Abstract
In situ and on-board pulse-chase experiments were carried out on a sublittoral fine sand in the German Bight (southern North Sea) to investigate the hypothesis that sandy sediments are highly active and have fast turnover rates. To test this hypothesis, we conducted a series of experiments where we investigated the pathway of settling particulate organic carbon through the benthic food web. The diatom Ditylum brightwellii was labelled with the stable carbon isotope 13C and injected into incubation chambers. On-board incubations lasted 12, 30 and 132 h, while the in situ experiment was incubated for 32 h. The study revealed a stepwise short-term processing of a phytoplankton bloom settling on a sandy sediment. After the 12 h incubation, the largest fraction of recovered carbon was in the bacteria (62%), but after longer incubation times (30 and 32 h in situ) the macrofauna gained more importance (15 and 48%, respectively), until after 132 h the greatest fraction was mineralized to CO2 (44%). Our findings show the rapid impact of the benthic sand community on a settling phytoplankton bloom and the great importance of bacteria in the first steps of algal carbon processing.
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
Introduction
Shelf seas cover one tenth of the ocean area, but contribute 30% to the world ocean primary production (Jørgensen Citation1996). In the northern temperate latitudes, large phytoplankton blooms occur in spring (Reid et al. Citation1990), where sand is the predominant sediment type on the shelf. Due to the shallowness of shelf seas, up to 50% of this biomass can settle to the sea floor (Jørgensen et al. Citation1990), building up the bulk of the food supply for the benthic community (Conley & Johnstone Citation1995). Most of the deposited material is directly mineralized in these sediments (Berner Citation1982; Jahnke et al. Citation2000), causing low organic carbon contents and emphasizing the prominent role of the continental shelf in the marine carbon cycle. In these sediments, the organic matter is degraded along aerobic and anaerobic pathways, with oxygen and sulphate as the most important electron acceptors (Jørgensen Citation1982). The bacterial communities inhabiting sands are regarded as highly active (Rusch et al. Citation2003; Ehrenhauss et al. Citation2004a), even though sandy sediments typically show lower bacterial densities than muddy sites (Llobet-Brossa et al. Citation1998). The high activity is probably caused by advective porewater transport (e.g. Boudreau et al. Citation2001), an exchange process driven by surface gravity waves or interactions between bottom currents and sediment topography or biogenic structures (Huettel & Rusch Citation2000; Huettel & Webster Citation2001).
The benthic community processes the arriving organic material by incorporation, excretion and respiration. Benthic organisms and interactions within the benthic community are thus thought to have a great impact on the fast turnover rates of settling particles in these environments. However, these interactions have been poorly studied. Information is available on the impact of settling phytoplankton on single benthic compartments like macrofauna (Boon et al. Citation1998), on the bacterial response after bloom events (Stoeck & Kröncke Citation2001) or on the decomposition of settling particles (Sun et al. Citation1997). However, the in situ determination of the benthic community structure is still one of the most urgent problems. Above all, information is missing on microbial communities. Investigations of microbial lipid biomarkers have tremendously increased our understanding of the microbial world. They represent a very useful tool to investigate microbial community patterns in situ (Rajendran et al. Citation1992; Hinrichs et al. Citation1999; Rütters et al. Citation2002a), as shown at our study site as well by Bühring et al. (Citation2005). The application of isotopically labelled substrates has made tracer studies on carbon cycling of microbial populations possible (Boschker et al. Citation1998; Middelburg et al. Citation2000; Moodley et al. Citation2000 Citation2002; Witte et al. Citation2003b).
The leading hypothesis for this study was to prove that sandy sediment communities are highly active and react rapidly on phytoplankton sedimentation events. We conducted in situ and on-board experiments simulating a phytoplankton bloom on a fine sandy station near Spiekeroog Island (German Bight, North Sea). To test our hypothesis, we carried out pulse-chase chamber experiments with 13C-labelled phytoplankton. The total uptake of algal carbon into macrofauna, the incorporation into bacterial biomarkers and its mineralization to 13CO2 was then followed by isotopic ratio mass spectrometry (IRMS).
Material and methods
Sites and sampling
During two cruises of RV Heincke in April (HE 145) and June 2001 (HE 148), experiments were conducted on a 19 m water depth fine sandy station in the southern North Sea, seaward of the island of Spiekeroog (53°51′N 007°44′E). The German Bight is a shallow part of the southern North Sea. Because of tides, waves and storm-generated bottom currents, the sediment is frequently eroded and redeposited (Antia Citation1995). The permeability was 3.02±1.66×10−12 m2 (n = 14; average core length was 110.29 mm) and the density was 2.06±0.02 g ml−1 (Janssen et al. 2005). The median grain size was 163±20 µm. The salinity of the sampling site was 31–32 and the water temperature was 9°C in April and 13°C in June. During HE 145 we conducted on-board incubations of 12, 30 and 132 h duration and during HE 148 an in situ experiment of 32 h was conducted.
Prior to the experiments, we cultured an axenic clone of the diatom Ditylum brightwellii (Bacillariophyceae, Biddulphiales) at 25°C in artificial seawater with a salinity of 33 (Grasshoff Citation1999) enriched with f/2 medium (Guillard & Ryther Citation1962). This medium contained 25% 13C-bicarbonate (99% NaH13CO3, Cambridge Isotope Laboratories). The algal material was harvested by centrifugation (404g, 4 min), rinsed three times with an isotonic sodium chloride solution and centrifuged again. The axenic state of the algae was verified by microscopic observation of 4′,6′-diamido-2-phenylimdole (DAPI) stained cells and by investigation of the fatty acid composition. Neither bacteria nor bacterial fatty acids could be detected. The algae consisted of 15 atom% 13C (HE 145) and 9 atom% 13C (HE 148). The amount of carbon added to each chamber corresponded to 0.31 g C m−2 (HE 145) and 0.36 g C m−2 (HE 148).
The experiments were performed in the dark in acrylic cylindrical chambers (Huettel & Gust Citation1992), 20 cm in diameter and 31 cm in height. A horizontal disk stirred a water column of approximately 10 cm height at 20 rpm. The rotating water generated a pressure gradient (ca. 1.5 Pa cm−2) comparable with the pressure gradient at a sediment ripple interacting with bottom currents (Huettel & Rusch Citation2000). This gradient induces advective porewater flows in permeable sediments.
Chambers for the on-board experiments were attached to a frame that was lowered to the sea floor by the ship's winch. They were inserted into the sediment by divers and the bottom was directly sealed with a lid before hauling the chambers back to the ship's laboratory. They were stirred directly after recovery and incubated at the in situ temperature in the ship's laboratory. For the in situ experiments the frame was deployed, divers inserted the chambers into the sediment, and also injected the algae into the chambers. After the incubation time of 32 h, divers closed the chambers at the sea floor. At the end of the experiments the sediment was sliced at 0.5 cm intervals down to 1 cm, 1 cm intervals down to 10 cm, and 2.5 cm intervals below 10 cm sediment depth. The macrofauna were carefully picked from every sediment horizon and the sediment was then homogenized before taking samples for porewater 13CO2 and lipid analysis. For the porewater δ13CO2 analysis, 20 ml of sediment was centrifuged (2800g, 10 min) through GF/F filters and 2 ml of the porewater was kept at 4°C in a 4 ml gas-tight glass scintillation vial containing mercury chloride (end concentration 0.2%). The macrofauna were sorted under the stereo-microscope and stored at −20°C. [For further details on the subsequent treatment of macrofauna samples, see Kamp & Witte (Citation2005)]. The lipid samples were stored in glass bottles at −20°C.
Sediment cores for background samples were taken with a small multicorer system equipped with four cores with an inner diameter of 36 mm. Sediment cores were sliced at the same depth intervals as for the experiments.
Lipid analysis
Lipids were extracted ultrasonically, four times for 20 min, from the wet sediment material with dichloromethane–methanol following an extraction protocol of Elvert et al. (Citation2003). The resulting dichloromethane extract was separated from the aqueous methanol phase by adding 0.05 M KCl solution. For saponification, an aliquot of the total extract was treated with 6% KOH in methanol for 3 h at 80°C. Neutral lipids were released by the addition of 0.05 M KCl for phase separation and extracted with hexane. HCl (25%) was added to the aqueous reaction mixture (pH < 1) and free fatty acids were extracted with hexane. The fatty acid fraction was then evaporated to near dryness and derivatized by adding 1 ml of BF3-methanol (12–14%, Merck) for 1 h at 60°C to yield fatty acid methyl esters (FAME).
The concentrations of FAMEs were determined by gas chromatography (GC)-flame ionization detection (Hewlett Packard 5890, series II), operating in splitless mode. A 50 m apolar HP-5 fused silica capillary column was used (0.32 mm internal diameter, 0.17 µm film thickness; Hewlett Packard). The carrier gas was He (2.0 ml min−1). The initial oven temperature was 60°C held for 1 min, increased to 150°C with 10°C min−1, then to 310°C with 4°C min−1 and finally kept at 310°C for 15 min. The injector temperature was set at 300°C and the detector at 310°C. C19:0 fatty acid was added prior to extraction as an internal standard for quantification. The identification of FAMEs was based on the comparison of retention time with known standards. Fatty acid short-hand nomenclature is according to Guckert et al. (Citation1985).
The identification of unknown compounds was carried out using a Thermoquest Trace GC interfaced to a Finnigan Trace MS using the same conditions. Mass spectra were collected (m/z 30–580, 0.9 scans s−1), operating in electron impact mode at 70 eV. The samples were injected in splitless mode at an injection temperature of 295°C on a HP-5 MS fused silica capillary column (60 m, internal diameter 0.32 mm; Hewlett Packard) coated with a cross-linked methyl silicone phase (film thickness 0.25 µm, Hewlett Packard). The detector was set at 350 V, the interface was 310°C and the carrier gas was He with a constant flow of 1.4 ml min−1.
The double bond positions of fatty acids were determined by the analysis of dimethyl disulphide adducts, prepared after the method of Nichols et al. (Citation1986).
The carbon isotope composition of FAMEs was determined by GC-c-IRMS. The MS (Finnigan Delta plus) was connected via a Finnigan Combustion Interface III to a HP 6890 Series GC equipped with a 50 m HP-5 (0.32 mm internal diameter, 0.17 µm stationary phase, Hewlett-Packard). The flow rate was 1.5 ml min−1. Samples were injected in splitless mode (injector temperature 290°C) and subjected to the same temperature programme as given for GC and GC-MS measurements. The oxidation and reduction ovens were operated at 940 and 640°C, respectively. The results were corrected for the introduction of the additional carbon atoms during derivatization with BF3-methanol.
13CO2 analysis
The water samples for the δ13CO2 measurements were filtered (0.2 µm) directly into 12 ml gas-tight glass scintillation vials sealed with a rubber septum and containing mercury chloride in an end concentration of 0.2% to stop all activity. The samples were stored refrigerated until the analysis.
Prior to the measurement, a headspace was created by injecting 3 ml of nitrogen gas and the sample was then acidified with sulphuric acid (20%). The δ13CO2 and the concentration of CO2 in the headspace was measured using a Carlo Erba 1106 elemental analyser coupled online with a Finnigan Delta S isotope ratio MS. Reproducibility of the measurements was ∼0.10‰.
Calculations
Carbon isotopic ratios (13C/12C) are expressed in the delta notation (δ13C) relative to Vienna Pee Dee Belemnite Standard (13C/12CVPDB=0.0112): δ13C (‰)=[(Rsample/Rstd)–1)]×1000, where Rsample and Rstd are the 13C/12C of the sample and the standard, respectively (Craig Citation1957). The total uptake, I, of 13C was calculated as the product of excess 13C (E) and macrofauna biomass (organic carbon content) or carbon content in CO2 concentrations. For bacteria, I was calculated after Middelburg et al. (2000) from label incorporation into bacterial fatty acids (iC15:0, aiC15:0, iC16:0, iC17:1ω7, 10Me-C16:0, iC17:0, aiC17:0, C17:1ω8 and C17:1ω6) as Ibact=∑ Ibact.fatty-acids/(a×b), where a is the average phospholipid-derived fatty acid (PLFA) concentration in bacteria of 0.056 g of carbon PLFA g−1 biomass (Brinch-Iversen & King Citation1990) and b is the average fraction-specific bacterial PLFA encountered in sediment dominated by bacteria (0.19; calculated after Findlay & Dobbs Citation1993; Rajendran et al. Citation1994; Guezennec & Fiala-Medioni Citation1996). E is the difference between the fraction F of the sample and background: E = Fsample–Fbackground, where F = 13C/(13C/12C) = R/(R + 1) and R=(δ13C/1000 + 1)×RVPDB with RVPDB=0.0112372.
The fatty acid composition of the added diatom Ditylum brightwellii was dominated by the fatty acid C14:0, C15:0, C16:3, C16:1ω7, and C16:0.
In our experiments, the amount of added particulate organic carbon (POC) (0.31 and 0.36 g C m−2) corresponds to half the daily primary production (0.5 g C m−2 day−1; Wollast Citation1991) in the southern North Sea. Thus, the experimental conditions were comparable with the natural environment with respect to the addition of organic matter.
Results
Incorporation of 13C into bacterial fatty acids
shows the total incorporation of label into selected bacterial fatty acids. 13C was detectable in all investigated bacterial fatty acids, although in varying amounts. The highest incorporation was found in iC16:0 and the lowest in 10Me-C16:0. IC16:0 showed a fast and high incorporation after 12 h, with decreasing values after 30 and 132 h. The fatty acids iC15:0 and aiC15:0 showed increasing incorporation up to 30 h experimental duration. Only minor incorporation with only slight changes over time was visible for iC17:1ω7, 10Me-C16:0, iC17:0, aiC17:0 and C17:1ω6. The fatty acid C17:1ω8 displayed a continuously increasing uptake, with maximum incorporation after 132 h. Bacterial incorporation of 13C was generally lower in the 32 h in situ experiment in June than in the 30 h on-board experiment in April.
Figure 1. Incorporation of algal 13C into bacterial fatty acids (µg 13C m−2) (sampling depth 0–12 cm) from the on-board incubations of 12, 30 and 132 h and the 32 h in situ experiments. The bars represent the mean value and the error bars depict the higher value of two replicates.
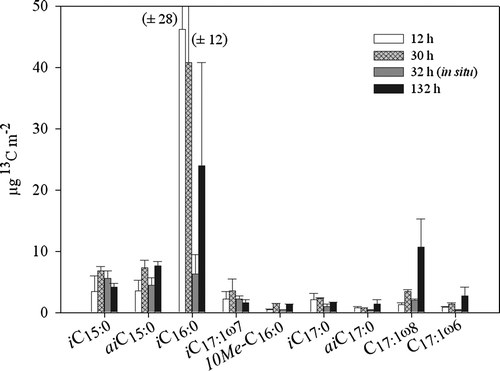
The total incorporation of tracer 13C in bacteria in the April experiments was relatively constant with time, with 5800 µg m−2 (12.4% of the added 13C) after 12 h, 6400 µg m−2 (13.8%) after 30 h and 5200 µg m−2 (11.2%) after 132 h. In situ, 2200 µg m−2 (6.7%) of the added label was detected in bacterial fatty acids after 32 h of incubation.
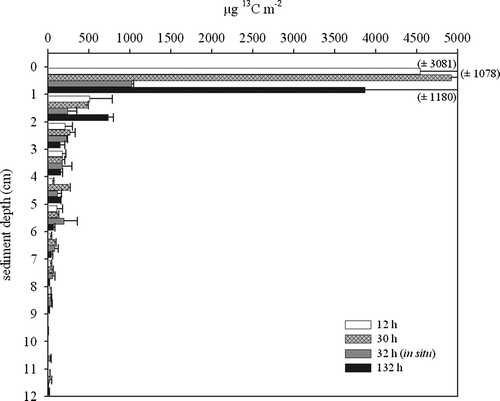
The vertical distribution of 13C incorporation into bacteria within the sediment is shown in . Bulk incorporation took place at the sediment surface during all incubations. After 12 h, labelled bacterial biomass was detectable down to 5 cm sediment depth.
Figure 2. Incorporation (µg 13C m−2) into bacteria versus sediment depth from the on-board incubations of 12, 30 and 132 h and the 32 h in situ experiments. The bars represent the mean value and the error bars indicate the range of two replicates.
Uptake of algal material by macrofauna
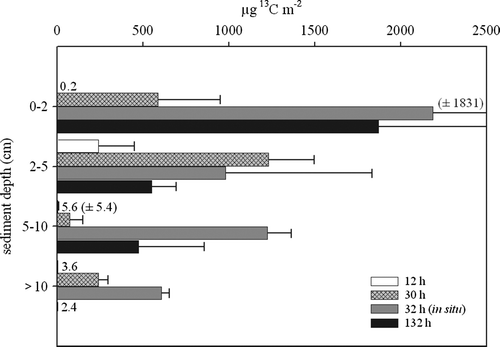
After 12 h, organisms at all sediment depths had ingested tracer material (). After 12 h, 250 µg m−2 (0.5%) of the added 13C label could be detected in macrofauna organisms. After 30 and 132 h, macrofauna had ingested 2100 µg m−2 (4.6%) and 2900 µg m−2 (6.2%), respectively. In the 32 h in situ experiment in June, we found a very high uptake by macrofauna of 5000 µg m−2 (15.5% of the added label).
Figure 3. Total uptake (µg 13C m−2) of label into macrofauna versus sediment depth during the 12, 30 and 132 h on-board incubations and the 32 h in situ experiment. The bars indicate the mean value and the error bars depict the standard deviation.
Mineralization of tracer POC to 13CO2
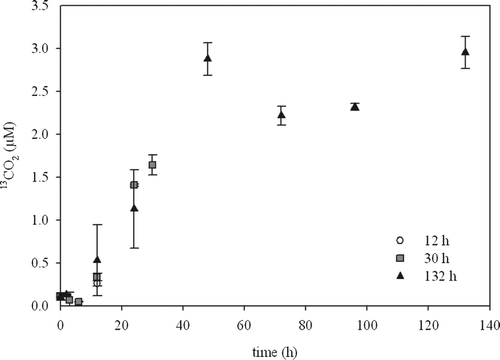
During the on-board experiments, chamber water 13CO2 was determined repeatedly during the experiments (). The labelling of CO2 increased over the course of the experiment. At the end of our experiments we found the following percentages of added label in the CO2 of the overlying water: 1% after 12 h, 6% after 30 h, 3% after 32 h (in situ) and 11% after 132 h.
Figure 4. Release of 13CO2 to overlying water (µM) over time for the 12, 30 and 132 h on-board incubations. The symbols indicate the mean value and the error bars depict the range of two parallels.
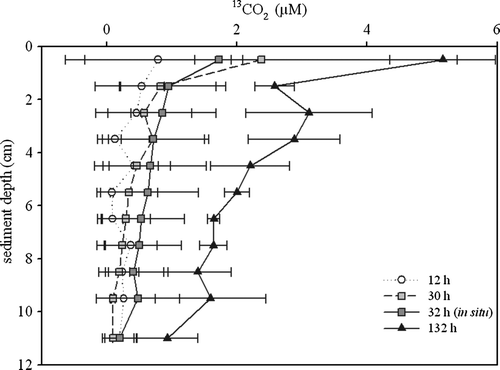
Mineralization of tracer POC was evident in the porewater as well, with increasing values after longer incubation times (). After 132 h incubation time, elevated 13CO2 values were detected over the whole sediment sampling depth.
Figure 5. Porewater 13CO2 (µM) versus sediment depth of the 12, 30 and 132 h on-board incubations and the 32 h in situ experiment. The symbols indicate the mean value and the error bars depict the range of two parallels.
Discussion
The decomposition of organic material in the sediment follows different pathways. At the sediment–water interface, aerobic mineralization takes place through the activity of benthic micro-organisms and animals. Below the oxygen penetration depth, mineralization occurs via anaerobic processes (Jørgensen Citation1983). The anaerobic food chain consists of different types of bacteria that mediate different metabolic steps, each of which is only partially completing the oxidation of the organic compounds. The participating organisms are usually denitrifying, sulphate-reducing (SRB) and methane-producing bacteria (Jørgensen Citation2000). Investigations of bacterial biomarkers allow differentiation between bacterial communities with different physiologies.
In our experiments, the bacterial community demonstrated the capability for fast processing organic carbon, but incorporation patterns into the investigated bacterial fatty acids were very different (). This was probably due to the different biosynthetic pathways and metabolic specificity of the bacteria. The branched-chained fatty acids iC15:0, aiC15:0 and especially iC16:0 showed fast and high label incorporation; these fatty acids must be biosynthesized quickly. They occur in several types of bacteria (Boschker & Middelburg Citation2002), but they are particularly abundant in Gram-positive prokaryotes (White et al. Citation1996). The fast incorporation of label indicates that these bacteria have direct access to the diatom material by either using exoenzymes or taking up dissolved organic carbon, which was already present in the added algal material (Ehrenhauss et al. Citation2004b). Middelburg et al. (Citation2000) also found rapid uptake into iC15:0, aiC15:0 and iC16:0 fatty acids. In their experiment on an intertidal sandy site in the Scheldt estuary, they sprayed 13C-bicarbonate on the surface and followed its path through the benthic food web. They found evidence for photosynthetically fixed 13C entering the microbial food web within hours and maximum labelling of bacterial fatty acids after 1 day.
Those fatty acids with a weak incorporation after 12 h, such as iC17:1ω7, 10Me-C16:0, iC17:0, aiC17:0, C17:1ω8 and C17:1ω6, are described as typical for Desulfobacter spp. (Boschker et al. Citation1998). Low label incorporation into C17 fatty acids was also observed by Boschker et al. (Citation1998) during an experiment with 13C-acetate. They concluded that Desulfobacter spp., which is the most commonly isolated SRB in coastal sediments, is probably of minor importance. The fatty acid C17:1ω8 is described as typical for Desulfomicrobium spp. (Rütters et al. Citation2002b). Our experiments demonstrated an increasing incorporation into C17:1ω8. This pattern could be caused by the incorporation of secondary carbon products (e.g. acetate and other short-chain carbon sources) produced by other organisms feeding directly on the diatoms, such as fermentative bacteria and meio- or macrofauna.
The total uptake of label into bacterial fatty acids versus sediment depth () was highest at the sediment surface. However, clear uptake into bacterial biomass was detectable down to 5 cm depth, especially during the 32 h in situ experiments.
As part of our project, the transport of algal-derived labeled total organic carbon (TO13C) into deeper sediment layers was observed (Ehrenhauss et al. Citation2004b). Uptake down to 6 cm sediment depth was found after 132 h. One possible explanation for this transport is advective porewater flow, although at our station with a permeability of 3.02±1.66×10−12 m2, advection should only cause transport down to 2 cm into the sediment (Huettel & Gust Citation1992). Recent results from Janssen et al. (Citation2005) using the in situ chamber system Sandy at the same station suggest that it is even less likely that advection is the predominant transport process at our station. They found negligible advective porewater exchange, which emphasizes the great importance of macrofauna-driven transport at our study site. Deeper penetration of algal-derived TOC, together with subsurface maxima in label uptake by bacteria and mineralization, must be caused by a different transport process, possibly by activity of macrofauna organisms. Even during the short experiments, the uptake into macrofauna was not restricted to distinct depth intervals (). The largest uptake was measurable in animals from the upper sediment layer, but after 12 h of incubation animals from all depths were already labelled. It is therefore possible that the deep penetration of labelled particles in our experiments was due to the activity of macrofauna organisms.
The transport of POM by the activity of bioturbating animals was also observed by Sun et al. (Citation1999) and Blair et al. (Citation1996) using labelling experiments. Ehrenhauss et al. (2004b) found labelled TOC 4 cm deep in the sediment after 30 h of incubation and we found increased uptake into bacterial fatty acids at the same depth. After 32 h in situ experiment, the TO13C was found down to 6 cm deep in the sediment and a peak of bacterial labelling was found at 4.5 cm sediment depth. Considering these results it seems likely that the transport of TO13C by bioturbating animals allows bacteria from deeper layers to access relatively fresh POM.
Macrofaunal organisms also digest the algae and alter them by passage through their gut system. This may also influence the availability of POC for other (micro-) organisms. Some of the detected SRB, such as Desulfosarcina, are capable of autotrophic pathways. Therefore, the uptake of 13C-bicarbonate released as 13CO2 by macro- and meiofauna inhabiting deeper sediment layers could be another explanation for subsurface incorporation into bacterial fatty acids.
Five per cent of the added label was converted to CO2 per day. Janssen et al. (Citation2005) found an average total oxygen uptake of 29.2 mmol O2 m−2 day−1 at the same station. We may therefore conclude that only 2.4% of total respiration was due to our added algae (assuming an Respiratory Quotient (RQ) of 1), revealing that the magnitude of carbon addition was similar to the settling POC at the study site.
displays the recovered label in the different compartments. The great importance of bacteria on the carbon turnover in this sediment is clearly visible. These results emphasize their importance for the very first reaction after the arrival of settling phytoplankton blooms. After longer incubation times, the macrofauna gain more importance, accompanied by increasing mineralization.
Table I. Biomass of macrofauna and bacteria and the recovered label in the different benthic compartments of the two parallels of the 12, 30 and 132 h experiments of the April cruise and the 32 h in situ experiment of the June cruise.
Differences between the on-board and in situ experiments were mainly due to macrofaunal uptake. This was probably caused by seasonal variations in abundance, which was doubled in June (1531±292 individuals m−2) compared with April (771±287 individuals m−2) (Kamp & Witte Citation2005). Meio- and macrofauna can significantly reduce the bacterial population due to their feeding activity. This could be an explanation for the lower bacterial incorporation, especially during the June in situ experiment with high macrofaunal abundances.
In total, 69 mg C m−2 day−1 was processed during the on-board incubations (based on the 12 and 30 h experiments). Compared with other investigations using the same labelling approach, we found much higher values than detected in the abyssal deep sea (4 mg C m−2 day−1; Witte et al. Citation2003b) and the deep sea at 2100 m depth (4.2 mg C m−2 day−1; Moodley et al. Citation2002) or on the continental slope (19 mg C m−2 day−1; Witte et al. Citation2003a). We have to take into account that we added only approximately half the daily primary production (0.5 g C m−2 day−1; Wollast Citation1991), implying that the potential for overall carbon processing in this sediment is considerably higher. Our results clearly support our hypothesis that sandy sediments are highly active, contrasting the former belief that they are biogeochemical deserts due to their low organic carbon content (Boudreau et al. Citation2001) and lower bacterial abundances compared with finer compartments (Llobet-Brossa et al. Citation1998). In accordance with investigations of Moodley et al. (Citation2005), it is also possible that our turnover rates were enhanced compared with previous studies, caused by higher water temperatures at our study site.
Our experiments revealed the great importance of the benthic bacterial community on the short-term fate of settling phytoplankton blooms. Graf (Citation1992) demonstrated that bacteria are responsible for the majority of the oxygen consumption. Investigations of the benthic reaction on phytoplankton input by Boon et al. (Citation1998) at several stations in the North Sea revealed a temporal uncoupling of macrofaunal and bacterial processes. In the present study, we observed that bacteria, macrofauna and mineralization follow different time patterns in their short-term reaction.
Conclusions
The investigation of turnover on a sandy North Sea station revealed a very rapid reaction of the benthic community to experimental POM addition. After 12 h, elevated values were detectable, with an incorporation of 6.4 mg 13C m−2 into bacteria, 0.3 mg 13C m−2 into macrofauna and 0.9 mg 13C m−2 found in the CO2. After 30 h the incorporation into bacteria remained constant, but the mineralization increased to over 3 mg 13C m−2 and uptake into the macrofauna increased 10-fold compared with the 12 h experiment. Finally, after 132 h the bacteria had incorporated 5.2 mg 13C m−2, the macrofauna had taken up 2.9 mg 13C m−2 and we found 8.1 mg 13C m−2 in the CO2. For the in situ experiments in June, we found 2.2 mg 13C m−2 in the bacteria, 5 mg 13C m−2 in the macrofauna and 1.9 mg 13C m−2 in the CO2. Incorporation into bacterial biomarkers varied, indicating different metabolic groups of bacteria in the sediment, with either direct access to the added material or SRB, which take up, e.g. acetate or other short-chain carbon products.
Editorial responsibility: Joel Kostka
Acknowledgments
M. Elvert is thanked for his invaluable support with the fatty acid analysis. F. Janssen and the divers are gratefully acknowledged for their help during the cruises. The captain and the crew of the RV Heincke are thanked for their assistance during the two cruises. A. Pernthaler is acknowledged for providing an axenic clone of Ditylum brightwellii. We thank G. Klockgether for her help with GC-c-IRMS measurements. U. Struck from the University of Munich is gratefully acknowledged for measuring the isotopic composition of the macrofauna. We are grateful to L. R. Hmelo for revising the English text and to two anonymous reviewers for constructive comments on the manuscript. B. B. Jørgensen and the Max-Planck Society are thanked for their support of this work.
Additional information
Notes on contributors
S. I. Bühring
Present address: DFG-Research Centre Ocean Margins, University of Bremen, P.O. Box 330440, 28334 Bremen, GermanyS. Ehrenhauss
Present address: Institute for Microbiology, University of Hannover, Schneiderberg 50, 30176 Hannover, GermanyU. Witte
Present address: University of Aberdeen, Oceanlab, Newburgh, Aberdeenshire AB41 6AA, UKNotes
Published in collaboration with the University of Bergen and the Institute of Marine Research, Norway, and the Marine Biological Laboratory, University of Copenhagen, Denmark
References
- Antia , EE . 1995 . Sedimentory deposits related to inlet–shoreface storm flow interaction in the German Bight. Estuarine . Coastal and Shelf Sciences , 40 : 699 – 712 .
- Berner , R . 1982 . Burial of organic carbon and pyrite sulfur in the modern ocean: its geochemical and environmental significance . American Journal of Science , 282 : 451 – 73 .
- Blair , NE , Levin , LA , DeMaster , DJ and Plaia , G . 1996 . The short-term fate of fresh algal carbon in continental slope sediments . Limnology and Oceanography , 41 : 1208 – 19 .
- Boon , AR , Duineveld , GCA , Berghuis , EM and van der Weele , JA . 1998 . Relationships between benthic activity and the annual phytopigment cycle in near-bottom water and sediments in the southern North Sea . Estuarine, Coastal and Shelf Sciences , 46 : 1 – 13 .
- Boschker , HTS and Middelburg , JJ . 2002 . Stable isotopes and biomarkers in microbial ecology . FEMS Microbiology Ecology , 40 : 85 – 95 .
- Boschker , HTS , Nold , SC , Wellsbury , P , Bos , D , de Graaf , W , Pel , R , Parkes , RJ and Cappenberg , RE . 1998 . Direct linking of microbial populations to specific biogeochemical processes by 13C-labelling of biomarkers . Nature , 392 : 801 – 5 .
- Boudreau , BP , Huettel , M , Forster , S , Jahnke , RA , McLachlan , A , Middelburg , JJ , Nielsen , P , Sansone , F , Taghon , G Van Raaphorst , W . 2001 . Permeable marine sediments: overturning an old paradigm. Eos, Transactions . American Geophysical Union , 82 : 133 – 40 .
- Brinch-Iversen , J and King , GM . 1990 . Effects of substrate concentration, growth state, and oxygen availability on relationships among bacterial carbon, nitrogen and phospholipid phosphorous content . FEMS Microbiology Ecology , 74 : 345 – 56 .
- Bühring , SI , Elvert , M and Witte , U . 2005 . The microbial community structure of different permeable sandy sediments characterised by the investigation of bacterial fatty acids and fluorescence in situ hybridisation . Environmental Microbiology , 7 : 281 – 93 .
- Conley , DJ and Johnstone , RW . 1995 . Biogeochemistry of N, P, Si in Baltic Sea sediments: response to a simulated deposition of a spring diatom bloom . Marine Ecology Progress Series , 122 : 265 – 76 .
- Craig , H . 1957 . Isotopic standards for carbon and oxygen and correction factors for mass-spectrometric analysis of carbon dioxide . Geochimica Cosmochimica Acta , 12 : 133 – 49 .
- Ehrenhauss , S , Witte , U , Janssen , F and Huettel , M . 2004a . Decomposition of diatoms and nutrient dynamics in permeable North Sea sediments . Continental Shelf Research , 24 : 721 – 37 .
- Ehrenhauss , S , Witte , U , Bühring , SI and Huettel , M . 2004b . Effect of pore water transport on distribution and degradation of diatoms in permeable North Sea sediments . Marine Ecology Progress Series , 271 : 99 – 111 .
- Elvert , M , Boetius , A , Knittel , K and Jørgensen , BB . 2003 . Characterization of specific membrane fatty acids as chemotaxonomic markers for sulfate-reducing bacteria involved in anaerobic oxidation of methane . Geomicrobiology Journal , 20 : 403 – 19 .
- Findlay , RH and Dobbs , FC . 1993 . “ Quantitative description of microbial communities using lipid analysis ” . In Handbook of Methods in Aquatic Microbial Ecology , Edited by: Kemp , PF , Sherr , BF , Sherr , EB and Cole , JJ . 271 – 84 . Boca Raton : Lewis Publishers .
- Graf , G . 1992 . Benthic–pelagic coupling: a benthic review . Oceanography and Marine Biology: an Annual Review , 30 : 149 – 90 .
- Grasshoff , K . 1999 . Methods of Seawater Analysis , Weinheim : Wiley-VCH .
- Guckert , JB , Antworth , CP , Nichols , PD and White , DC . 1985 . Phospholipid, ester-linked fatty acid profiles as reproducible assays for changes in procaryotic community structure of estuarine sediments . FEMS Microbiology Ecology , 31 : 147 – 58 .
- Guezennec , J and Fiala-Medioni , A . 1996 . Bacterial abundance and diversity in the Barbados Trench determined by phospholipid analysis . FEMS Microbiology Ecology , 19 : 83 – 93 .
- Guillard , RRL and Ryther , JH . 1962 . Studies of marine planktonic diatoms: 1. Cyclotella nana hustedt and Detonula confervacea (cleve) gran . Canadian Journal of Microbiology , 8 : 229 – 39 .
- Hinrichs , KU , Hayes , JM , Sylva , SP , Brewer , PG and DeLong , EF . 1999 . Methane-consuming archaebacteria in marine sediments . Nature , 398 : 802 – 5 .
- Huettel , M and Gust , G . 1992 . Impact of bioroughness on intertidal solute exchange in permeable sediments . Marine Ecology Progress Series , 89 : 253 – 67 .
- Huettel , M and Rusch , A . 2000 . Transport and degradation of phytoplankton in permeable sediments . Limnology and Oceanography , 45 : 534 – 49 .
- Huettel , M and Webster , IT . 2001 . “ Porewater flow in permeable sediments ” . In The Benthic Boundary Layer , Edited by: Boudreau , BP and Jørgensen , BB . 144 – 79 . Oxford : University Press .
- Jahnke , RA , Nelson , JR , Marinelli , RL and Eckman , JE . 2000 . Benthic flux of biogenic elements on the Southeastern US continental shelf: influence of pore water advective transport and benthic microalgae . Continental Shelf Research , 20 : 109 – 27 .
- Janssen , F , Huettel , M and Witte , U . 2005 . The impact of advection on benthic fluxes of natural permeable beds (II): organic matter remineralization of three neighboring sites of contrasting permeability (German Bight, southern North Sea) . Limnology and Oceanography , 50 : 779 – 92 .
- Jørgensen , BB . 1982 . Mineralization of organic matter in the sea bed – the role of sulfate reduction . Nature , 296 : 643 – 5 .
- Jørgensen , BB . 1983 . “ Processes at the sediment water interface ” . In The Major Biogeochemical Cycles and their Interactions , Edited by: Bolin , B and Cook , RB . 477 – 515 . New York : Wiley .
- Jørgensen , BB . 1996 . “ Material flux in the sediment ” . In Coastal and Estuarine Studies , Edited by: Jørgensen , BB and Richardsen , K . 115 – 35 . Washington, DC : American Geophysical Union .
- Jørgensen , BB . 2000 . “ Bacteria and marine biogeochemistry ” . In Marine Geochemistry , Edited by: Schulz , HD and Zabel , M . 173 – 207 . Berlin : Springer .
- Jørgensen , BB , Bang , M and Blackburn , TH . 1990 . Anaerobic mineralization in marine sediments from the Baltic Sea–North Sea transition . Marine Ecology Progress Series , 59 : 39 – 54 .
- Kamp , A and Witte , U . 2005 . Processing of 13C-labeled phytoplankton in a fine-grained sandy-shelf sediment (North Sea): relative importance of different macrofauna species . Marine Ecology Progress Series , 297 : 61 – 70 .
- Llobet-Brossa , E , Rossello-Mora , R and Amann , R . 1998 . Microbial community composition of Wadden Sea sediments as revealed by fluorescence in situ hybridization . Applied Environmental Microbiology , 64 : 2691 – 6 .
- Middelburg , JJ , Barranguet , C , Boschker , HTS , Herman , PMJ , Moens , T and Heip , CHR . 2000 . The fate of intertidal microphytobenthos: an in situ 13C labelling study . Limnology and Oceanography , 45 : 1224 – 34 .
- Moodley , L , Boschker , HTS , Middelburg , JJ , Pel , R , Herman , PMJ , de Deckere , E and Heip , CHR . 2000 . Ecological significance of benthic foraminifera: 13C labelling experiments . Marine Ecology Progress Series , 202 : 289 – 95 .
- Moodley , L , Middelburg , JJ , Boschker , HTS , Duineveld , GCA , Pel , R , Herman , PMJ and Heip , CHR . 2002 . Bacteria and foraminifera: key players in a short-term deep-sea benthic response to phytodetritus . Marine Ecology Progress Series , 236 : 23 – 9 .
- Moodley , L , Middelburg , JJ , Soetaert , K , Boschker , HTS , Herman , PMJ and Heip , CHR . 2005 . Similar rapid response to phytodetritus deposition in shallow and deep-sea sediments . Journal of Marine Research , 63 : 457 – 69 .
- Nichols , PD , Guckert , JB and White , DC . 1986 . Determination of monounsaturated fatty acid double-bond position and geometry for microbial monocultures and complex consortia by capillary GC-MS of their dimethyl disulphide adducts . Journal of Microbiological Methods , 5 : 49 – 55 .
- Rajendran , N , Matsuda , O , Imamura , N and Urushigawa , Y . 1992 . Determination of microbial biomass and its community structure from the distribution of phospholipid ester-linked fatty acids in sediments of Hiroshima Bay and its adjacent bays . Estuarine, Coastal and Shelf Sciences , 36 : 501 – 14 .
- Rajendran , N , Matsuda , O , Urushigawa , Y and Simidu , U . 1994 . Characterization of microbial community structure in the surface sediment of Osaka Bay, Japan, by phospholipid fatty acid analysis . Applied Environmental Microbiology , 60 : 248 – 59 .
- Reid , PC , Lancelot , C , Gieskes , WWC , Hagmeier , E and Weichart , G . 1990 . Phytoplankton of the North Sea and its dynamics: a review . Netherlands Journal of Sea Research , 26 : 295 – 331 .
- Rusch , A , Huettel , M , Reimers , CE , Taghon , GL and Fuller , CM . 2003 . Activity and distribution of bacterial populations in Middle Atlantic Bight shelf sands . FEMS Microbiology Ecology , 44 : 89 – 100 .
- Rütters , H , Sass , H , Cypionka , H and Rullkötter , J . 2002a . Microbial communities in a Wadden Sea sediment core – clues from analyses of intact glyceride lipids, and released fatty acids . Organic Geochemistry , 33 : 803 – 16 .
- Rütters , H , Sass , H , Cypionka , H and Rullkötter , J . 2002b . Phospholipid analysis as a tool to study complex microbial communities in marine sediments . Journal of Microbiological Methods , 48 : 149 – 60 .
- Stoeck , T and Kröncke , I . 2001 . Influence of particle mixing on vertical profiles of chlorophyll a and bacterial biomass in sediments of the German Bight, Oyster Ground and Dogger Bank (North Sea). Estuarine . Coastal and Shelf Sciences , 52 : 783 – 95 .
- Sun , MY , Aller , RC , Lee , C and Wakeham , SG . 1999 . Enhanced degradation of algal lipids by benthic macrofaunal activity: effect of Yoldia limatula . Journal of Marine Research , 57 : 775 – 804 .
- Sun , MY , Wakeham , SG and Lee , C . 1997 . Rates and mechanisms of fatty acid degradation in oxic and anoxic coastal marine sediments of Long Island Sound, New York, USA . Geochimica et Cosmochimica Acta , 61 : 341 – 55 .
- White , DC , Stair , JO and Ringelberg , DB . 1996 . Quantitative comparisons of in situ microbial biodiversity by signature biomarker analysis . Journal of Industrial Microbiology , 17 : 185 – 96 .
- Witte , U , Aberle , N , Sand , M and Wenzhöfer , F . 2003a . Rapid response of deep-sea benthic community to POM enrichment: an in situ experimental study . Marine Ecology Progress Series , 251 : 27 – 36 .
- Witte , U , Wenzhofer , F , Sommer , S , Boetius , A , Heinz , P , Aberle , N , Sand , M , Cremer , A , Abraham , WR Jørgensen , BB . 2003b . In situ experimental evidence of the fate of a phytodetritus pulse at the abyssal sea floor . Nature , 424 : 763 – 6 .
- Wollast , R . 1991 . “ The coastal organic carbon cycle: fluxes, sources, and sinks ” . In Ocean Margin Processes in Global Change , Edited by: Mantoura , RFC , Martin , J-M and Wollast , R . 365 – 81 . Chichester : John Wiley & Sons .