprised initial denaturation at 95°C for 5 min, followed by 35 cycles of denaturing at 95°C for 30 s, annealing at 58°C for 30 s, extension at 72°C for 1 min by 35 cycles, and a final extension for 5 min at 72°C. The amplified fragments were purified and sequenced by an external service provider (Tianyihuiyuan Inc., China). The sequences were used to perform a BLAST search against the National Center for Biotechnology Information (NCBI) sequence database (GenBank) to compare the sequences with the most similarity to identify each endophytic fungus.
Phylogenetic analysis
The sequences of FQXY019 and FQXY017 were subjected to BLAST in the NCBI BLAST similarity search tool to find the closest match from the GenBank database. Phylogenetic trees were constructed using the neighbor-joining method on the basis of the available DNA data obtained during this study.
Symbiotic germination
The mycelium of each isolated endophytic fungi was inoculated onto 90-mm Petri dishes containing 30 mL of PDA medium (200 g potato, 10 g agar, 20 g glucose, and 1 L distilled water). This was repeated three times. After culturing for 2 weeks, it will be used for symbiotic seed germination tests.
Symbiotic germination tests were performed using the green fruit capsules of P. barbigerum. First, the fruit capsules were surface-sterilized using 70% ethanol for 30 s, followed by 0.1% mercuric chloride for 6 min, and rinsed with sterile distilled water five times. Then, the seeds were scooped out of the sterilized pods using a sterile scalpel and forceps.Approximately 100 seeds were sown in each fungus-PDA medium as described above. The plates were sealed and cultured in a 18-h light and 6-h dark cycle at 22°C-24°C and a light intensity of 1200 lux. After inoculation and culture for 2 weeks, seed germination, protocorm formation, and seedling development were monitored. The average of the three replicates of plate cultures was calculated in subsequent analyses. The germination rate was calculated as follows: the number of germinated seeds/number of inoculated seeds × 100%.
Effects of mycorrhizal fungus co-cultured with different media on seedling growth of P. barbigerum
The test seedlings were obtained from the symbiotic germination of seeds and mycorrhizal fungi. The mycorrhizal fungi and seedlings were co-cultured on six replicate plates of each treatment (I.e., Murashige and Skoog medium, Gamborg’s B5 medium, OMA medium, and PDA medium).First, the four abovementioned media were inoculated with a 0.5 × 1 cm2 block of FQXY019 into each tissue culture bottle,cultured in the dark at 28°C. Media without treatment (without the treatment of FQXY019) were considered as the control (CK).After 2 weeks of culture, the test seedlings were inoculated into tissue culture bottles. Finally, the bottles were sealed and incubated in 18-h light and 6-h dark cycle at 22°C–24°C at a light intensity of 1200 lux.
Statistical analysis
The SPSS (Statistical Package for the Social Sciences v.21.0) software was used for statistical analysis. Data were statistically analyzed using the single-factor analysis of variance method; significant differences between the treatments were determined using multiple comparisons using LSD (p< 0.05, p< 0.01).
Results
Isolation and identification of mycorrhizal fungi from P. barbigerum
Observation under an optical microscope revealed that the pelotons were unevenly distributed in the root cortex cells of wild P. barbigerum seedlings (). According to Bernard,Citation24 peloton formation in the culture that characterized Rhizoctonia-like orchid mycorrhizae was also produced by Fusarium isolated from the orchid.Citation25 Endophytic fungi were isolated from the root at different frequencies; 127 strains of endophytic fungi isolates were obtained in pure culture (), mostly belonging to the Sordariomycetes, Agaricomycetes, and Leotiomycetes class. These are Fusarium nematophilum, Fusarium sp., Fusarium solani, Fusarium redolens, Epulorhiza sp., Pezicula ericae, Dactylonectria novozelandica, Chaetomium nigricolor, and Tulasnella calospora (). Fusarium sp., Epulorhiza sp., and Tulasnella sp. accounted for 56.7%, 28.3%, and 11.8% of the total strains, respectively.
Table 1. Occurrence and frequency of the endophytic fungi isolates
Table 2. Identification of ITS sequences of endophytic fungi isolates from Paphiopedilum barbigerum roots
Figure 1. Longitudinal and transverse profiles of the roots of Paphiopedilum barbigerum grown from wild seedlings. Arrows represents peloton (a,b).
Symbiotic seed germination of P. barbigerum
This experiment evaluated the effects of endophytic fungi on promoting in vitro symbiotic seed germination. The results showed that the seeds inoculated with FQXY019 after 15 d of culture on PDA plates turned light brownish green. The seed coat ruptured due to enlarged embryos and protocorm-like bodies were formed after 30 d of culture (). Further, the first leaf emerged after 50 d of culture () and further development and rooting occurred after 80 d of culture (). Seeds inoculated with FQXY017 turned light brownish green after 40 d of culture on PDA, and the seed coat ruptured due to the enlarged embryo (). Further, protocorm-like bodies formed after 55 d of culture () and the coleoptile emerged after 75 d of culture (). Additionally, further development and browning after 90 d of culture could not be achieved (). However, seeds inoculated with FQXF001, FQXF002, FQXF005, FQXF006, FQXF008, FQXF016, FQXF015, FQXF042, FQXF045, and FQXF047 could absorb water and expand but not break through the testa on PDA (). Few seeds germinated and broke through the seed coat without inoculation but could not further develop CK () (). As a results, FQXY019 and FQXY017 could promote seed germination. However, seeds inoculated with FQXY019 germinated and formed protocorms 25 days earlier than FQXY017, respectively.Notably, only seeds inoculated with FQXY019 on PDA plates could promote seedling formation and rooting, whereas FQXY017 could not.
Table 3. Germination of Paphiopedilum barbigerum seeds inoculated with endophytic fungi cultured on PDA plates
Figure 2. Protocorm-like bodies were formed after 30 d with FQXY019 (a). The first leaf emerged after 50 d with FQXY019 (b). Rooting occurred after 80 d with FQXY019 (c).Turned light brownish green and seed coat ruptured after 40 d with FQXY017 (d). Protocorm-like bodies formed after 55 d with FQXY017 (e). The coleoptile emerged after 75 d with FQXY017 (f). Browning after 90 d with FQXY017 (g). Seeds expand but not break through the testa after 90 d with FQXY001 (h). Seeds expand but not break through the testa after 90 d with FQXY005 (i). Seeds germinated and broke through the seed coat but could not further develop after 90 d (CK) (j).

Normal sterilized P. barbigerum seeds were used as control (CK)
Effects of mycorrhizal fungus co-cultured with different media on seedling growth of P. barbigerum
We selected the beneficial mycorrhizal fungus FQXY019 to be inoculated into four co-culturing media to determine the best medium for co-culturing mycorrhizal fungus and seedlings. After 90 d of culturing, FQXY019 co-cultured with PDA medium significantly increased seedling fresh weight, leaf length, and root length compared with MS, B5, OMA medium, and CK (p < .01). Furthermore, PDA significantly promoted root and leaf numbers compared with MS, B5, OMA medium, and CK (p < .05). Remarkably, the root length increments of P. barbigerum seedlings were significantly inhibited by MS and B5 media (p < .01 and p < .05, respectively) (). Thus, the best co-culture medium was the PDA medium. The co-culture of different media and mycorrhizal fungi had different effects on the growth of P. barbigerum seedlings.
Figure 3. Effects of the mycorrhizal fungus FQXY019, co-cultured with different media on seedling growth of Paphiopedilum barbigerum.
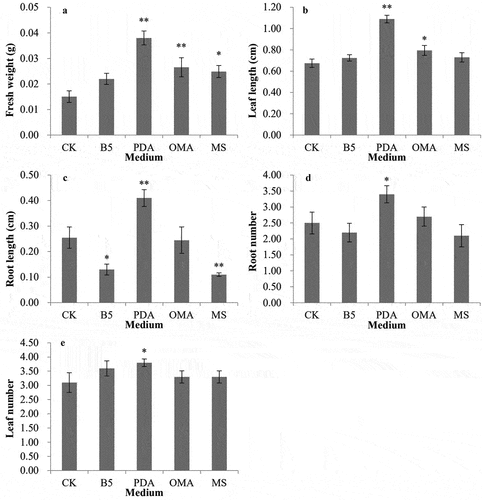
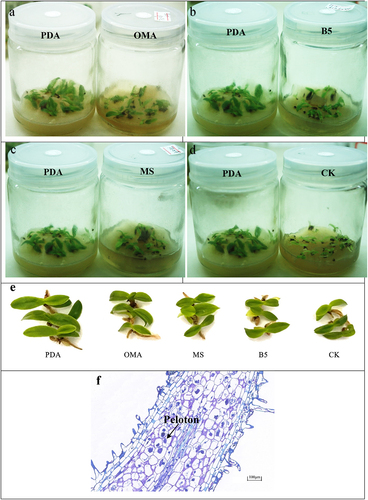
Figure 4. The growth state of Paphiopedilum barbigerum seedlings inoculated with FQXY019.Symbiotic mycorrhizal fungal and seedlings cultured on PDA, OMA, B5,MS and CK (a,b,c,d). Comparison of seedlings growth of five treatments (e). Longitudinal section of seedling root inoculated with FQXY019 on PDA (f).
*represents significant differences (p< .05), ** represents extremely significant differences (p < .01)
Morphological and molecular identification of beneficial mycorrhizal fungi FQXY019 and FQXY017
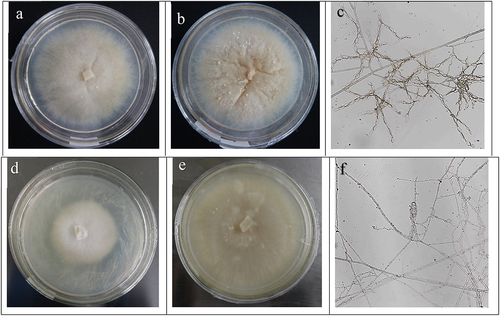
We screened out the beneficial mycorrhizal fungi FQXY019 and FQXY017 that promote seed germination of P. barbigerum by symbiotic seed germination test. Therefore, this study focuses on the morphological and molecular identification of the two strains on the PDA medium observed for FQXY019. Results showed that colonies were creamy-yellow and center-ring milky-white with irregular and serrated edges; hyphae grew close to the medium without aerial hyphae in its early stage (). Later, colonies became waxy and brown, had center-ring depression and small spherical bulges (). Hyphal branching occurred in the right- or near-right-angle (5.39–10.04 μm diameter); monilioid cells were oval, branched monilioid cell chains (16.79–23.82 × 34.35–45.38 μm) composed of more than 6–13 monilioid cells, which clustered to develop compact mass clusters of monilioid cell chains that form tufts (). PDA medium was observed for FQXY017, and colonies were light-yellow and a milky-white center-ring with velvet and even edges (). Later, colonies became creamy-yellow and waxy (). Hyphal branching occurred in the right- or near-right-angle (5.58–8.54 μm diameter); monilioid cells were long-cylinders with branched monilioid cells chains (5.48–9.76 × 13.21–28.31 μm) composed of more than 4–9 monilioid cells, which clustered and developed loose mass clusters ().
Figure 5. The morphology of the colony and mycelium of the mycorrhizal fungi FQXY019 and FQXY017. FQXY019 colonies were at the early stage (a); FQXY019 colonies were at the later period (b); mycelium of FQXY019 (c); FQXY017 colonies were at the early stage (d); FQXY017 colonies were at the later period (e); mycelium of FQXY017 (f).
The ITS sequence of the mycorrhizal fungi FQXY019 and FQXY017 exhibited the highest similarity to Epulorhiza sp. (Epulorhiza sp., GU166409.1) and Tulasnella calospora (Tulasnella calospora, GU166406.1), respectively. Furthermore, a phylogenetic tree was constructed using the ITS sequence of mycorrhizal fungi in Epulorhiza sp. and T. calospora. Finally, the best phylogenetic tree was constructed using the neighbor-joining method ().
Figure 6. Phylogenetic tree based on ITS-rDNA sequence of mycorrhizal fungi isolated from Paphiopedilum barbigerum and the closest identified relatives from GenBank.

Discussion
P. barbigerum is a wild plant species with an extremely small population in China. It only grows in the limestone hills in northern Guangxi and Guizhou, China. This study collected the root segments of P. barbigerum epiphytic on limestone in Guizhou with a shallow soil layer and poor habitat. Altogether, 127 endophytic fungi were isolated from P. barbigerum roots. Among them, the dominant genera were Epulorhiza, Fusarium, and Tulasnella. Tulasnella and Epulorhiza were considered universal orchid symbionts,Citation26 i.e., they can form mycorrhizal fungi with most orchids.Citation27–37 Studies have reported that Fusarium is the dominant species of P. armeniacum, P. micranthum, and Bletilla striata.Citation19,Citation28,Citation38 Additionally, Fusarium sp. has been recorded as an endophytic fungus.Citation39,Citation40 Such studies have reported that Epulorhiza, Fusarium, and Tulasnella are the mycorrhizal fungi of orchids,Citation11,Citation41–43 consistent with the results obtained in our study.
Recognition processes are an essential part of most symbiotic associations,Citation44but orchids with broad webs of fungi lack these recognition processes, which only germinate and grow in the presence of a suitable fungus.Citation45 According to this, our study analyzes and tests their roles in seed germination. Furthermore, The results of our in vitro study indicate that Epulorhiza sp. FQXY019 and T. calospora FQXY017 could promote seed germination and protocorm formation. However, only FQXY019 could promote seedling formation and rooting, whereas FQXY017 could not. The seeds germinate in the presence of fungus, but many of the esulting protocorms abort because of physiological imbalances with these fungus.Citation45 Because only fully compatible fungus resulted in orchid seedling growth.This result agrees with the reported findings that strains M2, M4, and HR1-1 stimulate seed germination and protocorm formation but do not promote seedling growth in B. striata.Citation11 Although Epulorhiza isolated from P. integra promoted seed germination of P. integra in vitro, its development was blocked when the protocorms grew with these mycorrhizal fungi.Citation46 These conclusions may be explained because different mycorrhizal fungi play different roles in seed germination and subsequent seedling growth stages of P. barbigerum.
Other isolated endophytic fungi besides Rhizoctonia, such as F.nematophilum, F. solani, F. redolens, Fusarium sp., P. ericae, D. novozelandica, C. nigricolor could not promote seed germination on PDA medium. The seed germination rate was lower than that of CK. Peschke and Volz defined Fusarium as one of the most ubiquitous and interesting orchid endophytes,Citation47 since they could promote seed germination and several species could cause diseases. Previous studies have identified that several Fusarium species do not make suitable symbionts for orchids, and some are pathogenic.Citation48 F. solani, F. oxysporum and F. proliferatum cause the some disease in orchids.Citation49 Under optimal growth conditions, F. solani as a virulent species, tend to asymptomatic endophyte rather than pathogens.Citation50 However, Fusarium species enhance seed germination and protocorm development in some orchids.Citation40,Citation42,Citation51 Although this effect of promoting germination and growth was relatively minor compared with that of specific Rhizoctonia-like orchid mycorrhiza.Citation25 In a recent study, F.oxysporum KB-3 significantly promoted the vegetative growth of Bletilla striata and Dendrobium candidum, typical orchid mycorrhizal structures were observed in their roots.Citation52 Endophytic Fusarium promoted seed germination in Cypripedium and Platanthera.Citation53 As reported, fungus-plant interactions can easily shift from mutualism to parasitism depending on the plant’s physiology and environmental conditions.Citation54 These studies showed that it is necessary to explore the role of endophytic and nonpathogenic Fusarium strains in controlling pathogens and promoting germination and development of orchid species.
Furthermore, this study compared the effectiveness of four media co-cultured with the compatible mycorrhizal fungus Epulorhiza sp. FQXY019 on seedling growth and development. The promoting effects of different media on seedling development of P. barbigerum were unequal. The seedlings were inoculated on PDA, and those co-cultured with FQXY019 developed best compared with those inoculated on MS, B5, OMA, and CK media. The average fresh weight, leaf length, root length, root number, and leaf number of seedlings among these treatments had significant or extremely significant differences. PDA medium has low carbon and nitrogen content and poor nutrition compared with the other three media, which agrees with the characteristics of the shallow soil layer and poor nutrition in the field habitat of P. barbigerum. Additionally, mycorrhizal fungi can promote the growth of Paphiopedilum in nutrient-poor media, which agrees with the findings of previous studies.Citation55 It is suggested that symbiosis with mycorrhizal fungi may be a reason for P. barbigerum to adapt to harsh habitats. Furthermore, it was observed from the experiments that the mycelium of the fungus FQXY019 on PDA medium grew vigorously and had dense colonies. Further, the hyphae were observed under the microscope with many monilioid cells, while the colony surfaces of MS, B5, and OMA media were sparse. This is probably because the FQXY019 exhibits a high diversity of morphological and growth characteristics on the PDA mediumCitation56 and that the fungus supplies carbohydrates, vitamins, or other growth factors to seedlings. These results indicate that FQXY019 is a beneficial orchid mycorrhiza in seed germination and subsequent seedling growth stages of P. barbigerum.
Symbiotic (co-culture with mycorrhizal fungi) seed germination and seedling development methods require a relatively simple medium formula, which is cost-effective and time-saving. Furthermore, symbiotic plantlets may have more advantages than asymbiotic plantlets because seedlings inoculated with mycorrhizal fungi grow better and have stronger resistance.Citation57,Citation58 Research shows that Epulorhiza sp. increased nutrient intake in plants and increased seed germination on Dendrobium nobile and Dendrobium chrysanthum.Citation16 Epulorhiza repens ML01 can successfully establish and survive in pot orchids under greenhouse conditions, and reduce the occurrence of diseases in the Cymbidium hybridum roots.Citation59 This study demonstrated the promoting effect of Epulorhiza sp. FQXY019 on seed germination and seedling development. Results showed that FQXY019 forms a good symbiotic system with P. barbigerum seed and seedlings and can be used as symbiotic mycorrhizal fungi for the growth and reproduction of P. barbigerum culture seedlings. Therefore, this this can be used as an alternative method for the artificial propagation of P. barbigerum.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- McCormick M, Burnett R, Whigham D. Protocorm-supporting fungi are retained in roots of mature Tipularia discolor orchids as mycorrhizal fungal diversity increases. Plants. 2021;10(6):1251–11. doi:10.3390/plants10061251.
- Chen J, Wang H, Liu SS, Li YY, Guo ,SX. Ultrastructure of symbiotic germination of the orchid Dendrobium officinale with its mycobiont, Sebacina sp. Aust J Bot. 2014;62(3):229–234. doi:10.1071/BT14017.
- Zhao D-K, Selosse M-A, Wu LM, Luo Y, Shao S-C, Ruan Y-L. Orchid reintroduction based on seed germination-promoting mycorrhizal fungi derived from protocorms or seedlings. Front Plant Sci. 2021;12:e701152. doi:10.3389/fpls.2021.701152.
- Li TQ, Wu SM, Yang W, Selosse M-A, Gao JY. How mycorrhizal associations influence orchid distribution and population dynamics. Front Plant Sci. 2021;12:e647114. doi:10.3389/fpls.2021.647114.
- Selosse M-A, Martos F. Do chlorophyllous orchids heterotrophically use mycorrhizal fungal carbon? Trends Plant Sci. 2014;19(11):683–685. doi:10.1016/j.tplants.2014.09.005.
- Suwannarach N, Kumla J, Rachanarin C, Srimuang KO. In vitro symbiotic seed germination of Epipactis flava (Orchidaceae) promoted by endophytic fungus, Tulasnella phuhinrongklaensis. Chiang Mai J Sci. 2021;48:787–792.
- Chutima R, Dell B, Vessabutr S, Bussaban B, Lumyong S. Endophytic fungi from Pecteilis susannae (L.) Rafin (Orchidaceae), a threatened terrestrial orchid in Thailand. Mycorrhiza. 2011;21:221–229. doi:10.1007/s00572-010-0327-1.
- Guimaraes FAR, Pereira MC, Felicio CD, Torres DP, Oliveira SF, Veloso TGR, Kasuya MCM. Symbiotic propagation of seedlings of Cyrtopodium glutiniferum Raddi (Orchidaceae). Acta Bot Bras. 2013;27(3):590–596. doi:10.1590/S0102-33062013000300016.
- Jiang JH, Lee YI, Cubeta MA, Chen LC. Characterization and colonization of endomycorrhizal Rhizoctonia fungi in the medicinal herb Anoectochilus formosanus (Orchidaceae). Mycorrhiza. 2015;25:431–445. doi:10.1007/s00572-014-0616-1.
- Herrera H, Valadares R, Contreras D, Bashan Y, Arriagada C. Mycorrhizal compatibility and symbiotic seed germination of orchids from the Coastal Range and Andes in south central Chile. Mycorrhiza. 2017;27(3):175–188. doi:10.1007/s00572-016-0733-0.
- Fuji M, Miura C, Yamamoto T, Komiyama S, Suetsugu K, Yagame T, Yamato M, Kaminaka H. Relative effectiveness of Tulasnella fungal strains in orchid mycorrhizal symbioses between germination and subsequent seedling growth. Symbiosis. 2020;81(1):53–63. doi:10.1007/s13199-020-00681-0.
- Tsutsumi C, Tsutsumi T, Yukawa T. In vitro seed propagation and conservation of the rediscovered rare Liparis hostifolia (Orchidaceae). Bull Natl Mus Nat Sci, Ser B. 2020;46:111–118.
- Smith SE, Read DJ. Mycorrhizal symbiosis (Third Edition). New York,USA: Academic Press; 2008.
- Xi GJ, Shi J, Li JB, Han ZM. Isolation and identification of beneficial orchid mycorrhizal fungi in Bletilla striata (Thunb.) Rchb.f. (OrchidaceaeRchb.f.(Orchidaceae). Plant Signaling Behav. 2020;15(12):e1816644. doi:10.1080/15592324.2020.1816644.
- Chase MW, Cameron KM, Freudenstein JV, Pridgeon AM, Salazar G, van Den Bert C, Schuiteman A. An updated classification of Orchidaceae. Bot J Linn Soc. 2015;177(2):151–174. doi:10.1111/boj.12234.
- Tsulsiyah B, Farida T, Sutra CL, Semiarti E. Important Role of Mycorrhiza for Seed Germination and Growth of Dendrobium Orchids. J Trop Biodiv Biotech. 2021;6(2):jtbb60805. doi:10.22146/jtbb.60805.
- Feng J-Q, Huang W, Wang J-H, Zhang S-B. Different strategies for photosynthetic regulation under fluctuating light in two sympatric Paphiopedilum species. Cells. 2021;10(6):e1451. doi:10.3390/cells10061451.
- Liu ZJ, Chen SC, Chen LJ, Lei SP. The genus Paphiopedilum in China. Beijing,China: Science Press; 2009. 4–12.
- Shi J, Luo Y-B, Bernhardt P, Ran J-C, Liu Z-J, Zhou Q. Pollination by deceit in aphiopedilum barbigerum (Orchidaceae): a staminode exploits the innate colour preferences of hoverflies (Syrphidae). Plant Biol. 2009;11(1):17–28. doi:10.1111/j.1438-8677.2008.00120.x.
- Liu ZJ, Chen SC, Chen LJ, Lei SP. The genus Paphiopedilum in China. Beijing,China: Science Press; 2009.
- Long J, Qin FX, Zhou JW, Ran JC, Qin LJ. A study on microhabitat parameters of the endangered species Paphiopedilum barbigerum. J Guizhou Norm Univ (Natural Sciences). 2009;27:15–20. doi:10.3969/j.1004-5570.2009.03.003.
- Pant B, Shah S, Shrestha R, Pandey S, Joshi PR. An Overview on Orchid Endophytes. Cham,Switzerland: Springer; 2017.
- Zhu GS, Yu ZN, Gui Y, Liu ZY. A novel technique for isolating orchid mycorrhizal fungi. Fungal Divers. 2008;33:123–137.
- Bernard N. L’e ´volution dans la symbiose. Les orchidées et leur champignons commenseux. Ann Sci Nat Bot Ser 1909;9:1–196.
- Vujanovic V, St-Arnaud M, Barabe D, Thibeault G. Viability testing of orchid seed and the promotion of colouration and germination. Ann Bot. 2000;86(1):79–86. doi:10.1006/anbo.2000.1162.
- Freitas EFS, Sliva M, Cruz ES, Mangaravite E, Bocayuva MF, Veloso TGR, Selosse M-A, Kasuya MCM. Diversity of mycorrhizal Tulasnella associated with epiphytic and rupicolous orchids from the Brazilian Atlantic Forest, including four new species. Sci Rep. 2020;10(1):e7069. doi:10.1038/s41598-020-63885-w.
- Yuan L, Yang ZL, Li S-Y, Hu H, Huang J-L. Mycorrhizal specificity, preference, and plasticity of six slipper orchids from South Western China. Mycorrhiza. 2010;20(8):559–568. doi:10.1007/s00572-010-0307-5.
- Tian F, Liao XF, Wang LH, Zhu GS, Gui Y, Bai XX. Classification and identifition of mycorrhizal fungi of Paphiopedilum micranthum. North Hortic. 2017;24:116–122. doi:10.11937/bfyy.20171417.
- Rasmussen HN. Terrestrial orchids from seed to mycotrophic plant. New York: Cambridge University Press; 1995.
- Pereira OL, Rollemberg CL, Borges AC, Matsuoka K, Kasuya MCM. Epulorhiza epiphytica sp. nov. isolated from mycorrhizal roots of epiphytic orchids in Brazil. Mycoscience. 2003;44(2):153–155. doi:10.1007/s10267-002-0087-7.
- Pereira MC, Rocha DI, Veloso TGR, Pereira OL, Francino DMT, Meira RMS, Kasuya MCM. Characterization of seed germination and protocorm development of Cyrtopodium glutiniferum (Orchidaceae) promoted by mycorrhizal fungi Epulorhiza spp. Acta Bot Bras. 2015;29(4):567–574. doi:10.1590/0102-33062015abb0078.
- Ma M, Tan TK, Wong SM. Identification and molecular phylogeny of Epulorhiza isolates from tropical orchids. Mycol Res. 2003;107(9):1041–1049. doi:10.1017/S0953756203008281.
- Látalová K, Baláž M. Carbon nutrition of mature green orchid Serapias strictiflora and its mycorrhizal fungus Epulorhiza sp. Biol Plant. 2010;54(1):97–104. doi:10.1007/s10535-010-0014-2.
- Swangmaneecharern P, Serivichyaswat P, Nontachaiyapoom S. Promoting effect of orchid mycorrhizal fungi Epulorhiza isolates on seed germination of Dendrobium orchids. Sci Hortic. 2012;148:55–58. doi:10.1016/j.scienta.2012.09.013.
- Linde CC, May TW, Phillips RD, Ruibal M, Smith LM, Peakall R. New species of Tulasnella associated with terrestrial orchids in Australia. IMA Fungus. 2017;8(1):27–47. doi:10.5598/imafungus.2017.08.01.03.
- Tan XM, Wang CL, Chen XM, Zhou YQ, Wang YQ, Luo AX, Liu ZH, Guo SX. In vitro seed germination and seedling growth of an endangered epiphytic orchid, Dendrobium officinale, endemic to China using mycorrhizal fungi (Tulasnella sp.). Sci Hortic. 2014;165:62–68. doi:10.1016/j.scienta.2013.10.031.
- Arifin AR, May TW, Linde CC. New species of Tulasnella associated with Australian terrestrial orchids in the Cryptostylidinae and Drakaeinae. Mycologia. 2020;13:212–230. doi:10.1080/00275514.2020.1813473.
- Li M, Zhang Z. Studies and applications on mycorrhiza of Paphiopedilum armeniacum. J Biol. 2001;18:17–18. doi:10.3969/j.2095-1736.2001.06.007.
- Rasmussen HN. Recent developments in the study of orchid mycorrhiza. In Smith SE, and Smith FA (Eds.), Diversity and Integration in Mycorrhizas (Proceedings of the 3rd International Conference on Mycorrhizas) Adelaide; 2001. p. 149–163. doi: 10.1007/978-94-017-1284-2_15
- Behera D, Tayung K, Mohapatra UB. PCR-based identification of endophytes from three orchid species collected from similipal biosphere reserve, India. American International Journal of Research in Formal, Applied & Natural Sciences (AIJRFANS). 2013;3:10–17.
- Jiang J, Zhang K, Cheng S, Nie Q, Zhou S, Chen Q, Zhou J, Zhen X, Li XT, Zhen TW, et al. Fusarium oxysporum KB-3 from Bletilla striata: an orchid mycorrhizal fungus. Mycorrhiza. 2019;29(5):531–540. doi:10.1007/s00572-019-00904-3.
- Chen J, Wang H, Guo S-X. Isolation and identification of endophytic and mycorrhizal fungi from seeds and roots of Dendrobium (Orchidaceae). Mycorrhiza. 2012;22(4):297–307. doi:10.1007/s00572-011-0404-0.
- Di Pardo AF, Chiocchio VM, Barrera V, Colombo RP, Martinez AE, Gasoni L, Godeas AM. Mycorrhizal fungi isolated from native terrestrial orchids of pristine regions in Cordoba (Argentina). Rev Biol Trop. 2015;63(1):275–283. doi:10.15517/rbt.v63i1.14226.
- Radutoiu S, Madsen LH, Madsen EB, Felle HH, Umehara Y, Gronlund M, Sato S, Nakamura Y, Tabata S, Sandal N, et al. Plant recognition of symbiotic bacteria requires two LysM receptor-like kinases. Nature. 2003;425(6958):585–592. doi:10.1038/nature02039.
- Bonnardeaux Y, Brundrett M, Batty A, Dixon K, Koch J, Sivasithamoaram K. Diversity of mycorrhizal fungi of terrestrial orchids: compatibilitywebs, brief encounters, lasting relationships and alien invasions. Mycol Res. 2007;111(1):51–61. doi:10.1016/j.mycres.2006.11.006.
- Zettler LW, Delaney STW. Symbiotic seed germination of an orchid in decline (Platanthera integra) from the green swamp, North Carolina. Castanea. 2000;65:207–212.
- Peschke HC, Volz PA. Fusarium moniliforme shield: association with species of orchids. Phytologia. 1978;40:347–356.
- Srivastava S, Kadooka C, Uchida JY. Fusarium species as pathogen on orchids. Microbiol Res. 2018;207:188–195. doi:10.1016/j.micres.2017.12.002.
- Latiffah Z, Hayati MZN, Baharuddin S, Maziah Z. Identification and pathogenicity of Fusarium species associated with root rot and stem rot of Dendrobium. Asian J Pl Pathol. 2009 ;3(1):14–21. doi:10.3923/ajppaj.2009.14.21.
- Ma X, Kang J, Nonachaiyapoom S, Wen T, Hyde KD. Non-mycorrhizal endophytic fungi from orchids. Special section:Endophyte. Curr Sci. 2015;108:1–16.
- Bayman P, Otero JT. Microbial Endophytes of Orchid Roots. In: Shultz BJE, Boyle CJC, and Sieber TN, editors. Microbial Root Endophytes. Berlin,Germany: Springer; 2006. p. 153–177. doi:10.1007/3-540-33526-9_9.
- Jiang JW, Zhang K, Cheng S, Nie QW, Zhou S-X, Chen QQ, Zhou JL, Zhen X, Li XT, Zhen TW, et al. Fusarium oxysporum KB-3 from Bletilla striata: an orchid mycorrhizal fungus. Mycorrhiza. 2019;29(5):531–540. doi:10.1007/s00572-019-00904-3.
- Sufaati S, Agustini V, Suharno. Short Communication: fusariumas endophyte of some terrestrial orchid from Papua, Indonesia. Biodiversitas. 2016;17:366–371.
- Chen D-Y, Wang X-J, Li T-Q, Li N-Q, Gao J-Y. In situ seedling baiting to isolate plant growth-promoting fungi from Dendrobium officinale, an over-collected medicinal orchid in China. Global Ecology and Conservation.2021. 2021;28:e01659. doi:10.1016/j.gecco.2021.e01659.
- Zhu XM, Hu H, Li SY, Yan N. Interaction between endophytic fungi and seedlings of two species of Paphiopedilum during symbiotic culture. Plant Diversity and Resources. 2012;34:171–178. doi:10.3724/SP.J.1143.2012.11144.
- Liu L, Li XZ, Guo CH, Li CJ. Effects of different medium on growth and sporulation of asexual stage of Epichloё endophytes. Pratacultural Science. 2014;32:859–869. doi:10.3389/fmicb.2017.02488.
- Takahashi K, Kumagai H, Ogiwara I. Effect of Orchid Mycorrhizal Fungi on the Growth of Daughter Tubers in Habenaria radiate (Thumnb.) K. Spreng. Plantlets Raised from Tubers in Vitro. Hortic Res (Japan) 2006;5(1):13–17. doi:10.2503/hrj.5.13.
- Chang DCN. The screening of orchid mycorrhizal fungi (OMF) and their applications. In: Chen WH, and Chen HH, editors. Orchid Biotechnology. Singapore,Singapore: World Scientific; 2007. p. 77–98. doi:10.1142/9789812775900_0005.
- Liu S, Liu M, Liao QG, Lü FB, Zhao XL. Effects of inoculated mycorrhizal fungi and non-mycorrhizal beneficial microorganisms on plant traits, nutrient uptake and root-associated fungal community composition of the Cymbidium hybridum in Greenhouse. J Appl Microbiol. 2020:14967. doi:10.1111/jam.