
Abstract
The present study was performed to estimate genetic parameters for growth and fur quality traits in Chinese silver blue mink. Recordings of body weight (BW), body length (BL), guard hair length (GL), underwool length (UL) and the rate of guard hair and underwool length from 1678 silver blue minks (280 male and 1398 female) spanning six generations were obtained from Dalian Ming Wei Marten Industry Company. Genetic parameters were estimated using a multi-trait animal model with year and sex as fixed effects. The heritability estimates for BW, BL, GL, UL and RATE were 0.41, 0.53, 0.53, 0.52 and 0.52, respectively. The genetic trends were estimated by regressing breeding value on year. The genetic trends for body weight and body length were close to zero, and positive. The genetic changes for GL, UL and nap were negative and parallel. Our results herein form a practical basis for designing optimal breeding schemes in silver blue mink.
Introduction
The American mink (Neovison vison), a semi-aquatic carnivore originating in North America, is the most widespread animal used in fur production. In the breeding of mink, proper quality of hair coat and body size are the utmost important production traits from the economical point of view (Peura et al. Citation2004a), and the other significant traits include litter size, feed efficiency, etc. A thorough understanding of the genetic variability of these traits in mink is key to genetic enhancements in production performance. Building upon advances in animal and plant breeding, genetic improvements in animal fur have been successfully implemented during the past three decades (Cai et al. Citation2008; Zongyue et al. Citation2011). In particular, advances have been made and are still in progress in the farming of mink (Socha & Kolodziejczyk Citation2006). Silver blue is the first mutant coat colour known in the mink farm, which is also named splatinum and maltese. The first silver blue mink appeared in 1931 on a farm owned by W. Whittingham in Arpin, Wisconsin. The first silver blue mink pattern was introduced in China in the 1980s for fur farming, but mink breeding started in 2002. Recently, many studies on the estimation of genetic parameters and breeding values for productive traits have been published for fur animals (Hansen & Berg Citation2008; Nielsen et al. Citation2012; Peura et al. Citation2004a; Wierzbicki & Jagusiak Citation2006). But, there is a large variability in heritability and correlation estimates among different mink populations, which is most probably caused by natural and artificial selection (Socha & Kolodziejczyk Citation2006). Another problem concerns the definition of traits within populations. The skin length, fur quality, colour type and pelt characters have had a marked influence on the price of mink pelt. There is now a general consensus that body size and fur traits are the most important economic traits in mink pelt. In virtue of selecting and breeding in modern animal production, mature size and fur quality have improved (Zongyue et al. Citation2011).
In the present study, data were collected on the production traits of silver blue mink from six generations. The purpose of this study was to estimate genetic parameters and breeding values for production traits of silver blue mink using Multiple Trait Derivative Free Restricted Maximum Likelihood programme (MTDFREML).
Material and methods
Data collection
Samples were obtained from Dalian Ming Wei Marten Industry Company. The mink has bulky lengthy body, with sedge-shaped head. It has bluish grey colour for the whole body, as well as back and belly. The guard hair is flush, bright and flexible. The under fur is thick, supple and dense. The nose and eyes are lighter than in standard mink. The mink is known to have good size and to be strong with good fecundity. The company with its equipment is well suited to study mink. The kits were randomly selected from different litter with similar age at birth, which were born from 25 April to 5 May. The third time evaluations of body dimensions were done in December. Throughout this period, the kits were fed ad libitum two times a day with a variety of commercially available, nutritionally complete diets, which are formulated for kit growth, at a level designed to maintain optimum body composition. After weaning, minks were kept in individual cages. From mating until pelting, animals were paired, with one male and female full-sib pair in each cage.
Traits measured
The individual body weight (±0.1g), body length (± 0.1 cm), guard hair length (± 0.1 cm), underwool length (± 0.1 cm) and the rate of the two hair types were recorded in December (the third time evaluation) throughout the six generations. The mink was weighed in the morning before feeding. The body length was measured from nose to the root of the tail. The natural unstretched length of guard hair and underwool was measured from skin to the top of hair with a ruler. These traits for each mink were available for 1686 individuals over six generations between 2005 and 2011. The individuals were arranged according to the family pedigree and requirement of MTDFREML.
Statistical analysis of data
Simple descriptive statistics were summarised in Table . Variance components and heritability for each trait were estimated with single trait and two traits animal model by derivative-free REML with a simplex algorithm using the MTDFREML. The genetic correlations between each pair of the above traits were estimated using two traits animal model, and phenotypic correlations between each pair of these traits were estimated by using the Pearson correlation method (SAS 2004). To calculate genetic trends, breeding values across generations were estimated with the Best Linear Unbiased Prediction (BLUP) method using the animal model. Means of animal estimated breeding values were regressed across years to predict annual genetic trends for these traits.
Table 1. Total number (N) of observations, means and standard deviation for the studied traits in the silver blue mink from 2005 to 2011.
Result and discussion
Genetic parameters
The estimated genetic parameters from the single trait animal model is given in Table . Body weight, body length, guard hair length, underwool length and the rate of two hair types are highly heritable, which is consistence with another study (Sengul & Kiraz Citation2005). In this study, the estimation of heritability for body weight and body length was 0.41 and 0.53, which showed moderate-to-high heritability. The value was similar to the estimates of 0.42 ± 0.07 and 0.45 ± 0.07 founded by Hansen and Berg (Citation2008) on standard and mahogany type, and 0.478 for standard and pastel colour type for body size and conformation. Estimates of heritability of guard hair length and underwool length were 0.520 and 0.521, respectively. Genetic parameter for fur quality has been rarely reported, and the assessment for the fur quality is based on different test results, which is commonly expressed in scores or referred to as grading trait (Peura et al. Citation2004a). Heritability estimate of fur quality is usually moderate by this kind of method. Heritability estimates for fur quality was 0.235 in the study by Socha & Kolodziejczyk (Citation2006); between 0.10 and 0.38 for fur quality (estimates related to the gender and colour type); between 0.24 and 0.93 for hair length, between 0.33 and 0.49 for hair density. The measurement method of hair that we used was more accurate by measuring from skin to the top of hair by ruler. The assumption is that transformation of a continuous quantitative trait to a categorical trait leads to smaller heritability because the transformation introduces inaccuracy or poorer resolution in the measurement.
Table 2. Estimates of genetic and phenotypic parameters among growth and fur quality traits in silver blue mink (heritability on diagonal, phenotypic correlations above diagonal and genetic correlations below diagonal).
The results showed that the phenotypic correlations between the rate of two hair lengths and body weight, body length, guard hair length and underwool length were negative (− 0.218, − 0.178, − 0.074, − 0.425), while the other phenotypic correlations were positive (0.298, 0.882, 0.869, 0.806, 0.788, 0.93). The genetic correlation of guard hair length with underwool length and the rate of two hair length was negative (− 0.941, − 0.983), while the rest of genetic correlation were positive (0.983, 0.731, 0.972, 0.981, 0.622, 0.992, 0.641, 0.987).
Estimates of phenotypic correlations between RATE and BW, BL, GL, UL were negative and moderate. The negative correlations between body weight and fur quality of minks have been detected. The high positive phenotypic correlations were found among BW, BL, GL, and are in the general range of values cited in literature (Peura et al. Citation2004a; Kempe et al. Citation2009, Citation2010). Most of the genetic correlations were positive, for example, between the BW and BL (0.983), between BW and GL (0.731), between BW and UL (0.972), respectively, implying that getting bigger size of mink leads to longer pelt and better fur quality. Negative correlations were found between BL and UL (−0.941), between BL and RATE (−0.983). These results are in agreement with the findings by Socha and Kolodziejczyk (Citation2006). The majority of the genetic correlations were negative, for instance, between fur quality and body size (−0.543).
Phenotypic and genetic tendency of growth and fur traits
The phenotypic trends of all traits in male and female animals between 2005 and 2011 are reported in Figure . For the body weight and body length, the phenotypic changes from 2005 to 2010 appear to be flat and then decline (female) or rise (male) until 2011. The phenotypic trends of the other three traits were unchanged during six generations with fluctuations in 2007 and 2008. The annual genetic trends for all traits are given in Figure . For all the traits, the genetic trend from 2005 to 2011 appears flat. The genetic trend of body weight and body length was close to zero, and a bit positive, respectively. For the body length, the curve of genetic trend showed upward trend. The slope of genetic trend for GL was steeper (−0.00395 ± 0.00126 cm/yr; p < .05) than for UL (−0.00258 ± 0.0000846 cm/yr; p < .05) and RATE (−0.0000385 ± 0.00011/yr; p > .05) (Table ).
Figure 1. Phenotypic change tendency of growth and fur trait in silver blue mink. (A) Phenotypic tendency of the body weight. (B) Phenotypic tendency of the body length. (C) Phenotypic tendency of the guard hair length. (D) Phenotypic tendency of the underfur length. (E) Phenotypic tendency of the two hair rate.
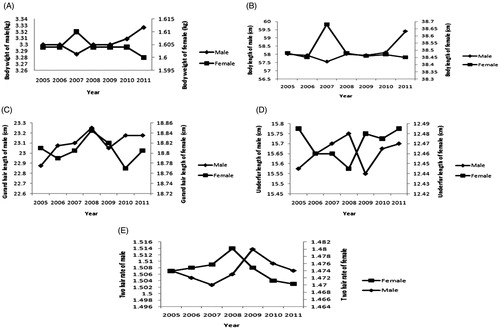
Figure 2. Genetic change tendency of growth and fur trait in silver blue mink.
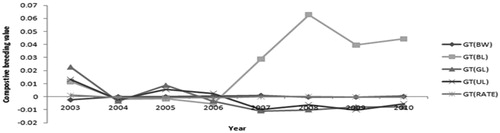
Table 3. Genetic trends measured as regression of yearly mean EBV for body weight, body length, guard hair length, underwool length, the rate of two hairs.
The genetic trends of BW and BL were downward during 2003 and 2006, the trend of BL was steep upward after 2006. The annual genetic trend for BW and BL found in this study was smaller than that reported by Socha and Kolodziejczyk (Citation2006) for standard and pastel colour-type mink body size and conformation (0.1) between 1996 and 2004. In mink selection, high body weight has shown a positive response within a few generations. This is the case both with selection for early body weight at four weeks after birth and selection for body weight in September and in November (Nielsen et al. Citation2012). Thus, the main reason for the differences in the genetic trends between these colour-type mink might be the result of different breeding model and feeding management. Our results show that there has been small genetic improvement in BW and BL and indicate that as the important role of breeding goal, BW and BL particularly for BL, will be effective in silver blue mink in long-term selection. This is in agreement with the heritability estimates for BW and BL obtained in the current and in a previous study for mink and fox (Peura et al. Citation2004b; Nielsen et al. Citation2012). The genetic changes of GL, UL and RATE between 2003 and 2010 appear to be flat, which was in agreement with the studies of Socha and Kolodziejczyk (Citation2006) on fur quality of mink. The low and negative values of these trends may prove the difficulties in the selection on traits of fur quality in mink population.
Conclusions
After six generations breeding, the main economic traits of silver blue mink were stable and showing good population characteristics. Results of this study show that growth and fur quality traits of sliver blue mink are highly heritable and should respond to selection. The observed genetic trends confirm that selective breeding can lead to significant genetic improvement in mink.
Disclosure statement
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of this article.
Related Research Data
References
- Cai W, Casey DS, Dekkers JCM. 2008. Selection response and genetic parameters for residual feed intake in Yorkshire swine. J Anim Sci. 86:287–298.
- Hansen B, Berg P. 2008. Reduced litter size and percent kits alive is a consequences of selecting for high body weight. Scientifur. 32:15.
- Kempe R, Koskinen N, Peura J, Koivula M, Stranden I. 2009. Body condition scoring method for blue fox (Alopex lagopus). Acta Agric Scand A Anim Sci. 59:85–92.
- Kempe R, Koskinen N, Mantysaari E, Stranden I. 2010. The genetics of body condition and leg weakness in the blue fox (Alopex lagopus). Acta Agric Scand A Anim Sci. 60:141–150.
- Nielsen VH, Møller SH, Hansen BK. 2012. Genetic parameters and effect of selection for body weight in lines of mink (Neovison vison) on ad libitumand restricted feeding. Acta Agric Scand A Anim Sci. 62:24–28. 3261–3272.
- Peura J, Stranden I, Smeds K. 2004a. Genetics of litter size, age at first insemination and animal size in blue fox (Alopex lagopus). Scientifur. 28:206–210.
- Peura J, Serenius T, Stranden I. 2004b. Economic weights for litter size and skin character traits in Finnish blue fox production. Paper presented at: Proceedings of the XXI Genetic Days. 1–3.
- Sengul T, Kiraz S. 2005. Non-linear models of growth curves in large white turkeys. J Vet Anim Sci. 29:331–337.
- Socha S, Kolodziejczyk D. 2006. Genetic parameters of animal size and fur quality in standard and pastel mink (Mustela vison). Acta Fytotech Zootech. 108–111.
- Wierzbicki H, Jagusiak W. 2006. Breeding value evaluation in Polish fur animals: estimates of (co)variances due to direct and litter effects for fur coat and reproduction traits. Czech J Anim Sci. 51:39–46.
- Zongyue L, Fangyong N, Zhiheng D, Chunsan Y, Jing F, Xing W, Xiujuan B. 2011. Modeling growth of five color types of mink. S Afr J Anim Sci. 41:116–125.