Abstract
In material from the northern Mid-Atlantic Ridge and the Reykjanes Ridge, five species of deep-sea mushroom corals belonging to the genera Anthomastus, Heteropolypus and Pseudoanthomastus were found, including three species new to science. The scope and diagnoses of the genera Heteropolypus and Pseudoanthomastus are re-evaluated based on examination of the type material. The main characteristics to distinguish these three deep-sea genera are tentacular and pharyngeal sclerites. A term sclerome as an inventory of all sclerites of the actual octocoral taxon is proposed. A replacement name Alcyonium pasternaki is proposed for the monomorphic Pseudoanthomastus pacificus Pasternak, 1981. Type materials of Anthomastus grandiflorus Verrill, A. agaricus Studer, A. canariensis Write & Studer, A. purpureus (Koren & Danielssen) and A. agassizii Verrill, all species described or reported from the North Atlantic, confirmed the validity of these species.
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
Introduction
It is difficult to find a genus of soft coral so easily distinguished by its external appearance (even on underwater images) as Anthomastus Verrill, Citation1878. Since the first description of Anthomastus grandiflorus Verrill, Citation1878, the unusual morphology of these corals has attracted attention. Images of red colonies with huge expanded autozooids can be found in most modern colourful books about life in the deep-sea. Species of this genus have been reported from all over the world. To date, 27 nominal species have been described in the genus (). However, most of them are known from a single locality and even from a single specimen.
Table I. All nominal species of the genus Anthomastus, with current taxonomic status and type localities.
By the beginning of the twentieth century, four species of Anthomastus had been described from the North Atlantic: Anthomastus grandiflorus Verrill, Citation1878, A. purpureus (Koren & Danielssen, 1883), A. canariensis Wright & Studer, 1889 and A. agaricus Studer, Citation1890. In 1927 Jungersen suggested all species of the genus Anthomastus known at that time from the North Atlantic to be junior synonyms of A. grandiflorus based exclusively on material from the Danish Ingolf-Expedition to Iceland and Greenland waters. Several growth forms were pictured in the paper, showing a clear relationship between the type of substrate and colony morphology of the coral. It was a turning point in the interpretation of A. grandiflorus, and after that most identifications were based mainly on the colour illustrations of Jungersen (Citation1927). In spite of a review by d'Hondt (Citation1992) demonstrating the validity of A. canariensis and A. purpureus, in some faunistic lists A. grandiflorus is still considered to be the only species of genus Anthomastus in European seas (van Ofwegen et al. Citation2001), and together with A. agassizii Verrill, Citation1922 (Moore et al. Citation2003, Watling & Auster Citation2005, Watling et al. Citation2011), is considered one of the only two valid species of the genus in the North Atlantic. The present article deals with the North Atlantic set of dimorphic and trimorphic alcyoniids with emphasis on species recorded from the Mid-Atlantic Ridge (MAR).
Material and methods
Material was obtained on cruises to the Reykjanes Ridge, the MAR north of the Azores and adjacent areas by several different research vessels (). On the RRS Challenger an epibenthic sledge as described by Aldred et al. (Citation1976) was used. On the RV Akademik Mstislav Keldysh sampling was done using a Sigsbee trawl (a modification of Agassiz trawl with frames 2.5 and 1.5 m wide) and a cylindrical geological dredge (Kidd et al. Citation1990). The RV G.O. Sars cruise used a modification of a semi-commercial otter trawl (the Campelen 1800 shrimp trawl), with mouth opening 17×12×4.5 m, with a 5-mm mesh liner at the cod end (Wenneck et al. Citation2008). On the ECOMAR cruises of the RRS James Cook sampling was done by semi-balloon otter trawl (OTSB) towed on a single warp and, ROV Isis (Priede et al. Citation2013). Specimens were fixed in 4% buffered formalin or preserved in 80% alcohol. Formalin-fixed materials were transferred to 80% alcohol within one month. All measurements were made on preserved specimens. For sclerite studies, pieces of soft tissue were dissolved in sodium hypochlorite solution (household bleach) and rinsed several times times in distilled water. Clean sclerites were transferred with pipette to microscope slides and dried. Permanent preparations were made using Canada balsam or Durcupan ACM mounting media (Fabricius & Alderslade Citation2001: 39–40). Multifocus images of sclerites were obtained with manual focus compound microscope (Leica DM5000B) with photo attachment (Leica DFC 490) using LAS Leica Microsystems Software.
Table II. Material.
Terminology is used in accordance with Illustrated Trilingual Glossary (Bayer et al. Citation1983). Up to now, no appropriate term has been introduced encompassing all sclerites of octocoral species. For this purpose I propose here a term sclerome be defined as an inventory of all sclerites of the octocoral taxon.
Holotypes of new species are deposited in ZMMU and NHMUK.
Comparative material examined
Anthomastus agassizii Verrill, Citation1922: holotype MCZ 4087; Anthomastus agaricus Studer, Citation1890: holotype MOM INV-6080 (120016); Anthomastus canariensis Wright & Studer, 1889: holotype NHMUK 1889.5.27.127; Anthomastus fisheri Bayer, 1952: holotype NHMUK 49623; Anthomastus grandiflorus Verrill, Citation1878: syntypes USNM 30181, YPM 4853 (photos and sclerites only); A. japonicus Nutting, 1906: syntypes USNM 30181; Anthomastus ritteri Nutting, 1909: holotype USNM 25422 (photos and sclerites only); A. rylovi Naumov, 1952: syntypes ZIN 10599; A. steenstrupi Wright & Studer, 1889: holotype NHMUK 1889.5.27.128; Heteropolypus insolitus Tixier-Durivault, 1964: holotype ZMUC ANT-118, paratype MNHN no number; Pseudoanthomastus inusitatus: Tixier-Durivault & d'Hondt, Citation1974: holotype MNHN IK-2009-2245; Pseudoanthomastus pacificus Pasternak, Citation1981: holotype IORAS IV-9-Alc-11-019; Sarcophyton purpureum Koren & Danielssen, 1883: ZMUB #509, #526.
List of abbreviations and acronyms used
IORAS, P.P. Shirshov Institute of Oceanology RAS (Moscow, Russia)
MCZ, Harvard University Museum of Comparative Zoology (Cambridge, MA, USA)
MNHN, Muséum national d'Histoire naturelle (Paris, France)
MOM, Muséum de Oceanographie de Monaco (Monaco)
NHMUK, Natural History Museum (London, UK)
OTSB, semi-balloon otter trawl
ROV, remotely operated vehicle
RRS, Royal Research Ship
RV, research vessel
TS, Sigsbee trawl
USNM, the National Museum of Natural History (Washington, DC, USA)
YPM, Yale Peabody Museum (Yale, USA)
ZMMU, the Zoological Museum of Moscow State University (Moscow, Russia)
ZMUB, Zoological Museum of Bergen University (Bergen, Norway)
ZMUC, Zoological Museum, Natural History Museum of Denmark (Copenhagen, Denmark)
Taxonomy
Class Anthozoa
Subclass Octocorallia
Order Alcyonacea
Family Alcyoniidae Lamouroux, 1812
Genus Anthomastus Verrill, Citation1878
Anthomastus Verrill, 1878: 376.
Anthomastus s. s. Bayer Citation1993: 6.
(in part) Anthomastus Studer 1901: 26–27; Kükenthal Citation1910: 57–58; Jungersen Citation1927: 5; Deichmann Citation1936: 52.
Type species: Anthomastus grandiflorus Verrill, Citation1878, by monotypy.
Diagnosis
Colonies are hemispherical to capitate with dome-shaped capitulum gradually passing via an inconspicuous stalk into the holdfast, or mushroom-shaped with a conspicuous stalk. Polyps are dimorphic and have sclerites. Autozooids are few in number, large sterile and retractile, arranged evenly over the capitulum. Sclerites are equally developed at dorsal and ventral sides of autozooids. Anthocodial armature usually more developed at the base of authozooid. Siphonozooids are fertile, numerous and densely set between autozooids, sometimes forming a continuous layer and they may be hardly visible. Distance between two neighbouring siphonozooids is about or less than the diameter of any of them. Sclerites are needles, rods, radiates, warted clubs, plates and spindles. Pharyngeal sclerites are predominately platelets. Tentacular sclerites are predominately rods, plates and crosses.
Species assigned here to the genus
Anthomastus grandiflorus Verrill, Citation1878; Sarcophyton purpureum Koren & Danielssen, 1883; A. canariensis Wright & Studer, Citation1889; Sarcophytum aberransThomson & Henderson, 1906; S. agaricoides Thomson & Henderson, 1906; A. antarcticus Kükenthal, Citation1910; A. muscarioides Kükenthal, Citation1910; A. agassizii Verrill, Citation1922; A. zealandicus Benham, 1928; A. hicksoni Bock, 1938; A. giganteus Tixier-Durivault, 1954; A. tahinodus d'Hondt, Citation1988; A. globosus d'Hondt, Citation1992; A. bathyproctus Bayer, Citation1993; A. gyratus sp. nov.
Distribution
Atlantic, Indian Ocean, Pacific, South Ocean.
Remarks
The genus Bathyalcyon Versluys, 1906 with only one autozooid and numerous siphonozoids, is considered here to be a valid genus but still defined by the diagnosis of the subgenus Anthomastus (Bathyalcyon) given by Bayer (Citation1993). In colony form with conspicuous siphonozooids three species – A. zealandicus, S. aberrans and S. agaricoides – closely resemble members of the genus Pseudoanthomastus. However, I was not able to examine the type material for A. zealandicus, which is stored at the Otago Museum (Dunedin, New Zealand) or S. aberrans and S. agaricoides, that are apparently stored in the Indian Museum (Calcutta, India). From the original description of S. aberrans it is apparent that two specimens described under the same name represent two different species. Study of the type and additional material of the mentioned species is required to clarify if they can be included in the genus Anthomastus.
Anthomastus grandiflorus Verrill, Citation1878
(A, B, )
Figure 1. A,B. Anthomastus grandiflorus (IORAS IV-9-Alc-11-025); C,D. A. gyratus sp. nov. holotype (ZMMU Ec-108); E,H. Pseudoanthomastus inusitatus holotype (MNHN IK-2009-2245), s - siphonozooids; F. P. agaricus holotype (MOM INV-6080); G. P. agaricus (IORAS IV-9-Alc-11-028); I,J. P. mariejoseae sp. nov. holotype (NHMUK 2012.46), au - autozooids. Scale A,B,E–I, 1 cm; C,D, 0.5 cm.
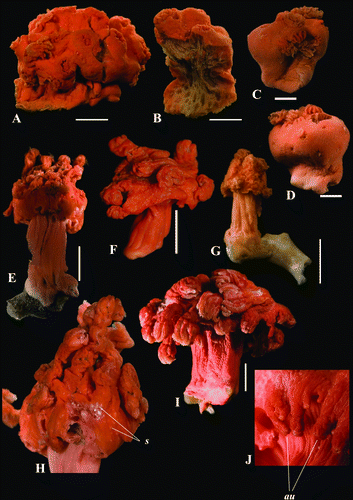
Figure 2. Anthomastus grandiflorus syntype (USNM 30181). A. Sclerites of tentacles of autozooid. B. Sclerites of anthocodia wall. Scale 0.1 mm.
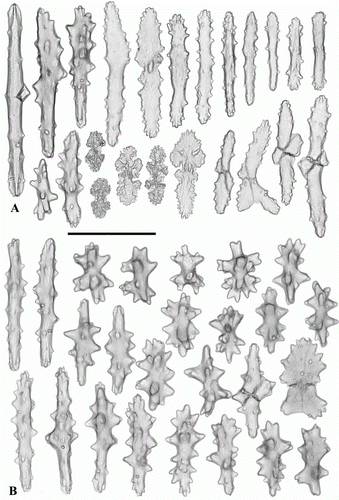
Figure 3. Anthomastus grandiflorus syntype (USNM 30181). Sclerites of capitulum. Scale 0.1 mm.
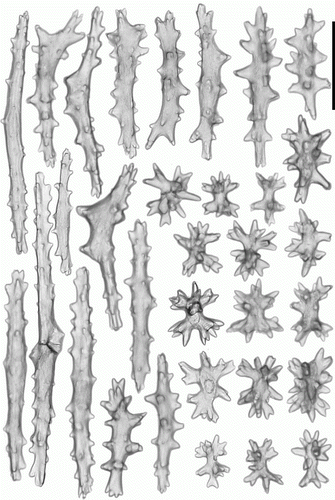
Figure 4. Anthomastus grandiflorus syntype (USNM 30181). Sclerites of stalk. Scale 0.1 mm.
Anthomastus grandiflorus Verrill, 1878: 376; Kükenthal Citation1910: 59; Bayer Citation1993: 6, plate 1.
(in part) Anthomastus grandiflorus Verrill 1883: 41–43; Verrill Citation1922: 40–42; Jungersen Citation1927: 5; Keller & Pasternak Citation2001: 556–564; van Ofwegen et al. Citation2001: 104.
(in part) Anthomastus agassizii (non Verrill, Citation1922) Deichmann Citation1936: 54 (synonymy).
non Anthomastus grandiflorus Verrill Citation1885: 513, 533; Deichmann Citation1936: 52–54; Tixier-Durivault & d'Hondt Citation1974: 1388.
Material
Syntype USNM 30181, syntype YPM 4853 off Sable Is. New Scotia; RV Akademik Mstislav Keldysh 4th cruise, St. 318-2: 2 specimens (IORAS IV-9-Alc-11-025); St. 464: 1 specimen (IORAS IV-9-Alc-11-026).
Diagnosis
Capitate colonies with spherical to dome-shaped capitulum supported by short stalk spreading proximally into holdfast. Autozooids are large and completely retractile, arranged evenly over the capitulum. Partly retracted authozooids can form spheroid knobs on the capitulum. Completely retracted polyps can be traced by star-like apertures on the even surface of the capitulum. Anthocodial armature is poorly developed in the middle part of anthocodia leaving extensive sclerite-free areas. The aboral side of the tentacular rachis is covered by the same sclerite form as found in the pharynx. Sclerites are denser in the proximal part of anthocodia and near the base of tentacles. Siphonozooids are small and form a continuous layer between autozooids.
Sclerites are rods, needles, spindles, platelets, multiradiates and crosses. Pharyngeal sclerites are platelets.
Sclerites of syntype USNM 30181 are illustrated in Figures . Tentacular sclerites (A) are mostly rods (0.08–0.22 mm long), plates (0.05–0.1 mm long) and crosses with few spindles (0.07–0.11 mm long). Anthocodial sclerites (B) are rods (0.13–0.15 mm long), spindles (0.08-0.15 mm long) crosses and multiradiates (0.05-0.08 mm long). Sclerites of the capitulum () are needles (0.27–0.3 mm long), girdled spindles (0.1–0.15 mm long), few clubs with feebly developed head (0.14–0.18 mm long), crosses and multiradiates (0.05–0.07 mm long). Sclerites of the stalk and holdfast () are spindles (0.1–0.26 mm long), multiradiates (0.04–0.07 mm long), few clubs (0.08–0.16 mm long) and crosses. Sclerome of the syntype YPM 4853 agree well with USNM 30181 by size and distribution of sclerites. However, studied material completely lack short spindles with few large tubercules arranged in three groups: in the middle part and at apices of the sclerites that were abundant in stalk and base of USNM 30181.
Distribution
Reykjanes Ridge, western Atlantic, 457–1760 m.
Remarks
The synonymy of A. grandiflorus is one of the most complicated in the genus. At least four species were considered in the literature (Jungersen Citation1927, Deichmann Citation1936, van Ofwegen et al. Citation2001) to be junior synonyms of A. grandiflorus. Specimens reported as A. grandiflorus by Verrill (Citation1883, Citation1885, Citation1922) and Deichmann (1936) represent a mixture of several species. By external appearance, A. grandiflorus can barely be distinguished from other globose species of the genus; therefore, in the distribution I included only the type locality of the species and studied material.
Anthomastus gyratus sp. nov.
(C, D, )
Figure 5. Anthomastus gyratus sp. nov. holotype (ZMMU Ec-108). A. Sclerites of tentacles of autozooid. B. Sclerites of anthocodia wall. Scale 0.1 mm.
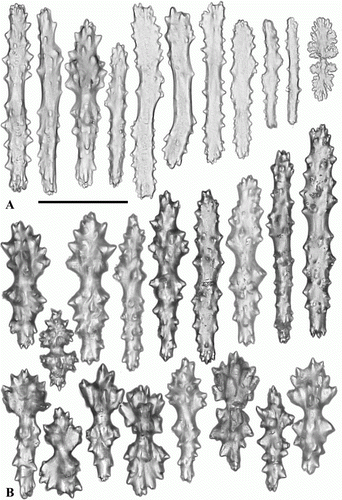
Figure 6. Anthomastus gyratus sp. nov. holotype (ZMMU Ec-108). Sclerites of capitulum. Scale 0.1 mm.
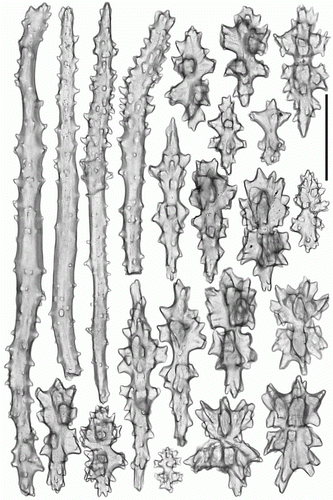
Figure 7. Anthomastus gyratus sp. nov. holotype (ZMMU Ec-108). Sclerites of stalk. Scale 0.1 mm.
(in part) Anthomastus grandiflorus Keller & Pasternak Citation2001: 556–564.
Anthomastus cf. canariensis Molodtsova et al. Citation2008: 120–121; Gebruk et al. Citation2010: 5.
? (in part) Anthomastus grandiflorus (non Verrill, Citation1878) Jungersen Citation1927: 5.
? (in part) Anthomastus agassizii (non Verrill, Citation1922) Deichmann Citation1936: 54.
Material
Holotype: RV Akademik Mstislav Keldysh 4th cruise, St. 444, 1 specimen (ZMMU Ec-108). Paratypes: RV Akademik Mstislav Keldysh 4th cruise, St. 496, 2 specimens (ZMMU Ec-109, Ec-110). Other material: RV Akademik Mstislav Keldysh 4th cruise, St. 460, 1 specimen (IORAS IV-9-Alc-11-027); RV G.O. Sars, MAR-ECO cruise, St. 70/385, 1 specimen (ZMUB 88018).
Diagnosis
Capitate colonies with spheroid capitulum supported by very short stalk spreading proximally into holdfast. Autozooids are large and completely retractile, arranged evenly over the capitulum. Completely retracted polyps can be traced by star-like apertures on even surface of the capitulum. Anthocodial armature forms crests of sclerites in proximal part of anthocodia and is uniformly developed in other parts of autozooids. Siphonozooids are small, inconspicuous from the surface and form continuous layer between autozooids. Sclerites are needles, rods, spindles, clubs, plates, multiradiates and crosses. Pharyngeal sclerites are platelets.
Description
Holotype ( C,D)
A complete, heavily contracted colony 19 mm high with a spheroid capitulum bearing 15 autozooids. Capitulum is elliptical 21×15 mm in diameter and 15 mm high (78–80% of colony); proximally capitulum contracted into very short stalk 14×7 mm in diameter and 3–4 mm high basally expending into holdfast. Autozooids arranged exclusively over distal three quarters of the capitulum. Most of autozooids are completely retracted into capitulum leaving star-like scars 0.8–1.5 mm in diameter and few polyps are fully or partly expanded. The most expanded autozooid is 3.4 mm in diameter and 8 mm long including tentacles. Tentacles are 3–4 mm long with 13–15 pairs of pinnules.
Anthocodial sclerites are more concentrated in proximal part of anthocodia and are uniformly developed in other parts of autozooids. Tentacles and pinnules with sclerites.
Siphonozooids are completely invisible from the surface, they are densely set and form a continuous layer between autozooids. Capitulum sclerites form a dense layer completely masking presence of the siphonozoids. The diameter was estimated approximately from the section as 0.3–0.4 mm.
Tentacular sclerites (A) are mostly rods (0.12–0.20 mm long) with few clubs (0.17–0.19 mm long) and plates (0.08–0.09 mm long). Anthocodial sclerites (B) are rods (0.14–0.25 mm long), clubs (0.10–0.15 mm long) and few multiradiates (0.09–0.11 mm long). Pharyngeal sclerites are platelets (0.06–0.08 mm long). Sclerites of the capitulum () are mostly needles (0.3–0.58 mm long), clubs (0.11–0.18 mm long), spindles (0.1–0.13 mm long) and multiradiates (0.05–0.14 mm long). Sclerites of the stalk and holdfast () are multiradiates (0.06–0.12 mm long), clubs (0.1–0.13 mm long), spindles (0.09–0.23 mm long) and crosses.
Paratypes
Two smaller colonies 11 and 13 mm high with a corresponding capitulum diameter 10 and 12 mm. Each colony has 5 autozooids. Paratype ZMMU EC-109 is firmly attached to basaltic stone and has a broad holdfast with maximum diameter 32 mm. Both specimens demonstrate the same colony morphology as the holotype. Siphonozooids are better visible in paratype ZMMU EC-109 with diameter 0.2–0.3 mm. Sclerome of both paratypes agree well with that of holotype.
Colour
Stalk and capitulum are puce red in ethanol, holdfast is light pink.
Etymology
Species name gyratus is from Latin ‘round’ to indicate characteristic globous form of the capitulum of the new species.
Distribution
Reykjanes Ridge, Mid-Atlantic Ridge, 1465–1800 m.
Comparisons
The new species can be distinguished from A. aberrans, A. agaricoides, A. antarcticus, A. bathyproctus, A. giganteus, A. hicksoni, A. muscarioides, A. tahinodus and A. zealandicus by the spheroidal shape of capitulum and the short stalk expanding into a broad holdfast. Also from A. agassizii, A. canariensis, A. globosus, A. grandiflorus and A. purpureus, all of which have a similar form of the colony, A. gyratus sp. nov. can be distinguished by characteristic form of the radiates and clubs (Figures ).
Genus Pseudoanthomastus Tixier-Durivault & d'Hondt, Citation1974
Pseudoanthomastus Tixier-Durivault & d'Hondt, Citation1974: 1384.
(in part) Pseudoanthomastus Pasternak Citation1981: 41–42.
(in part) Anthomastus Studer Citation1901: 26–27; Kükenthal Citation1910: 57–58; Deichmann Citation1936: 52; Tixier-Durivault & d'Hondt Citation1973; d'Hondt 1988: 265.
Type species: Pseudoanthomastus inusitatus Tixier-Durivault & d'Hondt, Citation1974 (jun. syn. of Anthomastus agaricus Studer, Citation1890), by monotypy.
Diagnosis
Mushroom-shaped, capitate colonies with dome-shaped capitulum sharply separated from a conspicuous stalk. Polyps are dimorphic and have sclerites. Autozooids are few in number, large, sterile and retractile but rarely completely retracted and arranged evenly over the capitulum. Sclerites are equally developed round autozooids. Anthocodial armature more developed near the base of the tentacles. Siphonozooids are fertile, usually conspicuous, feebly armoured, not numerous and are scattered among authozooids. Sclerites are radiates, warted clubs, clubs with head formed of distally pointed tubercules, rods, spindles and crosses. Pharyngeal sclerites are predominately rodlets. Tentacular sclerites are needles, rods, clubs and radiates.
Species assigned here to the genus
Anthomastus agaricus Studer, Citation1890; A. fisheri Bayer, 1952; A. venustus Tixier-Durivault & d'Hondt, Citation1973; P. mariejoseae sp. nov.
Distribution
North Atlantic, Indian Ocean, Central Pacific, South Pacific.
Remarks
The genus Pseudoanthomastus Tixier-Durivault & d'Hondt, Citation1974 was initially described to accommodate P. inusitatus Tixier-Durivault & d'Hondt, Citation1974, characterized by only one type of polyp. Later, d'Hondt (1988) relegated Pseudoanthomastus to a junior synonym of the genus Anthomastus. Indeed, closer investigation of the type material revealed polyps of two types (C). Nevertheless, due to well-defined colony morphology and differences in the sclerome (predominant rodlets in pharyngeal sclerites and highly diverse tentacular sclerites), I propose here to retain the generic name Pseudoanthomastus. At the same time, Pseudoanthomastus pacificus Pasternak, Citation1981 with monomorphic polyps cannot be considered as a member of the genus and has to be assigned to the genus Alcyonium Linnaeus, 1758. The name Alcyonium pacificum is already occupied (Yamada Citation1950), therefore I propose here A. pasternaki sp. nov. as a replacement name for P. pacificus Pasternak, Citation1981.
Associations with the polynoids reported for Anthomastus grandiflorus (Ditlevsen Citation1917; Martin & Britayev Citation1998; Watling et al. Citation2011) are in fact documented exclusively for members of the genus Pseudoanthomastus.
Pseudoanthomastus agaricus (Studer, Citation1890)
(E–H, )
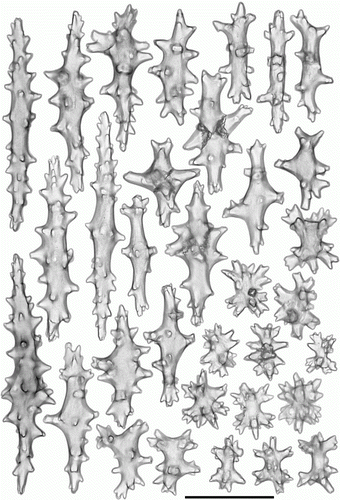
Figure 8. Pseudoanthomastus agaricus holotype (MOM INV-6080). Sclerites of tentacles of autozooids. Scale 0.1 mm.
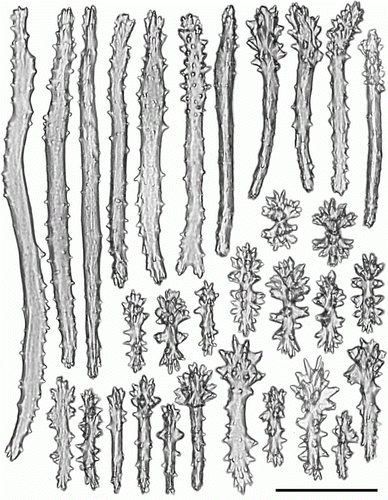
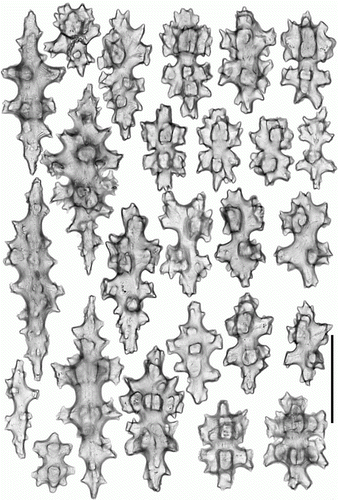
Figure 9. Pseudoanthomastus agaricus holotype (MOM INV-6080). Sclerites of anthocodia wall. Scale 0.1 mm.
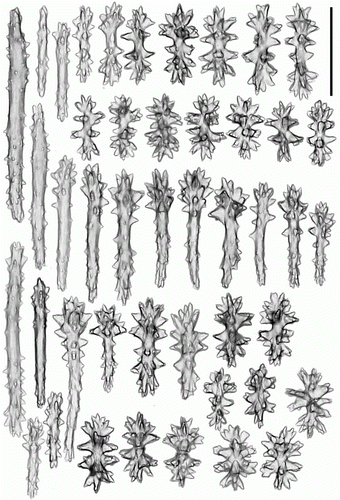
Figure 10. Pseudoanthomastus agaricus holotype (MOM INV-6080). Sclerites of capitulum. Scale 0.1 mm.

Figure 11. Pseudoanthomastus agaricus holotype (MOM INV-6080). Sclerites of stalk. Scale 0.1 mm.
(in part) Anthomastus agaricus Studer, Citation1890: 88; 1901: 27.
Anthomastus agaricus Kükenthal Citation1910: 6; Thomson Citation1927: 12; Keller & Pasternak Citation2001: 557.
Anthomastus canariensis (non Wright & Studer, 1878) Tixier-Durivault & d'Hondt Citation1974: 1388.
(in part) Jungersen Citation1927: 5; Deichmann Citation1936: 52 (synonymy); Keller & Pasternak Citation2001: 556–564.
(in part) Anthomastus agassizii (non Verrill, Citation1922) Deichmann Citation1936: 54 (synonymy).
Pseudoanthomastus inusitatus Tixier-Durivault & d'Hondt, Citation1974: 1384–1388.
non Anthomastus agaricus Tixier-Durivault & d'Hondt Citation1974: 1388; Molodtsova et al. Citation2008: 119–121; Gebruk et al. Citation2010: 5.
Material
A. agaricus holotype: MOM INV-6080 (120016); P. inusitatus holotype: MNHN IK-2009-2245; RV Akademik Mstislav Keldysh 4th cruise, St 460, 4 specimens (IORAS IV-9-Alc-11-028); St 464, 1 specimen (IORAS IV-9-Alc-11-029).
Diagnosis
Mushroom-shaped colonies with rather flattened capitulum separated from conspicuous longitudinally striated cylindrical stalk by distinct grove. The rim of the capitulum is uneven with few polyps at the margin. Authozooids retractile but rarely completely retracted. Sclerites are more densely set near the base of the tentacles and continue as ridges up the aboral side of the tentacular rachis. Pinnules (12–13 pairs) with sclerites. Siphonozooids are visible as small whitish tubercules (H). Distance between two neighbouring siphonozooids is more than one diameter of a siphonozooid. Anthocodial wall and tentacles of authozooids with multiradiates, clubs, rods and spindles; pharyngeal wall with rodlets. Outmost layer of capitulum has a similar set of sclerites as the anthocodial wall; deeper layers of the capitulum and stalk have spindles and rods. Stalk surface has rods, multiradiates and clubs.
Sclerites of holotype MOM INV-6080 are illustrated in Figures . Tentacular sclerites () are with needles (0.3–0.43 mm long), spindles (0.2–0.3 mm long), multiradiates (0.06–0.07 mm long), clubs (0.09–0.22 mm long) and rods (0.07–0.1 mm long). Anthocodial wall () with girdled spindles (0.07–0.11 mm long), multiradiates (0.07–0.08 mm long), clubs (0.1–0.18 mm long) and rods (0.09–0.25 mm long); pharyngeal wall with rodlets (0.07–0.11 mm long). Capitulum () has needles (0.3–0.4 mm long), girdled spindles (0.08–0.11 mm long), multiradiates (0.05–0.08 mm long), clubs (0.08–0.18 mm long) and rods (0.05–0.16 mm long). Stalk () has needles (0.3–0.6 mm long), spindles (0.18–0.22 mm long), multiradiates (0.06–0.08 mm long), clubs (0.08–0.14 mm long) and crosses.
Distribution
Reykjanes Ridge, Mid-Atlantic Ridge, off Newfoundland: 665–2567 m.
Remarks
The younger specimen of Anthomastus agaricus reported by Studer (1901: 27, plate 1, figure 7a–c) from the same station as the holotype belongs to another genus and species, Heteropolypus sol sp. nov., described below. The specimens reported by Molodtsova et al. (Citation2008) and Gebruk et al. (Citation2010) as A. agaricus belong to Pseudoanthmastus mariejoseae sp. nov., described here below.
Pseudoanthomastus mariejoseae sp. nov.
(I–J, )
Figure 12. Pseudoanthomastus mariejoseae sp. nov. holotype (NHMUK 2012.46). Sclerites of tentacles of autozooids. Scale 0.1 mm.

Figure 13. Pseudoanthomastus mariejoseae sp. nov. holotype (NHMUK 2012.46). Sclerites of anthocodia wall. Scale 0.1 mm.

Figure 14. Pseudoanthomastus mariejoseae sp. nov. holotype (NHMUK 2012.46). Sclerites of capitulum. Scale 0.1 mm.

Figure 15. Pseudoanthomastus mariejoseae sp. nov. holotype (NHMUK 2012.46). Sclerites of stalk. Scale 0.1 mm.

Anthomastus agaricus Molodtsova et al. Citation2008: 119–121; Gebruk et al. Citation2010: 5.
(in part) Anthomastus canariensis (non Wright & Studer, 1878) Tixier-Durivault & d'Hondt Citation1974: 1388.
Material
Holotype: RRS James Cook JC048/016 (NHMUK 2012.46). Paratype: RV G.O. Sars MAR-ECO cruise, St. 72/386, 1 specimen (ZMUB 88017). Other material: NO Jean Charcot Biaçores DR 166, 3 specimens (MNHN IK-2009-2247).
Diagnosis
Mushroom-shaped colonies with prominent capitulum sharply separated from conspicuous cylindrical stalk by distinct grove. Autozooids are large and retractile, arranged evenly over the capitulum. Sclerites are equally developed on all sides of autozooids. Anthocodial armature forms crests of sclerites in basal part of tentacles. Siphonozooids are fertile, small, and clearly visible from the surface with up to 10 rows of siphonozooids between adjacent autozooids. Sclerites are multiradiates, clubs, rods, plates, needles, spindles and crosses. Pharyngeal sclerites are predominately rodlets. Tentacular sclerites are rods, girdled spindles, radiates and crosses.
Description
Holotype ( E)
A complete colony 48 mm high with spheroid capitulum bearing 30 large autozooids evenly arranged over the capitulum and about 33 smaller ones arranged at the margin of the capitulum (F). The capitulum is 50 mm in diameter and 15 mm high (30% of colony); proximally capitulum separated from conspicuous cylindrical stalk by distinct grove inhabited by scale worm. Most of autozooids are expanded or partly retracted and only one autozooid is completely retracted leaving low lump on the capitulum. Fully expanded autozooids are also arranged at lumps. The most expanded autozooid is 7 mm in diameter and 22 mm long including tentacles. Tentacles are 8–10 mm long with 13–15 pairs of pinnules.
Siphonozooids are clearly visible from the surface and do not form a continuous layer, few rows of siphonozoids are clearly visible between neighbouring ones. The diameter of siphonozooids was estimated as 0.4–0.45 mm.
Tentacular sclerites () are rods (0.14–0.46 mm long), girdled spindles (0.1–0.17 mm long), rodlets (0.08–0.13 mm long) and crosses. Anthocodial sclerites () are rods (0.12–0.22 mm long), girdled spindles (0.1–0.15 mm long), clubs (0.12–0.19 mm long) and crosses. Pharyngeal sclerites are rodlets (0.1–0.12 mm long). Sclerites of the capitulum () are needles (0.3–0.43 mm long), rods (0.16–0.45 mm long), girdled spindles (0.1–0.22 mm long), sometimes with asymmetrically arranged tubercules, clubs (0.11–0.19 mm long) and multiradiates (0.06–0.09 mm long). Sclerites of the stalk and holdfast () are rods (0.2–0.40 mm long), girdled spindles (0.09–0.14 mm long), multiradiates (0.07–0.09 mm long) and crosses.
Paratype
A small mushroom-shaped colony with 10 autozooids and clearly visible siphonozooids. Size and distribution of sclerites agrees with that of the holotype. The specimen was initially preserved in the formalin and in one month transferred into alcohol so some minor difference in the form of sclerites may be due to the mode of preservation.
Colour
Stalk and capitulum are bright red in the ethanol, stalk is reddish-pink.
Etymology
Species name mariejoseae is in honour to late Marie-Jose d'Hondt, a researcher from MNHN who spent a great part of her career studying the genus Anthomastus.
Distribution
Reykjanes Ridge, Mid-Atlantic Ridge, Azores 130–1800 m.
Comparisons
The new species closely resembles other species of Pseudoanthomastus in the characteristic form of the colony, but in comparison to other species of the genus has a more stout appearance. P. mariejoseae sp. nov. can be distinguished from P agaricus (Studer, Citation1890), P. fisheri (Bayer, 1952) and P. venustus (Tixier-Durivault & d'Hondt, Citation1973) by the sclerome, namely by the presence of characteristic rods with high and densely set tubercules in all regions of the colony and by relatively low number of clubs in autozooids ( and ). A dense row of small autozooids around the rim of the capitulum present in all studied colonies of P. mariejoseae sp. nov. and not reported for other species of the genus can be due to the presence of symbiotic polynoid worms.
Genus Heteropolypus Tixier-Durivault, Citation1964
Heteropolypus Tixier-Durivault, Citation1964: 67–68.
Type species: Heteropolypus insolitus Tixier-Durivault, Citation1964, by monotypy.
Diagnosis
Colonies are mushroom-shaped to obconic, distinctly separated into capitulum and sterile stalk. Polyps are dimorphic or trimorphic and have sclerites. Autozooids are few in number, large sterile and retractile, arranged evenly over the capitulum or only at the margin. Anthocodial armature is often asymmetrically developed with ridges of sclerites more developed at dorsal side of authozooid. Siphonozooids are fertile, numerous, strongly armored, and densely set between autozooids. Mesozooids when present possess feebly developed tentacles and are retractile and scattered among siphonozooids. Sclerites are flanged rods, radiates, clubs, rods, plates and spindles. Pharyngeal sclerites are predominately platelets. Tentacular sclerites are rods, clubs, spindles, multiradiates, plates and crosses.
Species assigned here to the genus
Heteropolypus insolitus Tixier-Durivault, Citation1964; A. ritteri Nutting, 1909; H. sol sp. nov.; A. japonicusNutting, 1906; A. rylovi Naumov, 1952; A. steenstrupi Wright & Studer, Citation1889.
Distribution
North Atlantic, Indian Ocean, Pacific.
Remarks
In the original description of Heteropolypus (Tixier-Durivault Citation1964: 56–57) the presence of mesozooids was considered to be the main characteristic of the genus. However, I doubt that mesozooids can be considered as characteristic of generic level. Mesozooids were developed in other groups of Octocorallia. Thus, at least 3 of the 35 nominal species of the genus Pennatula (Pennatulacea: Pennatulidae) posess mesozooids (Kükenthal Citation1915; Williams Citation2011). In early developmental stages (less than 4 fully developed autozooids) and in badly preserved colonies I was not able to see the mesozooids of H. sol sp. nov. Probably mesozooids appear when the colony attains a certain size or in particular time of the life cycle.
Heteropolypus sol sp. nov.
(Figures )
Figure 16. Heteropolypus sol sp. nov. A,B,D–H, general view of colonies; C, side view of autozooid, small arrowheads indicate a ridge of sclerites at dorsolateral side of autozooid, large arrowheads indicate ventral side of autozoid free of ridges; I, fragment of capitulum with siphonozooids (s) and mesozooids (m). A–C,I, Holotype (ZMMU Ec-111); D, colony with one autozooid, paratype (ZMMU Ec-112/1); E,G, colonies with two autozooids, paratype (ZMMU Ec-112/2, Ec-112/3); F, colony with three autozooids, paratype (ZMMU Ec-112/4); H, colony with completely retracted autozooids, paratype (ZMMU Ec-112/5). Scale A,B,D–H, 1 cm; C, 0.5 cm; I, 2 mm.
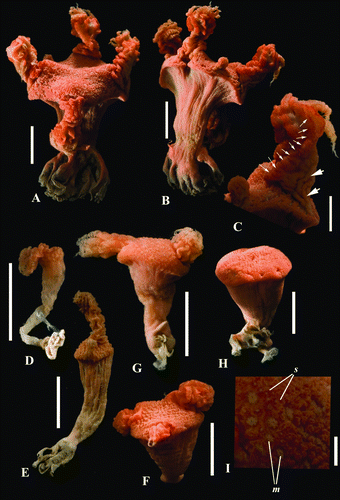
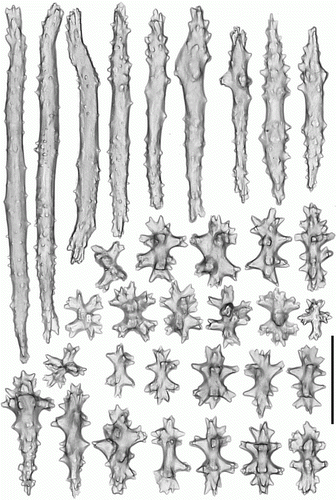
Figure 17. Heteropolypus sol sp. nov. holotype (ZMMU Ec-111). Sclerites of tentacles of autozooids. Scale 0.1 mm.

Figure 18. Heteropolypus sol sp. nov. holotype (ZMMU Ec-111). Sclerites of anthocodia wall. Scale 0.1 mm.

Figure 19. Heteropolypus sol sp. nov. holotype (ZMMU Ec-111). Sclerites of capitulum. Scale 0.1 mm.
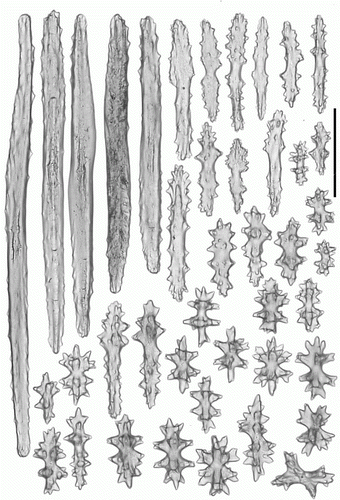
Figure 20. Heteropolypus sol sp. nov. holotype (ZMMU Ec-111). Sclerites of stalk. Scale 0.1 mm.

Heteropolypus cf. insolitus Molodtsova et al. Citation2008: 121.
Anthomastus grandiflorus (non Verrill, Citation1878) Verrill 1885: 513, 533, plate 2 (12); Deichmann Citation1936: 52–54; Tixier-Durivault & d'Hondt Citation1974: 1388.
(in part) Anthomastus grandiflorus Verrill 1883: 41–43, plate 1 (6–10); Verrill Citation1922: 40–42, plate 14 (7, 8); Jungersen Citation1927: 5; van Ofwegen et al. Citation2001: 104.
(in part) Anthomastus agaricus Studer 1890: 88; 1901: 27.
Material
Holotype: RV Akademik Mstislav Keldysh 4th cruise, St. 316-5 (ZMMU Ec-111). Paratypes: same station data as holotype: 56 specimens (ZMMU Ec-112). Other material: RV Akademik Mstislav Keldysh 4th cruise, St. 317, 14 specimens (IORAS IV-9-Alc-11-030); St 500, 1 specimen (IORAS IV-9-Alc-11-031); RV G.O. Sars, MAR-ECO cruise, St. 50/373, 1 specimen (ZMUB 88019); RRS Challenger 14A/1978, St 50304, 1 specimen (NOCS no number); RRS James Cook JC011/017 (NOCS no number); JC011/023B (NOCS no number); JC037/015 (NOCS no number); JC037/019 (NOCS no number); JC037/027 OTSB (NOCS no number); JC048/054 (NOCS no number); NO Jean Charcot, Biaçores, BI77, 6 specimens (IK-2009-2247).
Diagnosis
Colonies are obconic to mushroom-shaped, distinctly separated into convex or almost flat capitulum, polygonal from top view, with a longitudinally striated stalk. The basal part of the stalk is modified into rhizoids. Autozooids are few in number, arranged round the margin of the capitulum. Anthocodial armature is asymmetrically developed with ridges of sclerites more strongly developed at dorsal side (C). Siphonozooids are densely set between autozooids and armored with needles. There are at least 5 rows of siphonozooids between margin and autozooid. Mesozooids, possessing feebly developed tentacles with 2–3 pairs of pinnules, are retractile and scattered among siphonozooids. Sclerites are multiradiates, clubs, rods, platelets, spindles, girdled spindles and crosses. Pharyngeal sclerites are predominately platelets.
Description
Holotype ( A–C, I)
A mushroom-like colony about 36 mm high: capitulum 11 mm and stalk 25 mm. The capitulum is flattened from the top and elliptical in the crosssection with larger diameter of 26 mm. When viewing from the top capitulum has polygonal appearance with angles corresponding to the six autozooids arranged at the margin. Autozooids range from 1.5–3 to 6–9 mm in diameter and can be up to 15–16 mm long. Tentacles of autozooids 6–7 mm long with 13–15 pairs of pinnules. Sclerites form prominent crests on the aboral side of each tentacle that terminate near the base of three ventral tentacles but continue to anthocodia as ridges by 5 tentacles arranged dorsally with more developed ridges corresponding to 3 dorsal tentacles (C). Siphonozooids are 0.3–0.45 mm in diameter, without tentacles, highly armoured and densely crowded in such a way that the distance between two neighbouring siphonozooids is less than half of their diameter. Mesozooids (I) are 0.6–0.9 mm in diameter with short, feebly developed tentacles 0.2–0.3 mm long with 1–2 pairs of pinnules, sparsely arranged all around the capitulum with total number ~150.
Tentacular sclerites () are mainly flanged rods (0.31–0.38 mm long), girdled spindles (0.7–0.11 mm long), rods (0.06–0.21 mm long), clubs (0.12–0.14 mm long), plates (0.07–0.08 mm long), radiates (0.06–0.07 mm long) and crosses. Anthocodial sclerites () are flanged rods (0.25–0.38 mm long), girdled spindles (0.8–0.16 mm long), rods (0.2–0.26 mm long), clubs (0.08–0.16 mm long), plates (0.07–0.12 mm long) and radiates (0.06–0.08 mm long). Pharyngeal sclerites are platelets (0.04–0.14 mm long). Sclerites of the capitulum () surface are mostly multiradiates (0.06–0.08 mm long), clubs (0.12–0.4 mm long), rods (0.15–0.3 mm long), spindles (0.06–0.1 mm long) and crosses. Sclerites of the capitulum interior are predominately flanged rods (0.3–0.7 mm long). Sclerites from the stalk and rhizoids () are rods (0.16–0.4 mm long), clubs (0.11–0.13 mm long), spindles (0.05–0.12 mm long), radiates (0.07–0.08 mm long) and crosses.
Paratypes
Fifty-six specimens from the same station as the holotype were included in the type series. The specimens have colony form varying from conical to mushroom-shaped with an elliptical capitulum bearing from 1 (D) to 9 autozooids and having a diameter of 3.0×3.5 mm to 31×34 mm. Only 4 colonies out of 56 studied have one autozooid developed in the central part of the capitulum. In all other colonies autozooids were arranged exclusively along the margin. Developed mesozooids were generally found in colonies having more than 4 fully developed autozooids and are totally absent in colonies with up to 3 autozooids. Contracted mesozooids are barely visible from the surface.
Other material
All colonies agree with the type series.
Colour
Capitulum is red in ethanol, stem is greyish red with grey rhizoids.
Etymology
Species name sol is from Latin ‘sun’ to indicate the characteristic form of the colony of the new species, resembling a stylized image of the sun owing to expended autozooids arranged along the margin.
Distribution
Reykjanes Ridge, Mid-Atlantic Ridge, Nova Scotia, Rockall Trough, Azores, Porcupine Abyssal plain, 2132–4298 m.
Comparisons
Heteropolypus sol sp. nov. is close to H. insolitus but differs clearly from H. ritteri, H. japonicus and H. rylovi by having autozooids arranged mostly along the margin of the capitulum and by the presence of rhizoids. H. sol can be distinguished from H. insolitus by having only few developed autozooids (up to 9), polygonal form of the capitulum, more rows of siphonozoids between autozooids and the margin of capitulum (7–10 vs. 1–2) and also characteristic sclerome (Figures ). H. steenstrupi was described based on a small incomplete colony with polygonal capitulum having 6 autozooids around margin, without mesozooids and lacking the basal part. H. sol can be distinguished from H. steenstrupi by the fewer number of siphonozoids between autozooids and margin, clearly recognizable margin of the capitulum and also by its characteristic sclerome.
Apart from the four species considered in the present article, three other species of Anthomastus were described or reported earlier from the North Atlantic: A. purpureus (Koren & Danielssen, Citation1883), A. canariensis Wright & Studer, Citation1889 and A. agassiziiVerrill, Citation1922. In a number of reports and regional faunal lists (Jungersen Citation1927; Deitchmann 1939; van Ofwegen et al. 2001), A. purpureus and A. canariensis were considered as junior synomyms of A. grandiflorus Verrill. However, as correctly shown by d'Hondt (Citation1992) who also provided a brief redescription and illustration of sclerites of A. canariensis, each of these two species has a very characteristic set of sclerites different from that in A. grandiflorus. Therefore, these two species can be considered as valid.
Anthomastus agassizii was described in only a few lines in the paper dealing with material from the Canadian Arctic Expedition (Verrill Citation1922: 41), but the fact that the type specimen had been collected off Dominica (Lesser Antilles, the Caribbean Sea) () was overlooked by a number of researchers. The species was later reported from several areas including Georges Bank and other banks off Gloucester, Massachusetts (Deichmann Citation1936), Bear seamount (Moore et al. Citation2003) and in a recent review of deep-sea octocorals (Watling et al. Citation2011) it is considered in the North West Atlantic fauna, IIA sensu Cairns & Chapman (Citation2001). It is quite possible that due to misinterpretation of A. grandiflorus in later papers of Verrill (Citation1883, Citation1885, Citation1922) and Deichmann (1936), specimens reported as A. agassizii in fact belong to A. grandiflorus and A. gyratus sp. nov. The holotype of A. agassizii has dried out and redescription of this species based on freshly collected specimens is required.
Further analyses of available published data will help the understanding of patterns of distribution of particular species of di- and trimorphic alcyoniids in the North Atlantic.
Editorial responsibility: Ole S. Tendal
Acknowledgements
I would have had no progress and no results without the help of technicians and curators of Natural History museums, colleagues from various research institutes around the world who donated or loaned me material and helped with literature and advices, also crews of scientific vessels always welcomed and helped me, and good friends of mine who stayed close when I really needed them. In no particular order: S. Cairns, P. Greenhall, A. Collins (Smithsonian Institute), D. Tracey, K. Schnider, S. Mills (NIWA), E. Laso-Wasem (Peabody Museum), A. Baldinger (MCZ), G. Paterson, A. Cabrinovic (NHMUK), G. Williams (Californian Academy of Sciences), D. Fautin (KU), B. Richer de Forges (retired) A. Croisnier (retired), Ph. Bouchet, Ph. Maestrati, V. Heros, P. Lozouet, A. Andouche, N. Amesiane (MNHN, Paris), L. van Ofwegen (Naturalis, Leiden), E. Willassen, J. Kongsrud (Bergen Museum), E. Le Guilloux, J. Davies, B. Guillaumont, I. van den Beld (Ifremer Brest), E. Lazo-Wasem (YPM), V. Warenham (Northwest Atlantic Fisheries Centre), F.G. Hochberg (Santa Barbara Museum of Nat. History), J. Murillo Pérez (IEO), M. Bruni (Musée océanographique de Monaco), D. Billet, A. Gooday, C. Alt (NOCS), A. Mironov, A. Gebruk, E. Krylova, A. Rogacheva, N. Budaeva (IORAS), A. Tzetlin (WSBS, MSU). Special thanks to Professor Imants G. Priede for advice and editing the style of English and to Phil Alderslade (CSIRO) for his invaluable oral comments on the manuscript and help with sclerite imaging and terminology. I thank the UK NERC-funded ECOMAR consortium (NE/C512961/1) for access to the RRS James Cook samples from the Mid-Atlantic Ridge. Also, I am grateful for financial support from the Norwegian Ministry of Foreign Affairs granted to the Institute of Marine Research, Norway, RFBR research project 12-05-33049, and the MAR-ECO project.
Notes
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
Related Research Data
References
- Alderslade , P. 1994 . A redescription of Anthomastus agilis Tixier–Durivault with generic placement in Sinularia (Coelenterata: Octocorallia), and some remarks on Sinularia ramosa Tixier- Durivault . The Beagle , 11 : 133 – 39 .
- Aldred RG , Thurston MH , Rice AL , Morley DR. 1976 . An acoustically monitored opening and closing epibenthic sledge . Deep Sea Research and Oceanographic Abstracts 23 : 167 – 174 , IN1–IN6 . doi: 10.1016/S0011-7471(76)80025-3
- Bayer , FM. 1993 . Taxonomic status of the octocoral genus Bathyalcyon (Alcyoniidae: Anthomastinae), with descriptions of a new subspecies from the Gulf of Mexico and a new species of Anthomastus from Antarctic waters . Precious Corals and Octocoral Research , 1 : 3 – 13 .
- Bayer , FM , Grasshoff , M and Verseveldt , J. 1983 . Illustrated Trilingual Glossary of Morphological and Anatomical Terms Applied to Octocorallia , 1 – 75 . Leiden : EJ Brill .
- Cairns , SD and Chapman , RE. 2001 . “ Biogeographic affinities of the North Atlantic deep-water Scleractinia ” . In Proceedings of the First International Symposium on Deep-Sea Corals , Edited by: Willison , JHM , Hall , J , Gass , SE , Kenchington , ELR , Butler , M and Doherty , P . 30 – 57 . Halifax : Ecology Action Centre .
- Deichmann , E. 1936 . The Alcyonaria of the western part of the Atlantic Ocean. Memoires of the Museum of Comparative Zoology . Harvard , 53 : 1 – 317 .
- Ditlevsen , H. 1917 . Annelids. I. Danish Ingolf-Expedition . Copenhagen. , 4 ( 4 ) : 1 – 71 .
- Fabricius , KE and Alderslade , P. 2001 . Soft Corals and Sea Fans: A Comprehensive Guide to the Tropical Shallow Water Genera of the Central-West Pacific, the Indian Ocean and the Red Sea , Townsville : Australian Institute of Marine Science. 264 pages .
- Gebruk , AV , Budaeva , NE and King , NJ. 2010 . Bathyal benthic fauna of the Mid-Atlantic Ridge between the Azores and the Reykjanes Ridge . Journal of the Marine Biological Association of the United Kingdom , 90 : 1 – 14 . doi: 10.1017/S0025315409991111
- d'Hondt M-J. 1988 . Anthomastus tahinodus n. sp., octocoralliaire Alcyonacea du nord-est de Tahiti . Bulletin du Muséum National d'Histoire Naturelle, Paris (4) 10(A) : 256 – 76 .
- d'Hondt M-J. 1992 . Description d’Anthomastus globosus n. sp. (Octocorallia, Alcyonacea) de Nouvelle-Caledonie. Remarques sur quelques especes du genre . Bulletin du Muséum National d'Histoire Naturelle, Paris (4) 14(A) : 623 – 38 .
- Hickson , SJ. 1904 . The Alcyonaria of the Cape of Good Hope. Part II . Marine Investigations in South Africa , 3 : 211 – 39 .
- Jungersen , HFE. 1927 . Anthomastus. Danish Ingolf-Expedition . Copenhagen , 5 ( 11 ) : 1 – 14 .
- Keller , NB and Pasternak , FA. 2001 . Corals (Scleractinia, Alcyonacea, Gorgonacea, Pennatulacea) and their role in the seascapes formations of the rift zone in Reykjanes Ridge . Oceanology , 41 : 556 – 64 .
- Kidd , RB , Huggett , QJ and Ramsay , ATS. 1990 . The status of geological dredging techniques . Marine Geophysical Research , 12 : 131 – 43 . doi: 10.1007/BF00310568
- Kükenthal , W. 1910 . Zur Kenntnis der Gattung Anthomastus Verr. In: Doflein F, editor. Beitrage zur Naturgeschichte Ostasiens. Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften, Mathematisch-Physikalische Klasse . Suppl. , 1 ( 9 ) : 1 – 16 .
- Kükenthal W. 1915 . Pennatularia. Das Tierreich . 43 . Berlin : Verlag von R. Friedländer und Sohn . 132 pages.
- Koren J , Danielssen DC. 1883 . Nye Alcyoniider, Gorgonider og Pennatulider tilhorende Norges Fauna. Bergens Museum . Bergen : John Griegs Bogtrykkeri , p i–ii + I–XVI + 1–38 .
- Madsen , FJ. 1944 . Octocorallia. Danish Ingolf-Expedition . Copenhagen , 5 ( 13 ) : 1 – 65 .
- Martin , D and Britayev , TA. 1998 . Symbiotic Polychaetes: Review of known species . Oceanography and Marine Biology: An Annual Review , 36 : 217 – 340 .
- Molodtsova , TN , Sanamyan , NP and Keller , NB. 2008 . Anthozoa from the northern Mid-Atlantic Ridge and Charlie-Gibbs Fracture Zone . Marine Biology Research , 4 : 112 – 30 . doi: 10.1080/17451000701821744
- Moore , JA , Vecchione , M , Hartel , KE , Collette , BB , Galbraith , JK Gibbons , R . 2003 . Biodiversity of Bear Seamount, New England seamount chain: Results of exploratory trawling . Journal of Northwest Atlantic Fishery Science , 31 : 363 – 72 .
- Ofwegen L van , Grasshoff M , Land J van der . 2001 . Octocorallia (excl. Pennatulacea) . In : Costello MJ , Emblow CS , White R European Register of Marine Species: A Check-List of the Marine Species in Europe and a Bibliography of Guides to their Identification . Collection Patrimoines Naturels 50 : 104 – 105 .
- Pasternak , FA. 1981 . “ Alcyonacea and Gorgonacea ” . In Benthos of Submarine Mountains Marcus-Necker and Adjacent Pacific Regions , Edited by: Kuznetsov , AP and Mironov , AN . 40 – 55 . Moscow : P.P. Shirshov Institute of Oceanology, Academy of Sciences of the USSR .
- Priede IG , Billett DSM , Brierley AS , Hoelzel R , Inall M , Miller PI. 2013 . The ECO-MAR (Ecosystem of the Mid-Atlantic Ridge at the Sub-Polar Front and Charlie Gibbs Fracture Zone) project: description of the benthic sampling programme 2007–2010 . Marine Biology Research 9 : 624 – 28 .
- Studer , T. 1890 . Note preliminaire sur les alcyonaires provenant des campagnes du yacht ‘l'Hirondelle’ 1886–1887–1888 . Memoires de la Société zoologique de France , 3 ( 5 ) : 551 – 59 .
- Studer , T. 1901 . Alcyonaires provenant des campagnes de l'Hirondelle. Resultats des Campagnes Scientifiques . Monaco , 20 : 1 – 64 .
- Thomson , JA. 1927 . Alcyonaires provenant des campagnes scientifiques du Prince Albert Ier de Monaco. Resultats des Campagnes Scientifiques . Monaco , 73 : 1 – 77 .
- Tixier-Durivault , A. 1964 . Stolonifera et Alcyonacea . Galathea Report , 7 : 43 – 58 .
- Tixier-Durivault , A and d'Hondt , M-J. 1973 . Nouvelles récoltes d'Octocoralliaires à Madagascar . Tethys , 5 : 251 – 66 .
- Tixier-Durivault A , d'Hondt M-J . 1974 . Les octocoralliaires de la campagne Biaçores . Bulletin du Museum National d'Histoire Naturelle , Paris (3) 252 : 1361 – 433 .
- Utinomi , H and Imahara , Y. 1976 . A new second species of dimorphic alcyonacean octocoral Minabea from the Bays of Sagami and Suruga, with the emendation of generic diagnosis. Publications of the Seto Marine Biological Laboratory . Shirahama , 23 : 205 – 12 .
- Verrill AE . 1878 . Notice of recent additions to the marine fauna of the eastern coast of North America. 2 . American Journal of Science and Arts ( 3 ) 16 : 371 – 78 .
- Verrill , AE. 1883 . Report on the Anthozoa, and on some additional species dredged by the ‘Blake’ in 1877–1879, and by the U.S. Fish Commission steamer ‘Fish Hawk’ in 1880–82 . Bulletin of the Museum of Comparative Zoology, Harvard , 11 : 1 – 72 .
- Verrill , AE. 1885 . Results of the explorations made by the steamer ‘Albatross,’ off the northern coast of the United States, in 1883 . United States Commission of Fish and Fisheries Annual Report for , 1883 : 503 – 699 .
- Verrill AE . 1922 . Alcyonaria and Actinaria . Report of the Canadian Arctic Expedition 1913–1918 8(G) : 1 – 164 .
- Watling , L and Auster , PJ. 2005 . “ Distribution of deep-water Alcyonacea off the northeast coast of the United States ” . In Cold-Water Corals and Ecosystems , Edited by: Freiwald , A and Roberts , JM . 279 – 96 . Berlin : Springer-Verlag .
- Watling , L , France , SC , Pante , E and Simpson , A. 2011 . Biology of deep-water octocorals . Advances in Marine Biology , 60 : 41 – 122 .
- Wenneck , T de L , Falkenhaug , T and Bergstad , OA. 2008 . Strategies, methods, and technologies adopted on the R.V. G.O. Sars MAR-ECO expedition to the Mid-Atlantic Ridge in 2004 . Deep-Sea Research II , 55 : 6 – 28 . doi: 10.1016/j.dsr2.2007.09.017
- Williams , GC. 1990 . A new genus of dimorphic soft coral from the southwestern fringe of the Indo-Pacific (Octocorallia: Alcyoniidae) . Journal of Zoology , 221 : 21 – 35 . doi: 10.1111/j.1469-7998.1990.tb03772.x
- Williams , GC. 1992 . The Alcyonacea of southern Africa. Stoloniferous Octocorals and soft corals (Coelenterata, Anthozoa) . Annals of the South African Museum , 100 : 249 – 358 .
- Williams , GC. 2011 . The global diversity of sea pens (Cnidaria: Anthozoa: Octocorallia) . PLoS ONE , 6 ( 7 ) : 1 – 11 . doi: 10.1371/journal.pone.0022747
- Wright EP , Studer T . 1889 . Report on the Alcyonaria collected by H.M.S . Challenger during the years 1873–1876 . Reports of the Scientific Results of the Voyage of HMS Challenger, Zool . 31 : i–lxxvii + 1–314 .
- Yamada , M. 1950 . Descriptions of two Alcyonium from Northern Japan . Annotationes Zoologicae Japonensis , 23 : 114 – 16 .