Abstract
Viewing objects can result in automatic, partial activation of motor plans associated with them—“object affordance”. Here, we recorded grip force simultaneously from both hands in an object affordance task to investigate the effects of conflict between coactivated responses. Participants classified pictures of objects by squeezing force transducers with their left or right hand. Responses were faster on trials where the object afforded an action with the same hand that was required to make the response (congruent trials) compared to the opposite hand (incongruent trials). In addition, conflict between coactivated responses was reduced if it was experienced on the preceding trial, just like Gratton adaptation effects reported in “conflict” tasks (e.g., Eriksen flanker). This finding suggests that object affordance demonstrates conflict effects similar to those shown in other stimulus–response mapping tasks and thus could be integrated into the wider conceptual framework on overlearnt stimulus–response associations. Corrected erroneous responses occurred more frequently when there was conflict between the afforded response and the response required by the task, providing direct evidence that viewing an object activates motor plans appropriate for interacting with that object. Recording continuous grip force, as here, provides a sensitive way to measure coactivated responses in affordance tasks.
Appropriate, goal-directed interaction with our environment requires a close relationship between perception and action. Indeed, some argue that object representations in the brain are not simply perceptual but also include possibilities for action. Gibson Citation(1986) introduced the notion of “affordances”: properties of an object that somehow automatically activate possible actions towards that object. The effects of object affordance have been demonstrated using both behavioural and imaging methods. For example, functional imaging studies have revealed that medial frontal motor regions, such as the supplementary motor area (SMA), can be activated even when observers simply look at graspable objects (e.g., Grèzes & Decety, Citation2002). Reaction time (RT) studies have also demonstrated that responses to an object are facilitated when they are congruent with actions relevant for interacting with that object. For example, Tucker and Ellis Citation(1998) showed participants pictures of objects, which they had to classify as upright or inverted by responding with either the left or the right hand. Critically, objects were positioned so they readily afforded an action with one of the hands—for example, a teapot with the handle on the right affords a grasping action with the right hand. Even though the action afforded by the object was irrelevant to the classification task, responses were faster if they were made with the same hand as the afforded response (“congruent” trial) than if they were made with the opposite hand (“incongruent” trial).
Object affordance versus stimulus–response (S–R) mapping tasks
Although Gibson Citation(1986) considered object affordances to be somehow “automatic”, it is important to consider the possible mechanisms underlying affordance effects. Object affordance tasks require participants to categorize target objects and make their response according to a given rule (e.g., to classify the object as upright or inverted). Congruency effects have also been reported for tasks using very different stimulus–response (S–R) mappings to those employed in affordance tasks. For example, in the influential Eriksen flanker paradigm (e.g., B. A. Eriksen & Eriksen, Citation1974), participants make speeded responses to a centrally presented target stimulus—for example, an arrow, flanked by nontarget (distractor) arrows. The trial is “congruent” if the target and flankers are associated with the same response (e.g., << <<< ) and “incongruent” if target and flankers are associated with different responses (e.g., >><>>). Responses are slower when there are incongruent compared to congruent flankers because there is conflict between coactivated responses to both target and flankers.
Such coactivation of responses occurs in flanker tasks because the arrow stimuli have been mapped to particular responses; also flanker arrows automatically activate the responses associated with them even though they are not the response required to the target. In object affordance tasks, perhaps the object—or part of the object—primes the observer to respond in a particular way. The primed response can be either congruent or incongruent with the response required to interact with the object. Therefore, it might be possible to consider affordance effects within a more generalized conceptual framework whereby automatic sensorimotor activation is elicited by previously learnt S–R associations.
Recently, researchers have begun to investigate the relationship between object affordance effects and general S–R associations. In particular, how affordance might compare to Simon effects (Simon, Citation1969) has been examined by pitting these two against each other, often in a single experiment (e.g., Iani, Baroni, Pellicano, & Nicoletti, 2011; Pellicano, Iani, Borghi, Rubichi, & Nicoletti, 2010; Symes, Ellis, & Tucker, Citation2005). For example, Symes and colleagues reported three experiments in which participants made spatial responses to objects that simultaneously varied in both orientation (affordance effect) and location (Simon effect). These studies showed that dissociable Simon and affordance effects can co-occur. However, such findings do not necessarily clarify the potential extent of overlap between the mechanisms producing Simon (and other S–R mappings) and affordance effects.
One way to investigate whether affordance might fit into a wider framework of S–R associations would be to examine whether affordance effects show the same behavioural phenomena as those reported in other S–R tasks. The nature of the mechanisms responsible for producing congruency effects in S–R tasks (e.g., Simon effects) has been investigated by plotting the size of the congruency effects as a function of response speed, known as a “delta” plot. Delta plots from classic Simon tasks typically show initial positive effects that level off—or even become negative—as response times get longer (e.g., De Jong, Liang, & Lauber, 1994; see also van den Wildenberg et al., Citation2010, for a review). This pattern is usually explained in terms of suppression of unwanted responses activated by irrelevant stimuli by cognitive control mechanisms. The inhibition takes time to develop (e.g., Ridderinkhof, Citation2002; see also Eimer & Schlaghecken, Citation1998), and so a pattern of facilitation followed by inhibition is usually demonstrated in the RT distribution (Ridderinkhof, Citation2002).
As possible actions afforded by the environment will often conflict with the current goals of the observer, it follows that any motor responses automatically primed by object affordances might also need to be suppressed. Such inhibition of actions automatically elicited by object affordances might be expected to result in decreased congruency effects as RTs increase in affordance experiments—yielding delta plots similar to those shown in Simon effects. However, this is not the result usually reported.
Instead, affordance effects typically build up over time so that the size of the effect increases as RTs get longer and dissipate rapidly once the object has disappeared (e.g., Cho & Proctor, Citation2010; Derbyshire, Ellis, & Tucker, 2006; Iani et al., Citation2011; Pellicano et al., Citation2010; Tucker & Ellis, Citation2001; see also Phillips & Ward, Citation2002). Thus, affordance studies often show “positive” or “flat” delta plots. Importantly, “negative” delta plots are not always produced by S–R mapping tasks that are assumed to involve inhibition: Flat or even positive delta plots have emerged from some Eriksen flanker studies (Wylie et al., Citation2009) and from Simon tasks when the S–R mapping is more complex than a simple left–right button press with the left–right hand (e.g., when response hands are crossed or when stimuli and responses are arranged vertically; Wascher, Schatz, Kuder, & Verleger, 2001).
As well as evidence of cognitive control over coactivated responses that acts within a trial in S–R mapping tasks, there is also evidence of cognitive control over competing responses that acts between trials (e.g., Botvinick, Braver, Barch, Carter, & Cohen, 2001). For example, interference from flankers is weaker if an incongruent trial is preceded by another incongruent trial than if it is preceded by a congruent trial—the so-called “Gratton” effect (e.g., Gratton, Coles, & Donchin, Citation1992; Gratton, Coles, Sirevaag, Eriksen, & Donchin, Citation1988). It has been suggested that the conflict experienced on an incongruent trial ramps up levels of cognitive control, which in turn leads to reduced interference from flankers if the subsequent trial is also incongruent relative to if the preceding trial had been congruent (e.g., Botvinick et al., Citation2001).
This pattern of trial-to-trial effects has been reported for a range of S–R paradigms including the Simon task (e.g., Wühr & Ansorge, Citation2005) and the Stroop colour-word naming task (e.g., Egner & Hirsch, Citation2005), but to the best of our knowledge has not been investigated in object affordance. If object affordance effects fit into a wider framework of S–R associations, one might expect them, too, to produce similar types of adaptation in the presence of conflict as is shown in other S–R mapping paradigms.
Direct measurement of coactivated responses
Most experimental paradigms used to investigate automatic sensorimotor activation (e.g., Simon, Eriksen flanker) have shown the effects of conflict between coactivated responses indirectly by comparing response times for making manual button-press responses. Longer RTs are usually explained in terms of extra time required to resolve conflict, and errors are typically infrequent. But these button-press responses are an all-or-nothing, binary measure. Small amounts of force applied erroneously to the incorrect response button might be insufficient to trigger a response and therefore escape detection. Thus, such binary button-press measures might not be ideal for studying the effects of cognitive control because they cannot directly measure simultaneously activated motor plans.
Conflict between coactivated responses has been shown more directly by measuring responses that are continuous—for example, using electromyography (EMG) or event-related potentials (ERPs) or by measuring response as continuous force in “conflict tasks” (e.g., Burle, Allain, Vidal, & Hasbrouq, Citation2005; Coles, Gratton, Bashore, Eriksen, & Donchin, Citation1985; C. W. Eriksen, Coles, Morris, & O'Hara, Citation1985; Fournier, Scheffers, Coles, Adamson, & Abad, Citation1997). For example, C. W. Eriksen et al. Citation(1985) had participants complete a flanker task while recording continuous muscle activity (using EMG) simultaneously from both arms. On incongruent trials, correct button presses were often accompanied by muscle activity for the opposite response—that is, appropriate for the response associated with the flankers on incongruent trials. This provides direct evidence for simultaneous coactivation of responses associated with the target and flankers on incongruent trials.
To the best of our knowledge, continuous measures of response have not yet been applied to object affordance experiments. Here we introduce a continuous-force measure to study affordances, with participants making their responses by applying force to one of two transducers that recorded continuous grip force applied from each hand. This allows us potentially to detect whether a healthy observer viewing an object activates motor plans appropriate for interacting with that object, even if those motor plans are not relevant for the task. In addition, if object affordance effects are simply another example of S–R mapping, then one might expect them to produce similar types of conflict adaptation shown in other S–R mapping tasks. Therefore, one might anticipate negative sloping RT delta plots and reliable Gratton effects in a typical object affordance task, just as they are shown in other S–R mapping tasks.
Method
Participants
A total of 25 healthy adults (17 female; age 19–32 years) took part after giving informed consent. All participants self-reported normal or corrected-to-normal vision and right-handedness (Edinburgh handedness inventory; Oldfield, Citation1971).
Stimuli and apparatus
Stimuli were displayed on a 21″ CRT monitor (1,024 × 768 pixels), which participants viewed binocularly from 60 cm. Stimulus timing and presentation was locked to the screen refresh rate of 100 Hz. Stimuli were presented using a PC running Presentation software (Version 13.1; http://www.neurobs.com, Albany, CA: Neurobehavioral Systems). These were pictures of 10 household objects taken from the Object Databank (courtesy of Michael J. Tarr, Brown University, retrieved May 10, 2010, from http://www.tarrlab.org/) and Verfaillie and Bousten's 3D object database (see Boutsen, Lamberts, & Verfaillie, Citation1998; Verfaillie & Boutsen, Citation1995). Objects were matched for orientation.
Five objects belonged in a kitchen (fork, frying pan, knife, saucepan, spoon), and five in a toolbox (chisel, pliers, saw, screwdriver, spanner). Images subtended 10.6–17.3° horizontally and 2.8–5.3° vertically. Objects presented were oriented with their handles affording an action with the left or right hand.
Participants' responses were measured using two specially designed devices, one held in each hand. Participants held a rolled aneroid sphygmomanometer cuff (Boso-clinicus I, ref: 030-0-111), inflated to 20 mm Hg, connected to a pressure transducer. They were instructed to squeeze the rolled cuff in response to the object. Grip force was converted to voltage, which was digitized and stored using a LabJack U3 HV data acquisition device with DAQFactory Express (version 5.82; Azeo Tech Inc.) software. Data were sampled at 1,000 Hz.
Design and procedure
Before the experiment began, participants were shown the pressure transducers and practised making responses while observing the output from the pressure transducers on a computer screen. During the experiment, each trial began with presentation of a black fixation cross on a white background (). This cross subtended 1° × 1° and was presented in the centre of the screen for 500 ms. Following a blank interval (200 ms), the target object was presented at screen centre for 2,000 ms. Depending on the stimulus–response mapping given (counterbalanced across individuals), participants were instructed either to respond with the left hand to kitchen items and with the right hand to toolbox items, or vice versa. Thus, the hand action afforded by the object could be either “congruent” or “incongruent” with the required response.
Figure 1 Illustration of a participant responding to the stimulus sequence presented on an example trial. Participants made a squeeze response with either the left or the right hand to indicate whether the object presented belonged in either a kitchen or a toolbox. Each object was presented so that it afforded an action with the left or right hand (all trial types were equiprobable).
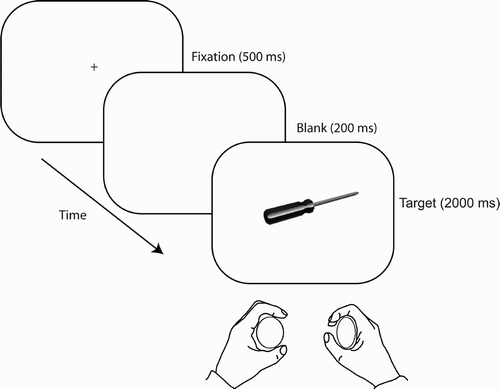
A blank intertrial interval (1,000 ms) was presented before the next trial began. Following a short practice block (12 trials), the experiment was arranged into four blocks of 64 trials, with rests between blocks. There were an equal number of congruent and incongruent trials with targets of each category (kitchen or toolbox) in each block (including the practice block). Order of presentation was shuffled randomly for each participant, and which image of the target category was presented was determined randomly and independently on a trial-by-trial basis. Participants were encouraged to respond as quickly as possible while remaining a high level of accuracy, but no response feedback was given to participants during the experiment.
Data analysis
Continuous-force recordings were locked to stimulus onset and epoched into periods of 2,500 ms, beginning 500 ms before target onset. Data were smoothed using a simple 5-point moving average to reduce high-frequency noise. The resulting waveforms were baseline corrected on a trial-by-trial basis according to the average baseline activity for each response device during the 200-ms prestimulus period on each trial.
A response (either correct or incorrect) was said to have occurred in a trial if at any point after stimulus onset until the end of the trial, the force measured was greater than 3 standard deviations from the mean force measured during the prestimulus baseline period and was followed by at least 8/10 points that also reached this threshold. Response onset time (RT) was defined as the first point that satisfied these criteria. Peak response was determined as the maximum amplitude of the response made in a trial that was surrounded by points on either side with the same or lower amplitude.
Results and discussion
Overall reaction times are modulated by congruency
A Wilcoxon signed-ranks test revealed that there was no reliable effect of trial congruency on peak response amplitude (Wilcoxon signed ranks test, Z = 0.28, p > .1). The effects of congruency on the peak response amplitude measure are not analysed any further.
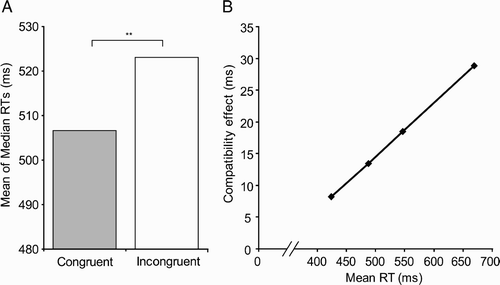
Median correct response times were calculated separately for each condition (congruent or incongruent) for each participant (. A repeated measures t test on median RTs showed that responses were significantly faster for congruent (507 ms) than for incongruent (523 ms) trials, t(24) = 3.49, p < .01. This result held when data were further separated by responding hand (average congruency effect in left hand: 16 ms; right hand: 17 ms) or target type (average congruency effect for kitchen objects: 18 ms; toolbox objects: 15 ms; all ps < .05). This is consistent with affordance effects previously documented using button-press or switch-press responses (e.g., Derbyshire et al., Citation2006; Ellis & Tucker, Citation2000; Tucker & Ellis, Citation1998; Vainio, Ellis, & Tucker, 2007).
Figure 2 (A). Mean of median correct response times (RTs) were significantly faster for congruent than for incongruent trials, which replicates affordance effects reported using button presses. **Denotes p < .01. (B). Mean quartile compatibility effect sizes as a function of mean RT for that quartile.
RT distributional analysis
In order to further explore the time course of the response time effect reported above, we carried out analysis on the distribution of RTs. Trials were divided according to trial congruency (congruent or incongruent), rank ordered for each participant and then divided into four bins of equal size. If the number of trials containing correct responses did not divide perfectly by 4, bin sizes were rounded to the nearest whole number. The median RT in each bin for each condition was then calculated. The difference in median RT for congruent and incongruent trials gives an indication of the size of the congruency effect for that part of the RT distribution. These median effect sizes were averaged across participants and are shown in .
To examine whether the size of the congruency effect changed as a function of RT, we conducted a 4 (quartile) × 2 (congruency) repeated measures analysis of variance (ANOVA) on the median RT for each participant in each condition. As expected, there were significant effects of quartile, F(1.05, 25.16) = 140.70, p < .01, with a Greenhouse–Geisser correction employed due to a violation of the sphericity assumption, and of congruency, F(1, 24) = 8.41, p < .01. Importantly, there was a significant interaction between the effects of quartile and congruency, F(1.30, 31.12) = 3.89, p < .05, with Greenhouse–Geisser correction, indicating that the size of the congruency effect changed as a function of RT.
This finding is in line with other recent investigations into the time course of affordance effects (e.g., Cho & Proctor, Citation2010; Tucker & Ellis, Citation2001). Thus, it seems that although object orientation was irrelevant to the task in this experiment, it continued to influence execution of motor responses and shows no evidence of the rapid decay that would be expected to result if responses primed by affordance were suppressed, as shown for typical Simon tasks (e.g., De Jong et al., Citation1994).
Conflict history modulates congruency effects
Next we examined whether the effect of congruency on a given trial was modulated by the congruency of the preceding trial, as it is in traditional “conflict” tasks such as Eriksen flanker (e.g., Gratton et al., 1988), Simon (e.g., Wühr & Ansorge, Citation2005), and Stroop tasks (e.g., Egner & Hirsch, Citation2005). On each trial, it was possible for the target object to be repeated from the immediately preceding trial (approximately 10% of trials). To avoid the possibility that any trial-to-trial effects were produced by repetition priming of the exact stimulus–response link from the immediately preceding trial (see Mayr, Awh, & Laurey, Citation2003), trials in which the same object was repeated, regardless of whether the action afforded by the object was repeated or not, were removed from analysis along with the first trial from each block. However, it should be noted that this does not remove the possibility that there might have been some featural or category overlap from the object presented on the previous trial, which may in turn have activated the same response.
The remaining data were entered into a 2 (previous trial congruency) × 2 (current trial congruency) repeated measures ANOVA to analyse median correct RTs. There was a significant interaction between the effects of current trial and previous trial congruency, F(1, 24) = 7.55, p = .011 (interaction shown in ). The form this interaction takes is consistent with the usual pattern of Gratton effects: The effect of conflict on trial n was greater when trial n – 1 was congruent than when the previous trial was incongruent.
Figure 3 Mean of median response times (RTs) as a function of current trial congruency and previous trial congruency. Results are consistent with the conflict monitoring hypothesis—if the current trial was incongruent, RTs are significantly faster if the immediately preceding trial was also incongruent than if the previous trial was congruent.
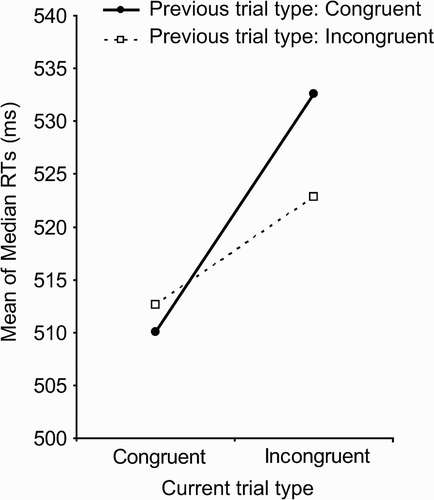
The conflict monitoring hypothesis (e.g., Botvinick et al., Citation2001) predicts that experience of conflict on the previous trial results in an increase in cognitive control in the subsequent trial—even when the same stimulus–response links are not repeated. A one-tailed, repeated measures t test on our data supports this hypothesis: If the current trial is incongruent, RTs are significantly faster if the previous trial was also incongruent compared to if the previous trial was congruent, t(24) = 1.81, p < .05.
Direct evidence for automatic activation of afforded responses
Trials containing responses from the erroneous hand were infrequent, accounting for a little over 5% of trials, and there was no reliable effect of trial congruency on overall error rate (Wilcoxon signed ranks test, Z = –1.29, p > .1).
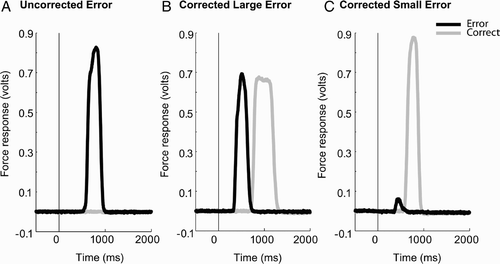
However, grip force measurements potentially allow us to observe some very small errors that might not be detected using all-or-nothing button-press measurements. Examples of individual trials containing errors are shown in .
Figure 4 Examples of responses on individual trials showing errors. shows an error response with the incorrect hand; and show examples of incorrect responses that were followed by a correct response. shows a trial that contains a large erroneous response. shows an example of one of the many trials where the error made was very small and might not have been detected using button-press measures. The stimulus was presented at time zero.
Here, we classified trials containing errors (responses with the incorrect hand) into two types:
| 1. | Uncorrected error trials (accounting for ∼19% of all errors), which contained an above-threshold response made by the incorrect hand and no above-threshold response made by the correct hand. | ||||
| 2. | Corrected error trials (accounting for ∼ 81% of all errors), which contained above-threshold activity for both correct and incorrect hands. Many of these corrected errors showed very small levels of response activity (see , which might not have been sufficient to produce an erroneous button press but are measureable using our force transducers, potentially providing direct evidence for simultaneous coactivation of responses. | ||||
We analysed data from uncorrected and corrected error trials separately. There was no significant effect of trial congruency on the number of uncorrected errors made (Z = –0.42, p > .1). However, corrected errors were significantly more frequent on incongruent trials (mean number of errors = 6) than on congruent trials (mean number of errors = 5; Wilcoxon signed ranks test, Z = –2.25, p = .024). This finding is consistent with the suggestion that object affordances (partially) prime the observer to interact with the object, even when the observer is engaged in a different task.
GENERAL DISCUSSION
We simultaneously recorded continuous measures of force from both hands to investigate the effects of conflict between coactivated responses in a classic object affordance task. Using this continuous measure of response, we found that RTs were faster if the (irrelevant) response afforded by the object was congruent with the response required by the task (. This is consistent with the effect on response time reported in the affordance literature (e.g., Derbyshire et al., Citation2006; Ellis & Tucker, Citation2000; Tucker & Ellis, Citation1998; Vainio et al., Citation2007) and provides further evidence consistent with the view that perceiving an object can partially activate motor plans associated for interacting with that object.
Using continuous grip force measurements we also found that corrected erroneous responses occurred more frequently on incongruent than on congruent trials—that is, when there was conflict between the action afforded by the object and the action required by the task. These findings provide direct evidence that viewing an object activates motor plans appropriate for interacting with that object, even if those motor plans are not relevant for the task. It is likely that small amounts of erroneous grip force activity measured here would not have been detected if participants had responded via a button press, because they would have been of insufficient magnitude to cross the threshold required to be measured. Therefore, grip force measurements might provide a more sensitive means for measuring motor plans activated by object affordances.
Affordance versus other S–R mapping tasks
We set out to investigate possible similarities between the congruency effects shown in a traditional object affordance task and those reported in other S–R mapping tasks. Here we have shown that the effect an afforded action has on responses is modulated by experience of conflict between coactivated responses on the immediately preceding trial. Responses on incongruent trials were reliably faster if they were preceded by another incongruent trial rather than by a congruent trial. This finding cannot be explained by a speeding of responses by repetition of exactly the same S–R linkage from the immediately preceding trial, as trials containing the same object as that in the preceding trial were removed from analysis.
Instead, the effect of trial history reported here is consistent with the view that experience of conflict adjusts levels of control on subsequent trials (Botvinick et al., Citation2001). Furthermore, medial frontal areas that are active when observers look at objects (Grèzes, Tucker, Armony, Ellis, & Passingham, Citation2003) have also been implicated in control over conflict between coactivated responses (see Nachev, Kennard, & Husain, Citation2008). The conflict adaptation effects in object affordance shown here are similar to those reported in other tasks showing automatic sensorimotor activation (e.g., the “Gratton” effects shown in Eriksen flanker and Stroop tasks). In fact, affordance tasks often use a large stimulus set, which make them well suited to investigating trial-to-trial effects while reducing confounds between repeating the exact same S–R linkage (which are often confounded in other tasks, e.g., flanker). The Gratton effects shown here suggest that object affordance demonstrates conflict effects similar to those shown in other stimulus–response mapping tasks and thus perhaps can be integrated into the framework of overlearnt stimulus–response associations.
Finding that object affordance can show analogous trial-to-trial (Gratton) effects to those shown in other S–R mapping tasks (e.g., Eriksen flanker) suggests that the same control mechanisms might be common to both tasks, and that object affordance effects could be integrated into the S–R literature. The evidence for control that is shown in the Gratton effect here might be expected to manifest in RT delta plots as a positive congruency effect that decreases—or even becomes negative—as RTs get longer (see van den Wildenberg et al., Citation2010, for a review). However, this was not the pattern of results reported here. Rather, the RT delta plot showed no evidence that congruency effects reduced as RT increased. In fact, we found that the size of the congruency effect increased as a function of RT.
It is not immediately clear why we would find evidence for cognitive control in the form of a Gratton effect, but no evidence for control in the form of suppressed responses in the delta plot. Whether a decreasing delta plot slope is shown will depend on the relative temporal properties of this suppression. Therefore, one possibility is that responses automatically activated by object affordances are suppressed but that this suppression occurs too late, or grows too slowly, for it to be seen in the delta plot.
Alternatively, Boy, Husain, and Sumner Citation(2010) have recently emphasized a possible distinction between proactive (prestimulus) and reactive (poststimulus) control. According to Boy et al., separate control mechanisms underlie the Gratton effect (proactive) and target–flanker congruency effect (reactive) in an Eriksen flanker task. Thus, in the present experiment, perhaps the (proactive) control mechanisms driving the Gratton effect are present and are distinct from the (reactive) mechanisms that would be expected to produce a negative sloping delta plot, which are absent here.
It should also be noted that positive delta plots have emerged in other affordance tasks (e.g., Tucker & Ellis, Citation2001; see also Phillips & Ward, 2002), and as noted in the introduction, similar flat or positive delta plots have been shown in S–R mapping tasks, which are believed to involve some inhibitory mechanism. For example, positive or flat delta plots have been produced in Eriksen flanker (e.g., Wylie et al., Citation2009) tasks and also in Simon tasks when the mapping between stimulus and response is more complex than a simple left–right button press with the left–right hands (e.g., when response hands are crossed or when stimuli and response are arranged vertically; Wascher et al., Citation2001).
Some investigators (e.g., Wascher et al., Citation2001; Wiegand & Wascher, Citation2005) have suggested that such positive or flat delta plots are produced by conflict between spatial “codes” that are activated for the stimulus and the response, so that responses are speeded when codes confer and are slowed when there is conflict. Cho and Proctor Citation(2010) recently reported positive delta plots for object affordance tasks as well as some Simon tasks where the stimulus did not afford any action. Therefore, Cho and Proctor suggested that increasing RT functions shown in object affordance are due to conflict between the spatial codes activated by action-relevant stimulus properties and the spatial codes for the response required by the task, just as in complex Simon tasks.
What mechanisms drive so-called “affordance” effects?
The nature of the mechanisms producing affordance effects has been a topic of recent debate, and the suggestion that affordance effects might be based on abstract spatial codes rather than activating responses that are limb- or grasp-specific (as originally suggested by Gibson; 1986, see also Greeno, Citation1994; Tucker & Ellis, Citation1998, Citation2001) is not new. Several researchers have now reported reliable within-hand affordance effects (e.g., Cho & Proctor, Citation2010; Vainio, Ellis, & Tucker, Citation2006, Experiment 4), which not be expected if affordances activated limb- or grasp-specific responses. Furthermore, reliable congruency effects have also been shown using responses made with crossed hands (e.g., Phillips & Ward, 2002, Experiment 2), and even when making foot responses (Phillips & Ward, 2002, Experiment 3), which would also not be expected if objects afforded an action that was specific for interacting with that object.
Other authors have rejected the idea that object representations include possibilities for action at all. Anderson, Yamagishi, and Karavia Citation(2002) noted that shifts of attention to the location of objects also generate spatial response codes (the well-known Simon effect, e.g., Simon, Citation1969). They reported that response times were faster whenever that response is spatially congruent with the most visually salient part of the visual stimulus, regardless of whether the stimulus was a graspable object or not. This suggested that participants directed their attention to the most salient part of the objects and that it was congruency between the response and this shift of attention (not the action afforded by the object) that was responsible for the congruency effects reported in affordance tasks.
However, there are at least two important differences between Anderson and colleagues' Citation(2002) experiment and other affordance experiments, which raise the possibility that different effects are being studied: (a) Anderson et al. used line drawings of common objects as stimuli, which may not have afforded actions in the same way as photographs or real objects as used in other affordance experiments; and (b) participants made their response according to the orientation of the stimulus presented—therefore orientation was not task irrelevant as it is in other affordance experiments. Furthermore, reliable congruency effects have now been reported in object affordance paradigms that rely on object size, not orientation, to produce robust effects (e.g., Derbyshire et al., Citation2006; Ellis & Tucker, Citation2000; Ellis, Tucker, Symes, & Vainio, Citation2007; Tucker & Ellis, Citation2001). It is not clear how the “attention directing” hypothesis put forward by Anderson and colleagues can explain object affordance effects such as these.
Of course, these different accounts for what drives object affordance effects need not be mutually exclusive: Perhaps graspable objects are linked to particular actions precisely because observers have a wealth of experience directing their attention to the graspable parts of objects, and where those objects have been linked to those particular actions. As such, the highly overlearnt associations between stimulus and response could be automatically activated when the object is viewed—resulting in apparent “affordance” effects. Further studies are required to examine the relationship between object affordance and conflict between other S–R mappings.
CONCLUSION
In summary, we report that the effect of afforded actions on responses is modulated by previous experience of conflict in much the same way as reported in other S–R paradigms (e.g., flanker), suggesting that object affordance could be considered as a type of highly overlearnt S–R association. Furthermore, grip force measurements allow detection of erroneous responses that could escape detection by conventional button presses. As such, they provide a sensitive means for measuring the effects of simultaneously coactivated motor responses in conflict tasks, including object affordance.
Acknowledgments
The authors wish to thank Clare O'Donoghue and Hadley Pearce for help with data collection and Martin Donovan for technical assistance. This research was supported by The Wellcome Trust and the National Institute for Health Research Cambridge Biomedical Research Centre (NIHR CBRC) at University College London/University College London Hospitals (UCL/UCLH).
Related Research Data
REFERENCES
- Anderson , S. J. , Yamagishi , N. and Karavia , V. 2002 . Attentional processes link perception and action . Proceedings of the Royal Society B , 269 : 1225 – 1232 .
- Botvinick , M. M. , Braver , T. S. , Barch , D. M. , Carter , C. S. and Cohen , J. D. 2001 . Conflict monitoring and cognitive control . Psychological Review , 108 : 624 – 652 .
- Boutsen , L. , Lamberts , K. and Verfaillie , K. 1998 . Recognition times of different views of 56 depth-rotated objects: A note concerning Verfaillie and Boutsen (1995) . Perception & Psychophysics , 60 : 900 – 907 .
- Boy , F. , Husain , M. and Sumner , P. 2010 . Unconscious inhibition separates two forms of cognitive control . Proceedings of the National Academy of Sciences , 107 : 11134 – 11139 .
- Burle , B. , Allain , S. , Vidal , F. and Hasbroucq , T. 2005 . Sequential compatibility effects and cognitive control: Does conflict really matter? . Journal of Experimental Psychology: Human Perception & Performance , 31 : 831 – 837 .
- Cho , D. and Proctor , R. W. 2010 . The object-based Simon effect: Grasping affordance or relative location of the graspable part? . Journal of Experimental Psychology: Human Perception & Performance , 36 : 853 – 861 .
- Coles , M. G. H. , Gratton , G. , Bashore , T. R. , Eriksen , C. W. and Donchin , E. 1985 . A psychophysical investigation of the continuous flow model of human information processing . Journal of Experimental Psychology: Human Perception & Performance , 11 : 529 – 553 .
- De Jong , R. , Liang , C.-C. and Lauber , E. 1994 . Conditional and unconditional automaticity: A dual-process model of effects of spatial stimulus–response correspondence . Journal of Experimental Psychology: Human Perception & Performance , 20 : 731 – 750 .
- Derbyshire , N. , Ellis , R. and Tucker , M. 2006 . The potentiation of two components of the reach-to-grasp action during object categorisation in visual memory . Acta Psychologica , 122 : 74 – 98 .
- Egner , T. and Hirsch , J. 2005 . The neural correlates and functional integration of cognitive control in a Stroop task . NeuroImage , 24 : 539 – 547 .
- Eimer , M. and Schlaghecken , F. 1998 . Effects of masked stimuli on motor activation: Behavioural and electrophysiological evidence . Journal of Experimental Psychology: Human Perception and Performance , 24 : 1737 – 1747 .
- Ellis , R. and Tucker , M. 2000 . Micro-affordance: The potentiation of components of action by seen objects . British Journal of Psychology , 91 : 451 – 471 .
- Ellis , R. , Tucker , M. , Symes , E. and Vainio , L. 2007 . Does selecting one visual object from several require inhibition of actions associated with nonselected objects? . Journal of Experimental Psychology: Human Perception and Performance , 33 : 670 – 691 .
- Eriksen , B. A. and Eriksen , C. W. 1974 . Effects of noise letters upon the identification of a target letter in a nonsearch task . Perception & Psychophysics , 16 : 143 – 149 .
- Eriksen , C. W. , Coles , M. G. H. , Morris , L. R. and O'Hara , W. P. 1985 . An electromyographic examination of response competition . Bulletin of the Psychonomic Society , 23 : 165 – 168 .
- Fournier , L. R. , Scheffers , M. K. , Coles , M. G. H. , Adamson , A. and Abad , E. V. 1997 . The dimensionality of the flanker compatibility effect: A psychophysiological analysis . Psychological Research , 60 : 144 – 155 .
- Gibson , J. J. 1986 . The ecological approach to visual perception , Boston, MA : Houghton Mifflin .
- Gratton , G. , Coles , M. G. H. and Donchin , E. 1992 . Optimizing the use of information: Strategic control of activation of responses . Journal of Experimental Psychology: General , 121 : 480 – 506 .
- Gratton , G. , Coles , M. G. H. , Sirevaag , E. J. , Eriksen , C. W. and Donchin , E. 1988 . Pre- and poststimulus activation of response channels: A psychophysiological analysis . Journal of Experimental Psychology: Human Perception & Performance , 14 : 331 – 344 .
- Greeno , J. G. 1994 . Gibson's affordances . Psychological Review , 101 : 336 – 342 .
- Grèzes , J. and Decety , J. 2002 . Does visual perception of object afford action? Evidence from a neuroimaging study . Neuropsychologia , 40 : 212 – 220 .
- Grèzes , J. , Tucker , M. , Armony , J. , Ellis , R. and Passingham , R. E. 2003 . Objects automatically potentiate action: An fMRI study of implicit processing . European Journal of Neuroscience , 17 : 2735 – 2740 .
- Iani , C. , Baroni , G. , Pellicano , A. and Nicoletti , R. 2011 . On the relationship between affordance and Simon effects: Are the effects really independent? . Journal of Cognitive Psychology , 23 : 121 – 131 .
- Mayr , U. , Awh , E. and Laurey , P. 2003 . Conflict adaptation effects in the absence of executive control . Nature Neuroscience , 6 : 450 – 452 .
- Nachev , P. , Kennard , C. and Husain , M. 2008 . Functional role of the supplementary and pre-supplementary motor areas . Nature Reviews Neuroscience , 9 : 856 – 869 .
- Oldfield , R. C. 1971 . The assessment and analysis of handedness: The Edinburgh Inventory . Neuropsychologia , 9 : 97 – 113 .
- Pellicano , A. , Iani , C. , Borghi , A. M. , Rubichi , S. and Nicoletti , R. 2010 . Simon-like and functional affordance effects with tools: The effects of object perceptual discrimination and object action state . The Quarterly Journal of Experimental Psychology , 63 : 2190 – 2201 .
- Phillips , J. C. and Ward , R. 2002 . S–R correspondence effects of irrelevant visual affordance: Time course and specificity of response activation . Visual Cognition , 9 : 540 – 558 .
- Ridderinkhof , K. R. 2002 . “ Activation and suppression in conflict tasks: Empirical clarification through distributional analyses ” . In Attention & Performance XIX: Common mechanisms in perception and action , Edited by: Prinz , W. and Hommel , B. Oxford, UK : Oxford University Press .
- Simon , J. R. 1969 . Reactions towards the source of stimulation . Journal of Experimental Psychology , 81 : 174 – 176 .
- Symes , E. , Ellis , R. and Tucker , M. 2005 . Dissociating object-based and space-based affordances . Visual Cognition , 12 : 1337 – 1361 .
- Tucker , M. and Ellis , R. 1998 . On the relations between seen objects and components of potential actions . Journal of Experimental Psychology: Human Perception and Performance , 24 : 830 – 846 .
- Tucker , M. and Ellis , R. 2001 . The potentiation of grasp types during visual object categorization . Visual Cognition , 8 : 769 – 800 .
- Vainio , L. , Ellis , R. and Tucker , M. 2006 . Manual asymmetries in visually primed grasping . Experimental Brain Research , 173 : 395 – 406 .
- Vainio , L. , Ellis , R. and Tucker , M. 2007 . The role of visual attention in action priming . The Quarterly Journal of Experimental Psychology , 60 : 241 – 261 .
- van den Wildenberg , W. P. M. , Wylie , S. A. , Forstmann , B. U. , Burle , B. , Hasbroucq , T. and Ridderinkhof , K. R. 2010 . To head or to heed? Beyond the surface of selective action inhibition: A review . Frontiers in Human Neuroscience , 4 : 1 – 13 .
- Verfaillie , K. and Boutsen , L. 1995 . A corpus of 714 full-color images of depth-rotated objects . Perception & Psychophysics , 57 : 925 – 961 .
- Wascher , E. , Schatz , U. , Kuder , T. and Verleger , R. 2001 . Validity and boundary conditions of automatic response activation in the Simon task . Journal of Experimental Psychology: Human Perception & Performance , 27 : 731 – 751 .
- Wiegand , K. and Wascher , E. 2005 . Dynamic aspects of stimulus–response correspondence: Evidence for two mechanisms involved in the Simon effect . Journal of Experimental Psychology: Human Perception & Performance , 31 : 453 – 464 .
- Wühr , P. and Ansorge , U. 2005 . Exploring trial-by-trial modulations of the Simon effect . Quarterly Journal of Experimental Psychology , 58A : 705 – 731 .
- Wylie , S. A. , van den Wildenberg , W. P. M. , Ridderinkhof , K. R. , Bashore , T. R. , Powell , V. D. Manning , C. A. 2009 . The effect of speed–accuracy strategy on response interference control in Parkinson's disease . Neuropsychologia , 47 : 1844 – 1853 .