
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
This paper deals with a plant–pollinator model with diffusion and time delay effects. By considering the distribution of eigenvalues of the corresponding linearized equation, we first study stability of the positive constant steady-state and existence of spatially homogeneous and spatially inhomogeneous periodic solutions are investigated. We then derive an explicit formula for determining the direction and stability of the Hopf bifurcation by applying the normal form theory and the centre manifold reduction for partial functional differential equations. Finally, we present an example and numerical simulations to illustrate the obtained theoretical results.
1. Introduction
It is believed that the explosive diversification and present-day abundance of flowering plants is due to their co-evolution with animal pollinators, especially insects [Citation13]. The interactions between flowering plants and their insect pollinators remain an important ecological relationship crucial to the maintenance of both natural and agricultural ecosystems [Citation15]. Mathematical modeling plays a useful role in pollination research and various mathematical models have been proposed to study plant–pollinator population dynamics, see Soberon and Del Rio [Citation24], Lundberg and Ingvarsson [Citation19], Jang [Citation14], Neuhauser and Fargione [Citation20], Fishman and Hadany [Citation8], Wang et al. [Citation29], Wang [Citation26], and the references cited therein.
Consumer–resource systems model some biological phenomena and relationships between consumer and resource in the real world. A resource is considered to be a biotic population that helps to maintain the population growth of its consumers, whereas a consumer exploits a resource and then reduces its growth rate. Consumer–resource systems have been extensively studied by many researchers (see Chamberlain and Holland [Citation3], Holland and DeAngelis [Citation11], Li et al. [Citation17], Neuhauser and Fargione [Citation20], Wang and DeAngelis [Citation27], Wang et al. [Citation28]). Bi-directional consumer–resource interactions occur when each species acts as both a consumer and a resource of the other. Uni-directional consumer–resource interactions occur when one acts as a consumer and the other as a material and/or energy resource, but neither acts as both.
Recently, Wang, DeAngelis and Holland [Citation29] derived a plant–pollinator model based on unidirectional interactions between plants and pollinators [Citation11]. Pollinators travel from their nest to a foraging patch, collecting food, flying back to their nests, and unloading food. Interacting with flowers individually, the pollinators remove nectar, contact pollen, and provide pollination service. Therefore, the plants provide food, seeds, nectar, and other resources for the pollinators, while the pollinators have both positive and negative effects on the plants. Let and
represent the population densities of plants and pollinators, respectively. The plant–pollinator model takes the following form:
(1)
(1) where
and
are positive constants. The parameter
is the intrinsic growth rate of the plants and
the self-incompatible degree. Following Fishman and Hadany [Citation8], the positive effect of pollinators on plants is described by the Beddington–DeAngelis functional response
, where the parameter a is the effective equilibrium constant for (undepleted) plant–pollinator interaction, which combines traveling and unloading times spent in central place pollinator foraging, with individual-level plant–pollinator interaction. b denotes the intensity of exploitation competition among pollinators (Pianka [Citation21]). Since a is fixed, the parameter
is regarded as the plants efficiency in translating plant–pollinator interactions into fitness (Beddington [Citation2], DeAngelis et al. [Citation6]) and
is the corresponding value for the pollinators.
denotes the per-capita negative effect of pollinators on plants.
is the per-capita mortality rate of pollinators. Wang et al. [Citation29] studied the globally asymptotically stability of the positive equilibria and demonstrated mechanisms by which interaction outcomes of this system vary with different conditions. In particular, it was shown in [Citation29] that system (Equation1
(1)
(1) ) has no periodic orbits or cycle chains in the positive quadrant.
In order to reflect the dynamical behaviours of models depending on the history, it is necessary to incorporate time delay into the models. Following Adams et al. [Citation1], we assume that there is a time delay in the process when the pollinators translate plant–pollinator interactions into the fitness. Also, as pollinators travel between their nests and foraging patches, we further introduce the spatial diffusion with zero-flux boundary conditions. Thus, the plant–pollinator model with diffusion and time delay effects is described by the following delayed reaction–diffusion system:
(2)
(2) where
denotes the diffusion coefficient of pollinators. Ω is a bounded open domain in
and its boundary
is smooth,
is the Laplacian operator in
ν is the outer normal direction on
, and the homogeneous Neumann boundary conditions reflect the situation where the population cannot move across the boundary of the domain.
Throughout this paper, without of loss of generality, we consider the domain . Thus,
We also assume that
and X is a suitable Hilbert space. For example, we can take
with the inner product
.
The rest of the paper is organized as follows. In Section 2, we consider the corresponding characteristic equation of system (Equation2(2)
(2) ) and give conditions on the stability of the positive constant steady-state and the existence of Hopf bifurcation. In Section 3, by applying the normal form theory and centre manifold reduction of partial functional differential equations (PFDEs) (Wu [Citation30], Faria [Citation7]), an explicit algorithm for determining the direction and stability of the Hopf bifurcation is given. Finally, some numerical simulations are included to support our theoretical predictions in Section 4 and a brief discussion is given in Section 5.
2. Stability and Hopf bifurcation
In this section, we consider the local stability of the positive constant steady-state and the Hopf bifurcation of system (Equation2(2)
(2) ) by regarding the time delay τ as the bifurcation parameter. We assume that
(A1)
(A2)
where
We can prove that, if (A1) or (A2) hold, then system (Equation2(2)
(2) ) has two boundary equilibria
and a unique positive constant steady-state
where
Let Then system (Equation2
(2)
(2) ) can be rewritten as
(3)
(3) The positive equilibrium
of system (Equation2
(2)
(2) ) is transformed into the zero equilibrium of system (Equation3
(3)
(3) ).
Let
By the definition of the above functions, for
define
and
as follow:
in particularly
Obviously, we have
By Taylor expansion, Equation (Equation3
(3)
(3) ) becomes
(4)
(4) Let
and
Then system (Equation4
(4)
(4) ) can be rewritten as an abstract differential equation in the phase space
(5)
(5) where
and
are defined by
and
respectively, for
The linearized system of system (Equation5
(5)
(5) ) at
has the form:
(6)
(6) and its characteristic equation is
(7)
(7) where
and
It is well known that the Laplacian operator Δ on X has eigenvalues
with corresponding eigenfunctions
Clearly,
form a basis of X. Thus, any
can be expanded as Fourier series in the following form:
Therefore, (Equation7
(7)
(7) ) is equivalent to
Hence, we conclude that the characteristic equation (Equation7(7)
(7) ) is equivalent to the following sequence of characteristic equations:
(8)
(8) Set
(9)
(9) Notice that (Equation8
(8)
(8) ) with
is the characteristic equation of the linearization of (Equation2
(2)
(2) ) without delay at the positive equilibrium. Because
so the characteristic equation (Equation8
(8)
(8) ) with
does not have a pair of purely imaginary roots for any
with
. According to the Hopf bifurcation theorem, we obtain the following result.
Theorem 2.1.
Assume that (A1) or (A2) hold. Then system (Equation2(2)
(2) ) without delay cannot undergo a Hopf bifurcation at the positive constant steady-state
.
Lemma 2.2.
Assume that (A1) or (A2) hold. Assume further that . Then
is not a root of
Equation (Equation8
(8)
(8) ) for any
with
.
Proof.
From Equation (Equation9(9)
(9) ), we have
Since
and
, we obtain
for any
which implies that
is not a root of Equation (Equation8
(8)
(8) ) for any
.
Lemma 2.3.
Assume that (A1) or (A2) hold. Assume further that Then all roots of Equation (Equation8
(8)
(8) ) with
have negative real parts for all
and the positive constant steady-state
of Equation (Equation2
(2)
(2) ) with
is locally asymptotically stable.
Proof.
When Equation (Equation9
(9)
(9) ) is equivalent to the following equation:
Let
and
be two roots of the above equation. Then
Since
and
all roots of Equation (Equation8
(8)
(8) ) with
have negative real parts.
Let be a purely imaginary root of Equation (Equation8
(8)
(8) ) for
with
Then we have
Separating the real and imaginary parts in the above equation, we obtain
(10)
(10) which imply that
(11)
(11) i.e.
(12)
(12) Set
, (Equation12
(12)
(12) ) is transformed into
(13)
(13) If
, then Equation (Equation13
(13)
(13) ) has only one positive root which is denoted by
. Hence Equation (Equation12
(12)
(12) ) has only one positive root
. From Equation (Equation10
(10)
(10) ), we know that Equation (Equation8
(8)
(8) ) with
has a pair of purely imaginary roots
when
, where
(14)
(14) with
(15)
(15)
Lemma 2.4.
Assume that (A1) or (A2) hold. Assume further that Then
Therefore,
is a simple root of (Equation8
(8)
(8) ) for
Proof.
Firstly, we have
Then, from
we obtain that
Thus, if
then
Since
we have
which implies that
However,
Hence, we have
This completes the proof.
Lemma 2.5.
Assume that (A1) or (A2) hold. Assume further that Let
be the root of Equation (Equation8
(8)
(8) ) for
satisfying
Then
satisfies the following transversality condition:
Proof.
Differentiating both sides of Equation (Equation8(8)
(8) ) with respect to τ yields
From Equation (Equation10
(10)
(10) ), we have
By inserting the expression of
into the last expression, we obtain that
The proof is complete.
Notice that Equation (Equation8(8)
(8) ) with k=0 is the characteristic equation of the linearization of (Equation2
(2)
(2) ) without diffusion at the positive equilibrium. By Rouché theorem and Lemmas Equation5
(5)
(5) – Equation7
(7)
(7) , we have the following results [Citation22,Citation23] :
Theorem 2.6.
Assume that (A1) or (A2) hold. Assume further that The following statements hold:
If
then all roots of Equation (Equation8
If
If
Furthermore, we can obtain the following results:
Theorem 2.7.
Assume that (A1) or (A2) hold. Assume further that
and
Then Equation (Equation8
(8)
(8) ) with
has a pair of simple purely imaginary roots
and all roots of Equation (Equation8
(8)
(8) ) for any
except
have no zero real parts. Moreover, for
all roots of Equation (Equation8
(8)
(8) ) for any
except
have negative real parts.
Theorem 2.8.
Assume that (A1) or (A2) hold. Assume further that
and
The following statements hold:
If
If
Denote
From Equation (Equation14(14)
(14) ), we have
for any
In the rest of this paper, we assume that
and have the following lemma. The case for
can be discussed in a similar way.
Lemma 2.9.
Let be defined as Equation (Equation14
(14)
(14) ). Assume that (A1) or (A2) hold. Assume further that
and
Then for any
Proof.
From Equation (Equation12(12)
(12) ), we have
where
Simple computation shows that
Notice that
is strictly decreasing in k for
Then we obtain that
is strictly decreasing in k for
From Equation (Equation15
(15)
(15) ), we have
By direct computation, we have
Since
by the fact that
and
, we obtain
That is,
is strictly decreasing in k for
So
is strictly increasing in k for
From Equation (Equation14
(14)
(14) ), if
then
is strictly increasing in k for
.
From the above lemma, we have for any
and
Denote
From the above analysis, we have the following conclusion.
Theorem 2.10.
Assume that (A1) or (A2) hold. Assume further that and
The following statements are true:
If
If
3. Properties of Hopf bifurcations
In this section, we shall study the direction, stability and the period of bifurcating periodic solution by applying the normal form theory and the centre manifold theorem presented in [Citation7,Citation10,Citation30]. Let Normalizing the delay τ in system (Equation4
(4)
(4) ) by the time-scaling
, Equation (Equation4
(4)
(4) ) is transformed into
(16)
(16) Let
and
Then system (Equation16
(16)
(16) ) can be rewritten in the abstract form in the space
as
(17)
(17) where
and
are defined by
respectively, for
with
(18)
(18)
Consider the linear equation
(19)
(19) According to results in Section 3, we know that the origin
is an equilibrium of
Equation (Equation16
(16)
(16) ), and under some conditions, the characteristic equation of (Equation19
(19)
(19) ) has a pair of simple purely imaginary eigenvalues
We now consider the ordinary functional differential equation:
(20)
(20) By the Riesz representation theorem, there exists a
matrix function
whose entries are of bounded variation such that
(21)
(21) for
In fact, we can choose
(22)
(22) Let
denote the infinitesimal generator of the semigroup induced by the solutions of system (Equation20
(20)
(20) ) and
be the formal adjoint of
under the bilinear pairing
(23)
(23) for
From the previous section, we know that
has a pair of simple purely imaginary eigenvalues
Because
and
are a pair of adjoint operators (see Hale [Citation9]),
are also eigenvalues of
Let P and
be the centre subspace, that is, the generalized eigenspace of
and
associated with
respectively. Then
is the adjoint space of P and
Direct computations yield the following results.
Lemma 3.1.
Let
(24)
(24) Then
form a basis of P with
and
form a basis of
with
.
Let and
with
for
and
for
Now we define
and construct a new basis Ψ for
by
Then
where
is the identity matrix. In addition,
where
Let
be defined by
for
Then the centre subspace of linear equation (Equation19
(19)
(19) ) is given by
where
(25)
(25) and we can decompose
as
, in which
denotes the complement subspace of
in C.
Let be the infinitesimal generator induced by the linear system (Equation19
(19)
(19) ), and Equation (Equation17
(17)
(17) ) can be rewritten as the following abstract form:
(26)
(26) where
By the decomposition of C, the solution of Equation (Equation17
(17)
(17) ) can be written as
(27)
(27) where
and
In particular, the solution of (Equation17
(17)
(17) ) on the centre manifold is given by
(28)
(28) Let
and
Then we obtain
Hence, Equation (Equation28
(28)
(28) ) can be transformed into
(29)
(29) where
From Wu [Citation30], z satisfies
(30)
(30) where
(31)
(31) Let
(32)
(32)
(33)
(33) Notice that
Let
Then by computation, we obtain the following quantities:
where
(34)
(34) with
(35)
(35)
(36)
(36) with
(37)
(37) and
So far, we have obtained and
which can be expressed by the parameters of system (Equation2
(2)
(2) ). Hence, we can compute the following quantities:
Thus, we obtain the following results:
Theorem 3.2.
For any critical value we have
4. Numerical simulations
In this section, we present some numerical simulations to illustrate the theoretical analysis for the system (Equation2(2)
(2) ).
Choose the parameter values as follows so that the conditions in Theorem 2.8 are satisfied:
The initial conditions are taken as
Then system (Equation2
(2)
(2) ) becomes
(38)
(38)
By computation, we have
First we choose
and plot the solutions
and
by using the software Matlab in Figure . From the numerical simulations we can see that the solutions of system (Equation38
(38)
(38) ) with
tend asymptotically to the positive equilibrium
Under the same initial values, now we choose
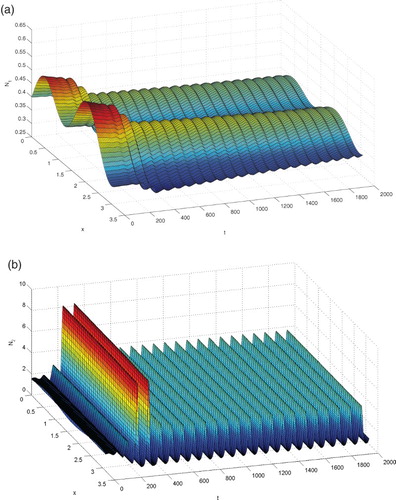
and plot the graphs of
and
in Figure . From Figure , we see that there exists a family temporal periodic solutions, which implies that Hopf bifurcation occurs for system (Equation38
(38)
(38) ) at
Figure 1. The positive equilibrium is asymptotically stable when
.
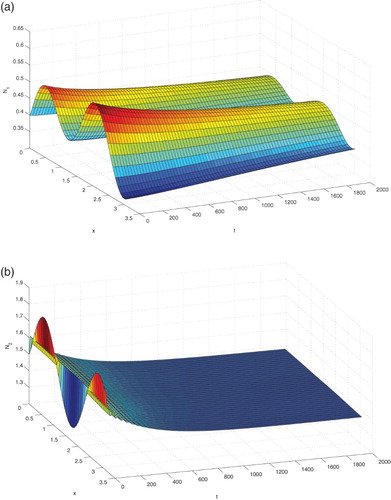
Figure 2. The temporal periodic solutions bifurcated from the equilibrium are stable, where .
5. Discussion
Various mathematical models have been proposed to study plant–pollinator population dynamics, see Soberon and Del Rio [Citation24], Lundberg and Ingvarsson [Citation19], Jang [Citation14], Neuhauser and Fargione [Citation20], Fishman and Hadany [Citation8], Wang et al. [Citation29], and Wang [Citation26]. Most of these models are described by ordinary differential equations. Since pollinators travel between their nests and foraging patches, we believe that reaction–diffusion equations are more suitable to model the interactions between the plants and pollinators. We also assumed that there is a time delay in the process when the pollinators translate plant–pollinator interactions into the fitness and considered a plant–pollinator model with diffusion and time delay effects. As far as we know, there are no results for system (Equation2(2)
(2) ) with diffusion and time delay.
Firstly, by considering the distribution of eigenvalues of the corresponding linearized equation, stability of the positive constant steady-state and existence of spatially homogeneous and spatially inhomogeneous periodic solutions were studied. Secondly, by applying the normal form theory and the centre manifold reduction for partial functional differential equations, an explicit formula for determining the direction and stability of the Hopf bifurcation was given. Finally, to explain the obtained results, numerical simulations were presented.
Our results showed that if and either
or
holds, where
then system (Equation2
(2)
(2) ) has a unique positive constant steady-state
in which
The first inequality
ensures the existence of
and
Recall that
is regarded as the pollinators efficiency in translating plant–pollinator interactions into fitness, a is the effective constant for plant–pollinator interaction, and
is the per-capita mortality rate of pollinators. This inequality means that the efficiency in translating plant–pollinator interactions into fitness of the pollinators must be greater than their mortality rate; otherwise the pollinators even cannot survive.
The inequality in (A1) is equivalent to
which indicates that the ratio of the efficiencies in translating plant–pollinator interactions into fitness of the plants and pollinators is greater than a certain value. In this case, an additional condition
is needed to ensure the existence of
Under the assumption (A2), it requires that
Note that now
so the condition is equivalent to
which, in turn, is equivalent to
The last inequality means that the intrinsic growth rate
of the plants must be large enough compared to the death rates of the plants and pollinators.
We were interested in not only the effect of diffusion but also the effect of delay [Citation4,Citation12,Citation31]. We found that system (Equation2(2)
(2) ) without delay cannot undergo Hopf bifurcations at the positive constant steady-state. But, under certain conditions, system (Equation2
(2)
(2) ) undergoes Hopf bifurcations at the positive constant steady-state under the effect of delay. Recall that
Our results demonstrated that if
then the positive equilibrium
is locally asymptotically stable if the time delay is less than a critical value
unstable when
and a family of periodic solutions bifurcates from
when τ passes through
via Hopf bifurcation. Moreover, the direction, stability and period of the bifurcating periodic solutions can be determined analytically. Notice that Wang et al. [Citation29] showed that the ODE model (Equation2
(2)
(2) ) does not have periodic solutions and Wang et al. [Citation25] proved that the unique positive steady-state solution of a reaction–diffusion plant–pollinator model is a global attractor. Our results thus indicate that the time delay causes bifurcations and induces temporal periodic patterns in the diffusive plant–pollinator model. Such properties have been observed in many delay differential equation models [Citation5,Citation16]. This is similar to the observation in our other work [Citation18] that oscillations occur in age-structured resource–consumer (plant–pollinator) models.
Wang et al. [Citation29] and Wang [Citation26] indeed investigated three species plant–pollinator–robber models. Since the movement of the nectar robbers plays an important role in their invasibility and coexistence of all species, it will be very interesting to study the population dynamics of the three species diffusive plant–pollinator–robber models. We leave this for future consideration.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- V.D. Adams, D.L. DeAngelis, and R.A. Goldstein, Stability analysis of the time delay in a host-parasitoid model, J. Theor. Biol. 83 (1980), pp. 43–62. doi: 10.1016/0022-5193(80)90371-9
- J.R. Beddington, Mutual interference between parasites or predators and its effect on searching efficiency, J. Anim. Ecol. 44 (1975), pp. 331–340. doi: 10.2307/3866
- S.A. Chamberlain and J.N. Holland, Density-mediated, context-dependent consumer–resource interactions between ants and extrafloral nectar plants, Ecology 89 (2008), pp. 1364–1374. doi: 10.1890/07-1139.1
- S. Chen, J. Shi, and J. Wei, Global stability and Hopf bifurcation in a delayed diffusive Leslie–Gower predator–prey system, Inter. J. Bifur. Chaos 22 (2012), pp. 1250061-1-11.
- J.M. Cushing, Integrodifferential Equations and Delay Models in Population Dynamics, Lecture Notes in Biomathematics, Vol. 20, Springer-Verlag, Heidelberg, 1977.
- D.L. DeAngelis, R.A. Goldstein, and R.V. O'Neill, A model for trophic interaction, Ecology 56 (1975), pp. 881–892. doi: 10.2307/1936298
- T. Faria, Normal forms and Hopf bifurcation for partial differential equations with delays, Trans. Amer. Math. Soc. 352 (2000), pp. 2217–2238. doi: 10.1090/S0002-9947-00-02280-7
- M.A. Fishman and L. Hadany, Plant-pollinator population dynamics, Theor. Popul. Biol. 78 (2010), pp. 270–277. doi: 10.1016/j.tpb.2010.08.002
- J.K. Hale, Theory of Functional Differential Equations, Springer-Verlag, Berlin, 1977.
- B.D. Hassard, N.D. Kazarinoff, and Y.-H. Wan, Theory and Application of Hopf Bifurcation, Cambridge University Press, Cambridge, 1981.
- J.N. Holland and D.L. DeAngelis, Consumer–resource theory predicts dynamic transitions between outcomes of interspecific interactions, Ecol. Lett. 12 (2009), pp. 1357–1366. doi: 10.1111/j.1461-0248.2009.01390.x
- G. Hu and W.-T. Li, Hopf bifurcation analysis for a delayed predator–prey system with diffusion effects, Nonlinear Anal. Real World Appl. 11 (2010), pp. 819–826. doi: 10.1016/j.nonrwa.2009.01.027
- S.S. Hu, D.L. Dilcher, D.M. Jarzen, and D.W. Taylor, Early steps of angiosperm pollinator coevolution, Proc. Natl. Acad. Sci. USA 105 (2008), pp. 240–245. doi: 10.1073/pnas.0707989105
- S.R.-J. Jang, Dynamics of herbivore–plant–pollinator models, J. Math. Biol. 44 (2002), pp. 129–149. doi: 10.1007/s002850100117
- C.A. Kearns, D.W. Inouye, and N.M. Waser, Endangered mutualisms: The conservation of plant–pollinator interactions, Annu. Rev. Ecol. Syst. 29 (1998), pp. 83–112. doi: 10.1146/annurev.ecolsys.29.1.83
- Y. Kuang, Delay Differential Equations with Applications in Population Dynamics, Academic Press, New York, 1993.
- X. Li, H. Wang, and Y. Kuang, Global analysis of a stoichiometric producer–grazer model with Holling type functional responses, J. Math. Biol. 63 (2011), pp. 901–932. doi: 10.1007/s00285-010-0392-2
- Z. Liu, P. Magal, and S. Ruan, Oscillations in age-structured models of consumer–resource mutualisms, Discret Contin. Dyn. Syst. B 21 (2016), pp. 537–555. doi: 10.3934/dcdsb.2016.21.537
- S. Lundberg and P. Ingvarsson, Population dynamics of resource limited plants and their pollinators, Theor. Popul. Biol. 54 (1998), pp. 44–49. doi: 10.1006/tpbi.1997.1349
- C. Neuhauser and J.E. Fargione, A mutualism–parasitism continuum model and its application to plant–mycorrhizae interactions, Ecol. Model. 177 (2004), pp. 337–352. doi: 10.1016/j.ecolmodel.2004.02.010
- E.R. Pianka, Evolutionary Ecology, Harper and Row, New York, 1974.
- S. Ruan, Absolute stability, conditional stability and bifurcation in Kolmogorov-type predator–prey systems with discrete delays, Quart. Appl. Math. 59 (2001), pp. 159–173.
- S. Ruan and J. Wei, On the zeros of a third degree exponential polynomial with applications to a delayed model for the control of testosterone secretion, IMA J. Math. Appl. Med. Biol. 18 (2001), pp. 41–52. doi: 10.1093/imammb/18.1.41
- J .M. Soberon and C.M. Del Rio, The dynamics of a plant-pollinator interaction, J. Theor. Biol. 91 (1981), pp. 363–378. doi: 10.1016/0022-5193(81)90238-1
- L. Wang, H. Jiang and Y. Li, Positive steady state solutions of a plant-pollinator model with diffusion, Discret. Contin. Dyn. Syst. B 20 (2015), pp. 1805–1819. doi: 10.3934/dcdsb.2015.20.1805
- Y. Wang, Dynamics of plant-pollinator-robber systems, J. Math. Biol. 66 (2013), pp. 1155–1177. doi: 10.1007/s00285-012-0527-8
- Y. Wang and D.L. DeAngelis, Transitions of interaction outcomes in a uni-directional consumer–resource system, J. Theor. Biol. 208 (2011), pp. 43–49. doi: 10.1016/j.jtbi.2011.03.038
- Y. Wang, D.L. DeAngelis, and J.N. Holland, Uni-directional consumer–resource theory characterizing transitions of interaction outcomes, Ecol. Complex. 8 (2011), pp. 249–257. doi: 10.1016/j.ecocom.2011.04.002
- Y. Wang, D.L. DeAngelis, and J.N. Holland, Uni-direction interation and plant-pollinator-robber coexistence, Bull. Math. Biol. 74 (2012), pp. 2142–2164. doi: 10.1007/s11538-012-9750-0
- J. Wu, Theory and Applications of Partial Functional Differential Equations, Springer-Verlag, New York, 1996.
- W. Zuo and J. Wei, Stability and Hopf bifurcation in a diffusive predator–prey system with delay effect, Nonlinear Anal. Real World Appl. 12 (2011), pp. 1998–2011. doi: 10.1016/j.nonrwa.2010.12.016