
ABSTRACT
Processes like gender socialization (the ways in which parents convey information to their children about how girls and boys should behave) often happen unconsciously and might therefore be studied best with neuroscientific measures. We examined whether neural processing of gender-stereotype-congruent and incongruent information is more robustly related to mothers’ gendered socialization of their child than mothers’ implicit and explicit gender stereotypes. To this end, we examined event-related potentials (ERPs) of mothers (N = 35) completing an implicit gender-stereotype task and mothers’ gender stereotypes in relation to observed gendered communication with their child (2–6 years old) in a naturalistic picture-book-reading setting. Increased N2 activity (previously related to attentional processes) to gender stimuli in the implicit gender-stereotype task was associated with mothers’ positive evaluation of similar gendered behaviors and activities in the picture book they read with their child. Increased P300 activity (previously related to attention to unexpected events) to incongruent trials in the gender-stereotype task was associated with a more positive evaluation of congruent versus incongruent pictures. Compared to mothers’ gender stereotypes, neural processing of gendered information was more robustly related to how mothers talk to their children about boys’ and girls’ stereotype-congruent and incongruent behavior, and masculine and feminine activities.
Introduction
Gender is one of the most important organizers of social life, starting from the moment a child is born (Blakemore, Berenbaum, & Liben, Citation2009). In the family context, it shapes how children are parented and talked to, the opportunities they are provided with, and parents’ reactions to certain behaviors, hobbies, interests, and play styles; processes which can be referred to as parental gender socialization (Mesman & Groeneveld, Citation2017). An important way for parents to convey information to their children about how girls and boys are supposed to behave is via gendered communication (Endendijk et al., Citation2014). For example, by making evaluative comments about stereotype-congruent and incongruent behavior (e.g., “That is strange, boys playing with dolls”), parents convey in a subtle way which toys and behaviors are gender-appropriate (Endendijk et al., Citation2014; Friedman, Leaper, & Bigler, Citation2007). It has been suggested that gendered communication often happens unconsciously (Endendijk et al., Citation2014; Mesman & Groeneveld, Citation2017). Consequently, we know little about the underlying processes of gendered communication. It is important to understand the underlying processes, because traditional gendered communication or socialization has been associated with gender-inequality and the development of traditional gender-stereotypes in children (Turner & Gervai, Citation1995) and the development of gender differences in child problem behavior (Endendijk et al., Citation2017). The current study examined whether mothers’ explicit and implicit gender stereotypes, and neural responses to gendered stimuli can predict observed gendered communication with their children.
Explicit and implicit gender stereotypes
Gender stereotypes are widely held beliefs about the characteristics, behaviors, and roles of men and women. The theory of planned behavior predicts that gender stereotypes unconsciously influence our behavior towards men and women (Ajzen, Citation1985). People can hold gender stereotypes both explicitly and implicitly. Explicit stereotypes are overtly expressed ideas that are under high levels of conscious control (Greenwald, Poehlman, Uhlmann, & Banaji, Citation2009) and are most often assessed with self-report questionnaires (Greenwald et al., Citation2009). Implicit stereotypes, on the other hand, are supposedly relatively inaccessible to conscious awareness (Gawronski & Bodenhausen, Citation2006). They are most often assessed with the implicit association test (IAT), that measures the strength of associations between concepts (e.g., men, women, black people, gay people) and attributes (e.g., career, family, angry, good, bad). The task consists of congruent blocks in which participants are requested to sort attributes and concepts in a way that is congruent with stereotypes (e.g., pair men with career words and women with family words), and incongruent blocks in which participants sort attributes and concepts in a way that is incongruent with stereotypes (e.g., pair men with family words and women with career words). The main idea is that making a response is easier when closely associated concepts are sorted with the same response key (i.e., the congruent condition). Because the IAT is based on automatic/fast or habitual responding, it is less prone to social desirability bias than explicit measures of stereotypes. Implicit stereotypes have meta-analytically been found to outperform explicit stereotypes in the prediction of actual behavior, in particular for controversial subjects such as gender and race (Greenwald et al., Citation2009). However, implicit stereotype measures have also been criticized for not only assessing truly implicit/unconscious processes, but also assessing more controlled processes (Hahn, Judd, Hirsh, & Blair, Citation2014; Williams & Themanson, Citation2011).
When we extend this reasoning to a parenting context, one could assume that parents’ implicit gender stereotypes, in particular, are related to how they communicate about gender with their sons and daughters. Parents with implicit stereotypical expectations about boys and girls (e.g., boys should play with cars, girls should play with dolls) indeed were found to make more positive comments in response to children’s stereotype-congruent behavior (e.g., boy playing with cars), compared to stereotype-incongruent behavior (e.g., boy playing with dolls) (Endendijk et al., Citation2014; Friedman et al., Citation2007). However, the available evidence on this regard is scarce and the effect sizes of the associations are small to moderate.
Brain responses to stereotype-congruent and stereotype-incongruent stimuli
Recently, it was discovered that mechanisms underlying stereotyped behavior can be assessed by studying brain responses to stereotype-congruent versus stereotype-incongruent material (Cloutier, Gabrieli, O’Young, & Ambady, Citation2011; Hehman, Ingbretsen, & Freeman, Citation2014). Based on this neuroscientific research, a neural model of implicit stereotypes and behavior was developed involving the anterior cingulate cortex (ACC; conflict monitoring) and dorsolateral prefrontal cortex (dlPFC; regulative control) as important for regulating behavior during tasks in which people have to make speeded responses to stereotype-congruent and incongruent material (for a review see Stanley, Phelps, & Banaji, Citation2008). Especially studies using electro-encephalography (EEG) have the potential to track fast processes underlying implicit stereotyped behavior, due to their millisecond temporal resolution.
Little is known, however, about the degree to which the neural processes underlying stereotyped responding to stereotype-congruent and incongruent material actually predict stereotyped behavior in more naturalistic settings (e.g., gendered parenting). Only one study related individual differences in event-related potentials (ERPs) in a prosocial-behavior IAT to actual donating behavior (Xiao, Zheng, Wang, Cui, & Chen, Citation2015). In this study, people who showed increased N2 activity in response to incongruent trials (associating prosocial words with ‘others’ and non-prosocial words with “self”) compared to congruent trials (associating prosocial words with “self” and non-prosocial words with “others”) on the prosocial IAT, donated more than people who showed increased N2 activity in response to congruent than incongruent trials. In addition, N2 activity during the IAT was a better predictor of donating behavior than implicit prosocial stereotypes. In the current study, we examined whether effects were similar for neural responses indicating gender stereotypes and gendered communication of mothers with their children.
N2 and stereotypes
Several studies already showed that ERPs to stereotype-congruent and incongruent material predict implicit stereotypes as measured with IAT-like tasks. However, few studies specifically examined gender stereotypes. The following ERP components are most often studied in the context of implicit stereotypes: N2/Medial Frontal Negativity (MFN) (e.g., Healy, Boran, & Smeaton, Citation2015; Hilgard, Bartholow, Dickter, & Blanton, Citation2015), and P300 (e.g., Healy et al., Citation2015). However, the direction of effects differs between studies, as well as the components the effects are found on. For example, a negative amplitude deflection measured over medial frontal regions occurring around 200–300 ms, from here on referred to as N2, was found to be larger to trials that are congruent rather than incongruent with the stereotype (Healy et al., Citation2015). This was specifically the case in people with medium stereotype scores, whereas this difference was nonexistent in people with low or high implicit stereotypes (Healy et al., Citation2015).
In contrast, others found the N2 to be larger for trials that are incongruent rather than congruent with the stereotype, in particular for people with strong racial stereotypes (Hilgard et al., Citation2015). This could indicate that stereotype-incongruent trials elicit increased attention, proactive control or conflict monitoring in people with strong racial stereotypes. In yet another study, White people with high racial stereotypes showed larger N2’s to White than to Black faces, while White people with low racial stereotypes showed larger N2’s to Black than toWhite faces (He, Johnson, Dovidio, & McCarthy, Citation2009). It is suggested that the N2 may reflect enhanced attentional processing of in-group targets (Ito & Bartholow, Citation2009; Ito & Urland, Citation2003), which is affected by individual and contextual differences in social identity (Derks, Stedehouder, & Ito, Citation2015). In sum, some study designs show that there is increased N2 activity, as a reflection of increased attention (Ito & Urland, Citation2003), to stereotype-congruent targets or ingroup targets. Others show increased N2 activity, as a reflection of conflict monitoring (West & Bailey, Citation2012), with stereotype-incongruent targets.
P300 and stereotypes
Regarding the P300, a positive amplitude deflection occurring around 250–500 ms (Polich, Citation2007), one study also found an IAT-effect on P300, demonstrating that stereotype strength (i.e., personal connectedness to nature) is associated with P300 amplitude difference in incongruent versus congruent conditions (Healy et al., Citation2015). The P300 is associated with several cognitive processes relevant to IAT-like tasks, such as response selection under difficult conditions (Twomey, Murphy, Kelly, & O’Connell, Citation2015), and attention allocation to stimuli that are negatively valenced, surprising or unexpected (Bartholow & Dickter, Citation2007; Gyurovski, Kubota, Cardenas-Iniguez, & Cloutier, Citation2018; Polich, Citation2007). Therefore, one would expect increased P300 activity to stereotype-incongruent (unexpected, difficult) conditions compared to stereotype-congruent conditions (Bartholow & Dickter, Citation2007), especially in people with strong stereotypes. Also, for people who tend to view deviations from expected gender roles negatively, stereotype-incongruent conditions contain negative valance and could increase P300 activity.
Current study
The current study extended previous work by relating (1) mothers’ brain activity (ERPs) to gender-stereotype congruent and incongruent stimuli, and (2) measures of explicit and implicit gender stereotypes, to (3) mothers’ observed gendered communication with their child during picture book reading. We tested three hypotheses. First, in line with Xiao et al. (Citation2015), ERP amplitude differences (N2, P300) across congruent and incongruent conditions in a gender-IAT-like task would be a better predictor of mothers’ gendered communication than mothers’ explicit or implicit gender stereotypes. These responses that occur within 500ms after stimulus processing are the least likely to be affected by controlled processes that do affect explicit and implicit stereotypes. Second, a larger difference in mothers’ evaluation of stereotype-congruent versus incongruent pictures (positive or negative) would be associated with a larger N2 amplitude difference between congruent and incongruent conditions of the IAT (Healy et al., Citation2015; Hilgard et al., Citation2015). Third, a more positive evaluation of stereotype-congruent compared to incongruent pictures by mothers would be associated with increased P300 amplitudes in incongruent versus congruent conditions of the IAT (Eagly & Diekman, Citation2005; Healy et al., Citation2015).
Methods
Participants
37 mothers with a child aged between 2–6 years old were recruited via the university website, parenting websites, and leaflets handed out in child-care centers. Two participants were excluded due to excessive noise and artifacts (> 25% of trials) in their EEG recordings, resulting in a final sample of 35 mothers. See for the demographic characteristics of the mothers and children. The Ethics Committee of the Faculty of Social and Behavioral Sciences of [Blinded for review] approved the study and it was performed in accordance with the latest version of the Declaration of Helsinki.
Table 1. Demographic characteristics of sample.
Procedure
First, a behavioral observation took place between mother and child. Mother and child were seated in a lab-room with a table and chairs and no further distractions. Mothers were presented with a picture book and were told to look at all the pictures and talk to their child about what they see in the pictures, for a maximum of 10 minutes. After the instructions, the experimenter left the room. The interaction was filmed and coded afterwards.
For the EEG assessment, the mother was taken to another lab-room by a second experimenter. The child stayed with the first experimenter in the behavioral lab. During EEG recording, participants were seated in a soundproof, normal lighted room without windows. They were instructed to minimize eye or body movements during the recording period.
After the EEG assessment, each mother completed several online questionnaires on her child’s behavior and her own stereotypes and parenting practices. Only the explicit gender stereotypes questionnaires (see below) were relevant for the research questions in the current study. Mothers and children respectively received a financial compensation (€15,-) and a small present for their participation.
Measures
For an overview of the measures and concepts used in this study, and the associations that were studied, see .
Figure 1. Overview of the study’s concepts, measures, and associations.
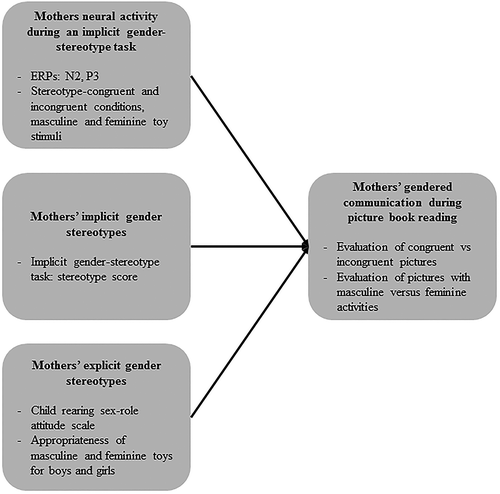
Observed gendered communication
An adapted version of the Gender Stereotypes Picture Book (Endendijk et al., Citation2014) was used to observe mothers’ gendered communication with their children. The book and coding system to rate gendered communication have been validated before (Endendijk et al., Citation2014). See Supplemental Material A for the adaptations made in the current study and an overview of the pictures in the book. Two aspects of gendered communication were assessed as these were most closely related to the stimuli used in the EEG experiment: (1) evaluation of stereotype-congruent (e.g., boys playing with skateboards) versus stereotype-incongruent pictures (e.g., boys dressing up as princesses), (2) evaluation of pictures with masculine activities (e.g., playing with cars) versus pictures with feminine activities (e.g., playing with dolls). Evaluative comments were rated on a range from 1–3 (1 = negative: e.g., “Throwing sand into another child’s face is not nice.”; 2 = neutral: e.g., “They are playing with dolls.”; 3 = positive: e.g., “Look at how much fun those boys are having, playing soccer”). Codes were given per picture and averaged across the picture types. Subsequently, we computed the difference in evaluation of congruent versus incongruent pictures, and feminine activities versus masculine activities (congruent minus incongruent evaluation, feminine activity minus masculine activity evaluation). One undergraduate student was trained by the first author to code the videos for evaluative comments. Agreement between the student and first author was satisfactory (90%, kappa = .80).
Explicit gender stereotypes
The Child Rearing Sex-Role Attitude Scale (CRSRAS, Freeman Citation2007) was used to assess mothers’ explicit stereotypes about boys’ and girls’ behavior and the way they should be parented. The questionnaire consisted of 19 items that were completed on a 5-point scale from 0 (strongly agree) to 4 (strongly disagree). The questionnaire was designed in a way that the items concerned similar statements about boys and girls. For example: “Boys who exhibit ‘sissy’ behavior will never be well adjusted” and “Girls who are ‘tomboys’ will never be well adjusted”. Scores were averaged in one composite score (Cronbach’s alpha = .84), with higher scores indicating stronger stereotypical attitudes about the behavior and parenting of boys and girls.
A second measure of explicit gender stereotypes consisted of 24 stereotypically feminine and masculine toys (also used in th IAT task, see below). For each toy mothers had to indicate whether it was more appropriate for boys, for girls, or equally appropriate for boys and girls. Each score was recoded to reflect a stereotypical response (= 1, masculine toy as more appropriate for boys, feminine toy as more appropriate for girls), an egalitarian response (= 0, toy as equally appropriate for boys and girls), or a counter-stereotypical response (= −1, masculine toy as more appropriate for girls, feminine toy as more appropriate for boys). Recoded scores were averaged in one composite score (Cronbach’s alpha = .95), with positive scores indicating stereotypical ideas about boys’ and girls’ toys, scores around zero indicating egalitarian stereotypes, and negative scores indicating counter-stereotypical ideas.
Implicit gender stereotypes
Mothers completed an IAT-like task to assess implicit gender stereotypes and associated neural responses. The validity of this task to assess implicit gender stereotypes in parents has been demonstrated (Endendijk et al., Citation2013). Participants were asked to divide toys between two children as quickly as possible, by means of pressing one of two keys on the keyboard that were assigned to each child. The two children (full-color) were presented constantly in the left and right hand upper corners on a 19-inch Dell monitor. Each full-color toy was presented in the middle of the screen until the participant hit the response key. Trials were separated by an interstimulus interval of 500 ms. Stimulus presentation, timing, and measurement of behavioral response time and accuracy were controlled by E-Prime (v 2.0) software (Schneider, Eschman, & Zuccolotto, Citation2002).
The task started with a practice block (20 trials) in which red and blue presents had to be divided between two children of unknown gender, followed by two stereotype-congruent blocks and two stereotype-incongruent blocks (68 trials in each block). In the congruent blocks mothers were instructed to assign stereotypically feminine toys (e.g., doll) to a girl and stereotypically masculine toys (e.g., car) to a boy. In the incongruent blocks mothers were instructed to assign stereotypically feminine toys to a boys and stereotypically masculine toys to a girl. To reduce order effects of the presentation of congruent and incongruent blocks (Nosek, Greenwald, & Banaji, Citation2005) mothers were randomly assigned to one of two task orders: (1) congruent-incongruent-congruent-incongruent, (2) incongruent-congruent-incongruent-congruent. In both task orders mothers made each possible switch between congruent and incongruent blocks. Participants were given a rest period between each block of self-determined length (instructions for the next block were also provided), and the entire experiment lasted between 10–15 min, depending on the length of the rest periods. The improved scoring algorithm of Greenwald, Nosek, and Banaji (Citation2003) for the implicit association test was used to determine the level of implicit stereotypes of the parent on the IAT. A high positive score represented more difficulties (e.g., longer reaction times, more errors) pairing masculine toys to girls and feminine toys to boys (stereotype-incongruent) compared to pairing masculine toys to boys and feminine toys to girls (stereotype-congruent), indicating stronger stereotypical ideas about the appropriateness of certain toys for girls and boys.
EEG assessment
Neural assessment
During the implicit gender-stereotype task (see above), EEG was continuously recorded from 32 scalp sites, using BioSemi ActiveTwo Ag-AgCl pin electrodes and hardware (Biosemi, Amsterdam, The Netherlands). The electrodes were placed according to the 10–20 electrode system (Klem, Lüders, Jasper, & Elger, Citation1999), using a nylon electrode cap. EEG signals were amplified with a bandpass of DC-400 Hz by BioSemi ActiveTwo amplifiers and sampled at 2048 Hz. Vertical and horizontal bipolar electrooculographic activity (EOG) was recorded to monitor eye movements using sintered Ag-AgCl electrodes placed above and below the right orbit and near the outer canthus of each eye.
Offline processing of EEG activity was performed with Brain Vision Analyzer software (v2.1). First, data was down-sampled to 256 samples per second, followed by bandpass filtering between 4 and 30 Hz. Data was re-referenced to the average activity of all electrodes. The Gratton & Coles method with Raw Average Subtraction (Gratton, Coles, & Donchin, Citation1983) was used to correct for eye movements and blinks. Epochs time locked to the onset of the toy stimuli were extracted from the cleaned data using a time window of −100 ms to 1000 ms. Artifacts were detected and rejected semi-automatically. Trials with the following characteristics were manually inspected: with a lowest activity below 0.5 uV, a peak-to-peak voltage greater than 100 uV within a 200 ms moving window, or a maximum allowed voltage step that exceeded 50 uV. These trials were deleted when the artifact was visible in multiple electrodes or on one of the electrode sites of interest (isolated artefacts on electrodes outside the region of interest were present on average in < 1% of all trials). A channel was marked as “bad” if noise levels were significantly larger compared to other channels (> 25% of trials rejected due to artifacts exclusively present in “bad” channels). Subsequently, “bad” channels were removed from all preprocessing steps and further analyses. For 13 participants, one or more channels (max. 4) were removed from the datasets (most often Fp1, Fp2, T7, or T8). On average 2% of trials (range: 0–18%) was deleted because of artifacts. The remaining trials were averaged in a grand average waveform, but also separately for each condition (masculine toy-congruent, masculine toy-incongruent, feminine toy-congruent, feminine toy-incongruent). Following Forbes et al. (Citation2012) average waveforms included both correct and incorrect trials, because the improved scoring algorithm to compute a participant’s behavioral stereotype score also included error trials. In addition, we did not exclude any trials on the basis of RTs, because all RTs where within an acceptable range (i.e., 400–1100 ms; see Greenwald et al., Citation2003).
It should be mentioned that no baseline correction was applied, just like in previous ERP studies with IAT-like measures (Forbes et al., Citation2012; Healy et al., Citation2015). The reason for this was the presence of pre-stimulus baseline activity, which might have been due to residual post-movement ERP activity (from the key press) or activity related to expectancy of the upcoming condition. This preparatory activity is inherent to the block-design of IATs, in which stimuli are not randomly presented, but are grouped together in congruent or incongruent blocks. Most importantly, this activity appeared to differentiate between mothers with high and low stereotypes for congruent and incongruent conditions. By baselining, these between-subjects differences in baseline ERP activity would have reduced variance in post stimulus ERP measures (see Supplemental Material B). Therefore, as recommended by others encountering similar problems, we used high-pass filtering at 4 Hz instead of baselining to overcome issues with systematic differences in pre-stimulus baseline activity (Bidet-Caulet et al., Citation2012; Healy et al., Citation2015). However, a consequence of this high-pass filtering is that it is detrimental to late ERP-component time-windows, such as N400 and LPP (Healy et al., Citation2015), which are comprised of lower EEG frequencies in the delta band (0–4 Hz; see Demiralp, Ademoglu, Comerchero, & Polich, Citation2001). Therefore, we focused exclusively on the early N2 and P300 components.
ERPs
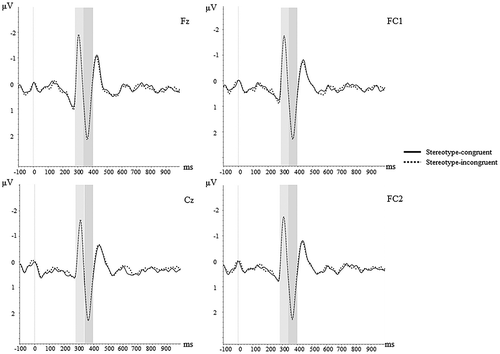
Time windows and electrodes were chosen based on visual inspection of the grand average waveforms. We focused on electrodes with largest amplitudes for each ERP component. All ERP components were quantified as the average activity in a discrete time window following stimulus presentation in the grand average waveform. Specifically, N2 was quantified post-stimulus from 285–340 ms (Fz, Cz, FC1, FC2) and P300 from 340–400 ms (Fz, Cz, FC1, FC2) (see ). These time windows and electrodes were similar to previous IAT-ERP studies (Healy et al., Citation2015; Williams & Themanson, Citation2011). Average activity in these time windows was exported for statistical analysis in SPSS.
Figure 2. N2 and P3 activity to stereotype-congruent and incongruent conditions.
Note. Shaded area’s in figure represent ERP time windows of interest: N2 (285–340ms), P300 (340–400ms)
Analyses
Behavioral data
Implicit stereotype data (RT, accuracy) were submitted to a 2 (stimulus: masculine toy, feminine toy) × 2 (congruence: congruent, incongruent) × 2 (order: congruent first, incongruent first) mixed model ANOVA. Gendered communication about the different picture types was compared in repeated-measures ANOVAs with picture type (congruent vs incongruent, or masculine activity vs feminine activity) as within subject factor. Correlations were computed between mothers’ gendered communication and gender stereotypes.
EEG data
Analyses were performed separately for each ERP component. Mean ERP amplitude values were submitted to a 2 (Stimulus: masculine toy, feminine toy) × 2 (Congruence: congruent, incongruent) × 4 (Electrode: Fz, Cz, FC1, FC2) repeated-measures ANOVA to examine differences in N2 and P300 activity between stimulus types and congruence conditions.
Behavioral and EEG data
Regression analyses were conducted to examine whether gendered communication was best predicted by explicit stereotypes, implicit stereotypes, or ERP amplitudes during the implicit-gender-stereotype task. Therefore, difference scores were computed for the difference in ERP amplitudes (N2, P300) between congruent and incongruent conditions, and between masculine and feminine toy stimuli. We entered ERP difference scores between congruent and incongruent conditions, together with implicit and explicit stereotypes, in analyses predicting mothers’ evaluation of congruent versus incongruent pictures. In an explorative way, we also examined N2/P300 amplitude differences to masculine and feminine toy stimuli in relation to mothers’ evaluation of masculine and feminine activities in the picture book. Therefore, we entered ERP difference scores between feminine and masculine toy stimuli, together with implicit and explicit stereotypes, in analyses predicting mothers’ evaluation of pictures with masculine versus feminine activities. Scatterplots were presented for significant predictors. In all analyses, we controlled for child sex and implicit-gender-stereotype task order (congruent first, incongruent first). However, results were similar with or without the inclusion of the covariates.
Results
Behavioral data: gender stereotypes and observed gendered communication
shows descriptive statistics of behavioral data.
Table 2. Descriptive statistics for behavioral data regarding gender stereotypes and gendered communication.
Implicit gender stereotypes
On average participants performed at 97% accuracy in congruent trials and at 96% accuracy in incongruent trials. RTs were significantly longer to incongruent trials than to congruent trials, F(1, 33) = 30.85, p < .01, partial η2 = .48. No other significant effects on the IAT were observed.
Gendered communication
Mothers were not significantly more positive to their children about stereotype-congruent pictures in the picture book than about stereotype incongruent pictures, F(1, 34) = 1.97, p = .17. There was also no difference in evaluation of pictures with masculine or feminine activities, F(1, 34) = 1.49, p = .23.
Associations between gender stereotypes and gendered communication
Mothers’ implicit stereotypes about gender-typed toys were not associated with gendered communication with their children (all r < .14, p > .43). The same was true for mothers’ explicit gender stereotypes (CRSRAS: all r < .27, p > .11; toy ratings: all r < .09, p > .61). Explicit gender stereotypes were also unrelated to implicit gender stereotypes (all r < .17, p > .33).
EEG data
Overall, there were no significant differences in N2 activity between stimulus types (masculine toy, feminine toy; F (1, 32) = 0.18, p = .67, partial η2 < .01) and congruence conditions (congruent, incongruent; F (1, 32) = 0.03, p = .87, partial η2 < .01). Similarly, on average P300 activity did not differ between congruence conditions (F (1, 32) = 0.10, p = .75, partial η2 < .03). P300 activity did differ between stimulus types (F (1, 32) = 5.27, p < .05, partial η2 = .14), with larger activity in response to feminine toys (M = 1.68, SD = 1.16) compared to masculine toys (M = 1.47, SD = 0.95).
Associations between observed gendered communication, explicit and implicit gender stereotypes, and EEG data
Mothers’ evaluation of congruent vs incongruent pictures
Neither mothers’ explicit gender stereotypes, nor mothers’ implicit gender stereotypes,Footnote1 significantly predicted mothers’ evaluation of congruent versus incongruent pictures while talking to their children about the picture book (all β < .21, p > .30). Difference in N2 activity between congruent and incongruent conditions in the gender-stereotype task was, however, significantly associated with mothers’ evaluation of congruent versus incongruent pictures in the picture book (β = −.48, SE = .17, p < .05, R2 = .22). See ) for a scatterplot of this association. A positive difference between N2 on congruent versus incongruent conditions in the gender-stereotype task (i.e., N2 incongruent > N2 congruent) was associated with mothers’ more positive evaluation of incongruent than congruent pictures. A negative difference between N2 on congruent versus incongruent conditions (i.e., N2 congruent > N2 incongruent) was associated with mothers’ more positive evaluation of congruent than incongruent pictures.
Figure 3. Scatterplots of the associations between mothers’ evaluation of congruent and incongruent pictures and difference in N2 (a) or P3 (b) between congruent and incongruent conditions.
Note. Negative scores on the y-axis of Figure (a) represent a larger N2 to the congruent condition compared to the incongruent condition, whereas positive scores represent a larger N2 to the incongruent condition compared to the congruent condition. Negative scores on the y-axis of Figure (b) represent a larger P3 to the incongruent condition compared to the congruent condition, whereas positive scores represent a larger P3 to the congruent condition compared to the incongruent condition. Negative scores on the x-axis (a & b) represent a more positive evaluation of incongruent pictures compared to congruent pictures, whereas positive scores represent a more positive evaluation of congruent pictures compared to incongruent pictures. In figure (b) solid black dot is an outlier, and dashed line represents regression line with exclusion of the outlier.ERPs were measured during a gender-stereotype task with stereotype congruent and incongruent conditions. Mothers evaluation of pictures was observed during picture-book reading with their own children.
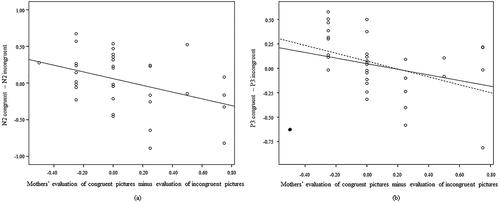
The difference in P300 activity between congruent and incongruent conditions in the gender-stereotype task was near significantly associated with mothers’ evaluation of congruent versus incongruent pictures in the picture book (β = −.34, SE = .18, p = .07, R2 = .16). See ) for a scatterplot of this association. One bivariate outlier could be identified in the lower left corner. Excluding this outlier led to a significant association between P300 activity in the gender-stereotype task and mothers’ evaluation of congruent versus incongruent pictures in the picture book (β = −.51, SE = .17, p < .01, R2 = .29). A positive difference between P300 on congruent versus incongruent conditions in the gender-stereotype task (i.e., P300 congruent > P300 incongruent) was associated with mothers’ more positive evaluation of incongruent than congruent pictures. A negative difference between P300 on congruent versus incongruent conditions (i.e., P300 incongruent > P300 congruent) was associated with mothers’ more positive evaluation of congruent than incongruent pictures.
Mothers’ evaluation of feminine vs masculine pictures
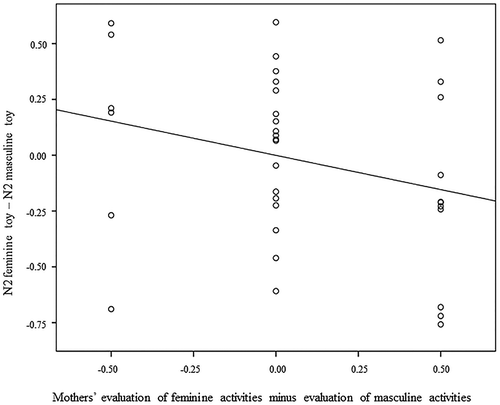
Neither mothers’ explicit nor implicit gender stereotypes, significantly predicted mothers’ evaluation of feminine versus masculine pictures while talking to their children about the picture book (all β < .29, p > .09). Interestingly, child sex was significantly associated with mothers’ evaluation of feminine versus masculine pictures (β = −.42, SE = .10, p < .01). Mothers of girls were more positive about feminine pictures compared to masculine pictures, whereas mothers of boys were more positive about masculine pictures compared to feminine pictures. The difference in N2 activity between feminine and masculine toy stimuli in the gender-stereotype task was significantly associated with mothers’ evaluation of feminine versus masculine pictures in the picture book (β = −.38, SE = .13, p < .05, R2 = .46). See for a scatterplot of this association. A positive difference between N2 for feminine versus masculine toy stimuli (i.e., N2 masculine > N2 feminine) was associated with mothers’ more positive evaluation of pictures with masculine activities compared to feminine activities. A negative difference between N2 on feminine versus masculine toy stimuli (i.e., N2 feminine > N2 masculine) was associated with mothers’ more positive evaluation of pictures with feminine activities compared to masculine activities. P300 activity in the gender-stereotype task was not significantly associated with mothers’ evaluation of masculine and feminine pictures (β < −.09, p = .57).
Figure 4. A scatterplot of the association between mothers’ evaluation of pictures with feminine and masculine activities and N2 difference between feminine and masculine toy stimuli.
Note. Negative scores on the y-axis represent a larger N2 to the feminine toys compared to the masculine toys, whereas positive scores represent a larger N2 to the masculine toys compared to the feminine toys. Negative scores on the x-axis represent a more positive evaluation of pictures with masculine activities compared to pictures with feminine activities, whereas positive scores represent a more positive evaluation of pictures with feminine activities compared to pictures with masculine activities. ERPs were measured during a gender-stereotype task with stereotype masculine and feminine toy stimuli. Mothers evaluation of pictures was observed during picture-book reading with their own children.
Discussion
This study examined whether mothers’ explicit and implicit gender stereotypes, and neural responses to gendered stimuli could predict observed gendered communication with their children. Several associations were found between ERP amplitude differences across congruent and incongruent conditions and toy stimuli in an implicit gender-stereotype task and mothers’ gendered communication with their own child. However, no associations were found between mothers’ implicit or explicit gender stereotypes with gendered communication. These findings indicate that neuroscientific measures were better able to discover underlying mechanisms of an unconscious process, such as gendered communication, than implicit and explicit gender-stereotype measures. This provides an important contribution for research on parenting by indicating that a neuroscientific approach is promising for unraveling unconscious processes underlying the way parents communicate stereotypes to children. This finding also adds to the ecological validity of neuroscientific measures in predicting actual social behavior in a naturalistic setting.
The lack of association between gender stereotypes and gendered communication could be due to a lack of power, as associations between implicit gender stereotypes and gendered communication have been found in larger samples (e.g., Endendijk et al., Citation2014). However, effect sizes for the interactions between mothers’ gendered communication and ERPs during the gender-stereotype task could be considered large (Richardson, Citation2011), while previous associations between gender stereotypes and gender socialization were small to moderate (e.g., Endendijk et al., Citation2014).
Specifically, increased N2 activity to trials that are congruent rather than incongruent with the stereotype on the gender-stereotype task was associated with a more positive evaluation of stereotype-congruent pictures versus stereotype-incongruent pictures in the book mothers read with their child. As N2 activity has been associated with attentional processing (Ito & Urland, Citation2003), our finding could indicate that mothers pay more attention to gendered behavior/activities (either congruent or incongruent with gender stereotypes) which they evaluate positively. This idea is strengthened by our explorative finding that increased N2 activity to masculine toys versus feminine toys was associated with a more positive evaluation by mothers of pictures with masculine activities versus pictures with feminine activities. Similarly, He et al. (Citation2009), demonstrated that in-group targets elicited a larger N2 for people who associate pictures of in-group members strongly with positive words, while for people who associate pictures of outgroup members strongly with positive words, outgroup targets elicited a larger N2. Thus, mothers’ positive evaluations about a group of stimuli (e.g., people, objects) are reflected in mothers’ early/automatic increased attentional processing of targets from this group (Cacioppo, Crites, Berntson, & Coles, Citation1993). These findings add to the neuroscientific literature that N2 activity can be seen as a reflection of early attentional processing, which is modulated by individual differences in gender stereotypes or attitudes.
Our findings regarding the N2 amplitude differences on the congruent and incongruent conditions of the gender stereotype task were partly in line with findings from the study by Healy et al. (Citation2015). They found a larger N2 with congruent trials than with incongruent trials, specifically in people with medium stereotype scores, but no difference in N2 for people with low or high implicit stereotypes. Differences between the current study and the study by Healy et al. (Citation2015) might be attributable to the fact that we uniquely linked ERPs to observed gender-stereotyped behavior. Healy et al. (Citation2015) studied associations with behavioral stereotype scores that were assessed with the same task in which the ERPs were assessed. In addition, Healy et al. (Citation2015) examined a different stereotype domain than the current study (i.e., personal connectedness to nature). Further, in the Healy et al. (Citation2015) study no people with counter-stereotypical attitudes were included, whereas the current study did include mothers who communicated with their child in a counter-stereotypical way (i.e., evaluating stereotype-incongruent pictures more positively than stereotype-congruent pictures). Associations between neural responses to stereotype-congruent and incongruent material with stereotyped behavior/responding might be more linear when the entire spectrum of stereotyped behavior is examined.
Our N2 findings were not in line with Hilgard et al. (Citation2015) and Xiao et al. (Citation2015), who found increased N2 activity in response to incongruent targets in people with strong stereotypes or in people who showed stereotyped behavior. Hilgard et al. (Citation2015) and Xiao et al. (Citation2015) focused on different stereotype domains than the current study (i.e., racial stereotypes, prosocial stereotypes about self versus others), which might explain the differences. However, N2 activity has also been associated with conflict monitoring processes (West & Bailey, Citation2012). So, it is possible that the increased N2 activity found by Hilgard et al. (Citation2015) and Xiao et al. (Citation2015) might reflect increased conflict monitoring, elicited by a discrepancy between a person’s own stereotypes/behavior and the stereotype-incongruent material they were presented with. Indeed, both studies examined negative amplitude deflections between 250–400 ms, which could be regarded as MFN activity, reflecting conflict monitoring, instead of N2 activity (West & Bailey, Citation2012).
Regarding P300 activity, we found increased P300 amplitudes to incongruent compared to congruent conditions of the gender-stereotype task were associated with mothers’ more positive evaluation of congruent versus incongruent pictures. As P300 is associated with attentional focus on unexpected items (Polich, Citation2007) or negatively valanced stimuli (Duval, Moser, Huppert, & Simons, Citation2013; Gyurovski et al., Citation2018) the direction of the association was as expected, and in line with previous studies showing that expectancy-violating stimuli elicited larger P300 than expectancy-consistent stimuli (Bartholow & Dickter, Citation2007). Mothers who showed increased P300 activity to gender-stereotype-incongruent information might unconsciously consider stereotype-incongruent behavior of boys and girls as unexpected or inappropriate. Further, as people tend to view deviations from expected gender roles negatively (Eagly & Diekman, Citation2005) it is not surprising that these mothers evaluated pictures with gender-stereotypical behaviors and activities more positive than pictures with behaviors and activities that are not consistent with gender stereotypes. The increased P300 amplitudes we found for negatively valanced stimuli could also indicate increased encoding strength (Karis, Fabiani, & Donchin, Citation1984). We cannot draw conclusions about which process (e.g., response selection, encoding) is involved in gendered communication. However, P300 activity does seem to be modulated by individual differences in gender-stereotyped evaluations.
Our results regarding P300 should be interpreted with caution, because the high-pass filtering of our EEG data, may have affected one aspect of the P300 component, P3b activity, more than P3a activity. P3a is partly comprised of theta band(4–8Hz) activity and is generated by the activation pattern of the ACC and related frontal structures (Polich, Citation2007). P3b is primarily comprised of lower-frequency activity (0–4Hz) (Demiralp et al., Citation2001) and is generated by temporal-parietal regions. This might explain why P300 activity in the current study was primarily visible in frontocentral electrodes.
The cross-over associations we found between the ERP differences in the gender-stereotype task and the difference in mothers’ evaluation of the pictures types can explain why we did not find main effects of congruence condition or stimulus type on N2/P300 activity. For example, by analyzing mothers who evaluated congruent pictures more positive together with mothers who evaluated incongruent pictures more positive, between-group variation in ERP differences between conditions of the gender-stereotype task cancelled each other out. This highlights an important point for neuroscientific studies on IAT-like tasks. Even if in the whole sample no main differences in ERPs are found between the conditions of the IAT, significant associations can still be present between stereotypes/stereotyped behavior and ERP differences in the conditions of the IAT. Thus, it is important to always take into account individual differences in stereotypes or stereotyped behavior and to see how they relate to neural responses.
Our findings can be interpreted in light of the iterative-reprocessing model (Cunningham & Zelazo, Citation2007), in which a gender-related stimulus elicits evaluative processes (1), relying on pre-existing gender attitudes (2), resulting in an actual evaluation of the stimulus (3), and gender-related behavior (4). Whether this process is conscious or unconscious depends on the number of iterations of the first 3 steps of the model. ‘Implicit’ evaluations have fewer iterations and recruit fewer processes than “explicit” evaluations. In the current study, implicit iterative processing of gender-related stimuli was most likely, given the early nature of the ERPs elicited by the gender-stereotype congruent and incongruent stimuli. Also, the gender-related behavior assessed was of an implicit nature, because making evaluative comments regarding gender towards children often happens unconsciously (Endendijk et al., Citation2014; Gelman, Taylor, Nguyen, Leaper, & Bigler, Citation2004).
The results need to be interpreted while considering the limitations of this study. First, this study had a correlational design. Therefore, no firm conclusions could be drawn about whether ERP activity in an implicit gender-stereotype task predicts mothers’ gender stereotypes and the evaluation of boys’ and girls’ stereotype-congruent and incongruent behavior, or the other way around. Second, because of the way we had to preprocess the EEG data due to the presence of pre-stimulus ERP activity (i.e., high-pass filtering instead of standard baselining), it was only possible to reliably examine early ERPs. However, later ERP components like N400 and LPP have also been associated with performance on implicit stereotype tasks (Forbes et al., Citation2012; Williams & Themanson, Citation2011). Third, because of our modest sample size we were not able to examine differences between mothers with a son and mothers with a daughter. A direction for future research is to examine predictive relationships between functional brain activity in response to stereotype-congruent versus stereotype-incongruent material and observed gendered parenting with boys and girls. Such studies could increase our understanding of specific brain areas underlying the differential treatment of boys and girls by parents. Relatedly, future studies could examine neurobiological underpinnings of several aspects of gendered parenting, next to gendered communication, such as parental modeling of gender roles. These studies can test whether neural processing of gendered stimuli more strongly predicts one aspect of gendered parenting over other aspects.
Conclusion
This study demonstrated that mothers showed enhanced early neural processing of gendered stimuli (congruent, incongruent, masculine toys, feminine toys) they had a positive attitude about. This bias in neural processing was associated with how positive mothers talked to their children about boys’ and girls’ stereotype-congruent and incongruent behavior, and masculine and feminine activities. Further, mothers’ more negative evaluation of boys’ and girls’ stereotype-incongruent behavior compared to stereotype-congruent behavior, was associated with enhanced P300 activity. This could reflect attention allocation to situations that are unexpected based on the gender stereotype (i.e., boy with feminine toy, girl with masculine toy). Most importantly, mothers’ relatively automatic neural processing of gendered stimuli was more robustly associated with mothers’ gendered communication with their children than mothers’ implicit or explicit gender stereotypes. These findings suggest gendered communication is indeed an unconscious process that can be predicted by a person’s attention allocation to gendered stimuli. Therefore, it is particularly important to increase mothers’ awareness of their own gendered attentional biases and the consequences of these attentional processes for their parenting behavior with their sons and daughters.
Supplemental materials.docx
Download MS Word (352.6 KB)Disclosure statement
No potential conflict of interest was reported by the authors.
Supplemental data
Supplemental data for this article can be accessed here.
Additional information
Funding
Notes
1 Although not the focus of the current study, associations between implicit gender stereotypes on the implicit-gender-stereotype task and differences in ERPs between congruent and incongruent conditions of this task can be found in Supplemental Material C.
References
- Ajzen, I. (1985). From intentions to actions: A theory of planned behavior. In J. Kuhl & J. Beckmann (Eds.), Action control: From cognition to behavior (pp. 11–39). New York: Springer.
- Bartholow, B. D., & Dickter, C. L. (2007). Social cognitive neuroscience of person perception: A selective review focused on the event-related brain potential. In E. Harmon-Jones & P. Winkielman (Eds.), Social neuroscience: Integrating biological and psychological explanations of social behavior (pp. 376–400). New York: Guilford Press.
- Bidet‐Caulet, A., Barbe, P.-G., Roux, S., Viswanath, H., Barthélémy, C., Bruneau, N., … Bonnet-Brilhault, F. (2012). Dynamics of anticipatory mechanisms during predictive context processing. European Journal of Neuroscience, 36, 2996–3004.
- Blakemore, J. E. O., Berenbaum, S. A., & Liben, L. S. (2009). Gender development. New York: Psychology Press.
- Cacioppo, J. T., Crites, S. L., Jr, Berntson, G. G., & Coles, M. (1993). If attitudes affect how stimuli are processed, should they not affect the event-related brain potential? Psychological Science, 4, 108–112.
- Cloutier, J., Gabrieli, J. D., O’Young, D., & Ambady, N. (2011). An fMRI study of violations of social expectations: When people are not who we expect them to be. NeuroImage, 57, 583–588.
- Cunningham, W. A., & Zelazo, P. D. (2007). Attitudes and evaluations: A social cognitive neuroscience perspective. Trends in Cognitive Sciences, 11, 97–104.
- Demiralp, T., Ademoglu, A., Comerchero, M., & Polich, J. (2001). Wavelet analysis of P3a and P3b. Brain Topography, 13, 251–267.
- Derks, B., Stedehouder, J., & Ito, T. A. (2015). Social identity modifies face perception: An ERP study of social categorization. Social Cognitive and Affective Neuroscience, 10, 672–679.
- Duval, E. R., Moser, J. S., Huppert, J. D., & Simons, R. F. (2013). What’s in a face? Journal of Psychophysiology, 27, 27–38.
- Eagly, A. H., & Diekman, A. B. (2005). What is the problem? Prejudice as an attitude-in-context. In J. F. Dovidio, P. Glick, & L. Rudman (Eds.), On the nature of prejudice: Fifty years after Allport (pp. 19–35). Malden, MA: Blackwell.
- Endendijk, J. J., Groeneveld, M. G., Mesman, J., Van der Pol, L. D., Van Berkel, S. R., Hallers-Haalboom, E. T., & Bakermans-Kranenburg, M. J. (2017). Gender differences in child aggression: Relations with gender-differentiated parenting and parents’ gender-role stereotypes. Child Development, 88, 299–316.
- Endendijk, J. J., Groeneveld, M. G., Van Berkel, S. R., Hallers-Haalboom, E. T., Mesman, J., & Bakermans-Kranenburg, M. J. (2013). Gender stereotypes in the family context: Mothers, fathers, and siblings. Sex Roles, 68, 577–590.
- Endendijk, J. J., Groeneveld, M. G., Van der Pol, L. D., Van Berkel, S. R., Hallers-Haalboom, E. T., Mesman, J., & Bakermans-Kranenburg, M. J. (2014). Boys don’t play with dolls: Mothers’ and fathers’ gender talk during picture book reading. Parenting: Science and Practice, 14, 141–161.
- Forbes, C. E., Cameron, K. A., Grafman, J., Barbey, A., Solomon, J., Ritter, W., & Ruchkin, D. S. (2012). Identifying temporal and causal contributions of neural processes underlying the Implicit Association Test (IAT). Frontiers in Human Neuroscience, 6, 320.
- Freeman, N. K. (2007). Preschoolers’ perceptions of gender appropriate toys and their parents’ beliefs about genderized behaviors: miscommunication, mixed messages, or hidden truths?. Early Childhood Education Journal, 34(5), 357-366.
- Friedman, C. K., Leaper, C., & Bigler, R. S. (2007). Do mothers’ gender-related attitudes or comments predict young children’s gender beliefs? Parenting: Science and Practice, 7, 357–366.
- Gawronski, B., & Bodenhausen, G. V. (2006). Associative and propositional processes in evaluation: an integrative review of implicit and explicit attitude change. Psychological Bulletin, 132, 692–731.
- Gelman, S. A., Taylor, M. G., Nguyen, S. P., Leaper, C., & Bigler, R. S. (2004). Mother-child conversations about gender: Understanding the acquisition of essentialist beliefs. Monographs of the Society for Research in Child Development, 69(1). doi:10.1111/j.1540-5834.2004.06901002.x
- Gratton, G., Coles, M. G., & Donchin, E. (1983). A new method for off-line removal of ocular artifact. Electroencephalography and Clinical Neurophysiology, 55, 468–484.
- Greenwald, A. G., Nosek, B. A., & Banaji, M. R. (2003). Understanding and using the Implicit Association Test: I. An improved scoring algorithm. Journal of Personality and Social Psychology, 85, 197–216.
- Greenwald, A. G., Poehlman, T. A., Uhlmann, E. L., & Banaji, M. R. (2009). Understanding and using the Implicit Association Test: III. Meta-analysis of predictive validity. Journal of Personality and Social Psychology, 97, 17–41.
- Gyurovski, I., Kubota, J., Cardenas-Iniguez, C., & Cloutier, J. (2018). Social status level and dimension interactively influence person evaluations indexed by p300s. Social Neuroscience, 13(3), 333–345.
- Hahn, A., Judd, C. M., Hirsh, H. K., & Blair, I. V. (2014). Awareness of implicit attitudes. Journal of Experimental Psychology: General, 143, 1369–1392.
- He, Y., Johnson, M. K., Dovidio, J. F., & McCarthy, G. (2009). The relation between race-related implicit associations and scalp-recorded neural activity evoked by faces from different races. Social Neuroscience, 4, 426–442.
- Healy, G. F., Boran, L., & Smeaton, A. F. (2015). Neural patterns of the implicit association test. Frontiers in Human Neuroscience, 9, 605.
- Hehman, E., Ingbretsen, Z. A., & Freeman, J. B. (2014). The neural basis of stereotypic impact on multiple social categorization. Neuroimage, 101, 704–711.
- Hilgard, J., Bartholow, B. D., Dickter, C. L., & Blanton, H. (2015). Characterizing switching and congruency effects in the Implicit Association Test as reactive and proactive cognitive control. Social Cognitive and Affective Neuroscience, 10, 381–388.
- Ito, T. A., & Bartholow, B. D. (2009). The neural correlates of race. Trends in Cognitive Sciences, 13, 524–531.
- Ito, T. A., & Urland, G. R. (2003). Race and gender on the brain: Electrocortical measures of attention to the race and gender of multiply categorizable individuals. Journal of Personality and Social Psychology, 85, 616–626.
- Karis, D., Fabiani, M., & Donchin, E. (1984). “P300” and memory: Individual differences in the von Restorff effect. Cognitive Psychology, 16, 177–216.
- Klem, G. H., Lüders, H. O., Jasper, H. H., & Elger, C. (1999). The ten-twenty electrode system of the International Federation. Electroencephalogry and Clinical Neurophysiology, 52, 3–6.
- Mesman, J., & Groeneveld, M. G. (2017). Gendered parenting in early childhood: Subtle but unmistakable if you know where to look. Child Development Perspectives. online first publication. doi:10.1111/cdep.12250
- Nosek, B. A., Greenwald, A. G., & Banaji, M. R. (2005). Understanding and using the implicit association test: II. Method variables and construct validity. Personality and Social Psychology Bulletin, 31, 166–180.
- Polich, J. (2007). Updating P300: An integrative theory of P3a and P3b. Clinical Neurophysiology, 118, 2128–2148.
- Richardson, J. T. (2011). Eta squared and partial eta squared as measures of effect size in educational research. Educational Research Review, 6(2), 135–147.
- Schneider, W., Eschman, A., & Zuccolotto, A. (2002). E-Prime user’s guide. Pittsburgh: Psychology Software Tools.
- Stanley, D., Phelps, E., & Banaji, M. (2008). The neural basis of implicit attitudes. Current Directions in Psychological Science, 17, 164–170.
- Turner, P. J., & Gervai, J. (1995). A multidimensional study of gender typing in preschool children and their parents: Personality, attitudes, preferences, behavior, and cultural differences. Developmental Psychology, 31, 759–772.
- Twomey, D. M., Murphy, P. R., Kelly, S. P., & O’Connell, R. G. (2015). The classic P300 encodes a build‐to‐threshold decision variable. European Journal of Neuroscience, 42, 1636–1643.
- West, R., & Bailey, K. (2012). ERP correlates of dual mechanisms of control in the counting Stroop task. Psychophysiology, 49, 1309–1318.
- Williams, J. K., & Themanson, J. R. (2011). Neural correlates of the implicit association test: Evidence for semantic and emotional processing. Social Cognitive and Affective Neuroscience, 6, 468–476.
- Xiao, F., Zheng, Z., Wang, Y., Cui, J., & Chen, Y. (2015). Conflict monitoring and stimulus categorization processes involved in the prosocial attitude implicit association test: Evidence from event-related potentials. Social Neuroscience, 10, 408–417.