

ABSTRACT
The following review focuses on the scientific studies related to the role of endocannabinoid system (ECS) in pancreatic islet physiology and dysfunction. Different natural or synthetic agonists and antagonists have been suggested as an alternative treatment for diabetes, obesity and metabolic syndrome. Therapeutic use of Cannabis led to the discovery and characterization of the ECS, a signaling complex involved in regulation of various physiological processes, including food intake and metabolism. After the development of different agonists and antagonists, evidence have demonstrated the presence and activity of cannabinoid receptors in several organs and tissues, including pancreatic islets. Insulin and glucagon expression, stimulated secretion, and the development of diabetes and other metabolic disorders have been associated with the activity and modulation of ECS in pancreatic islets. However, according to the animal model and experimental design, either endogenous or pharmacological ligands of cannabinoid receptors have guided to contradictory and paradoxical results that suggest a complex physiological interaction. In consensus, ECS activity modulates insulin and glucagon secretions according to glucose in media; over-stimulation of cannabinoid receptors affects islets negatively, leading to glucose intolerance, meanwhile the treatment with antagonists in diabetic models and humans suggests an improvement in islets function.
Graphical Abstract
Overview of the endocannabinoid system components
The endocannabinoid system (ECS) is a lipid signaling complex involved in metabolism regulation and specific physiological processes. Today, ECS role in the development and treatment of some diseases, including metabolic syndrome and type-2 Diabetes Mellitus, is still under research.Citation1,Citation2 The ECS was discovered after the use of tetrahydrocannabinol (THC) and the activation of a transmembrane G protein-coupled receptor (GPR) in the nervous system, renamed cannabinoid receptor 1 (CB1).Citation3,Citation4 CB2 and GPR55 receptors were discovered by their interaction with endogenous ligands identified as endocannabinoids (EC): N-arachidonyl ethanolamine, also named anandamide (AEA), and 2-arachidonylglycerol (2-AG), both derived from phospholipids containing arachidonic acid (AAc).Citation5–7 Enzymes involved in biosynthesis and degradation of EC are present in many cells, anchored in the plasmatic membrane or close to: AEA production is regulated by N-acyl phosphatidylethanolamine phospholipase D (NAPE-D), and degradation by fatty acid-acyl hydrolase (FAAH), meanwhile 2-AG depends of diacylglycerol lipase (DAGL) for synthesis and monoacyl-glycerol lipase (MAGL) for metabolism.
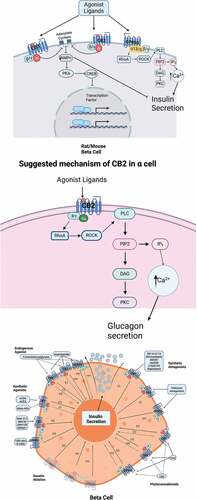
Figure 1. Mechanisms of action of cannabinoid receptors proposed for pancreatic beta cell: CB1 and CB2 receptors decreases cAMP, and GPR55 is associated with intracellular calcium increase.
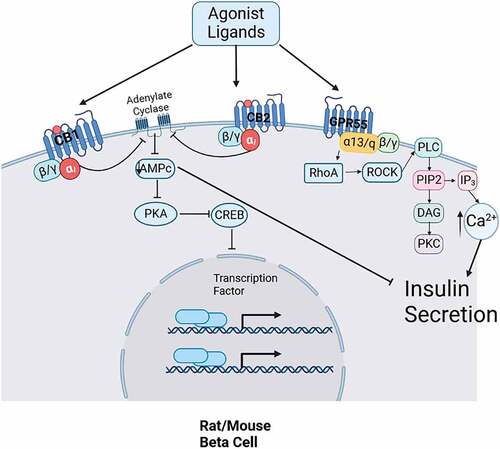
Figure 2. Mechanisms of action of cannabinoid receptors proposed for pancreatic alpha cell. CB2 receptor probably is coupled to PLC and PKC activity, increasing glucagon secretion.
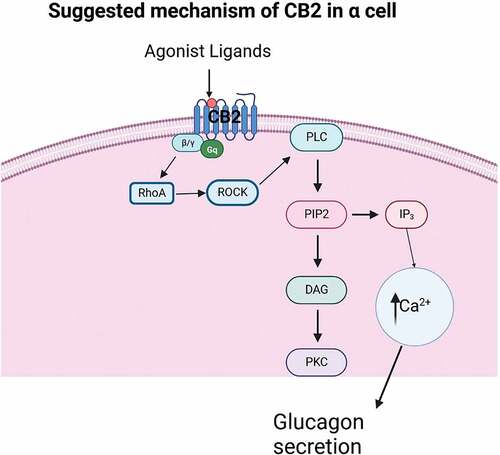
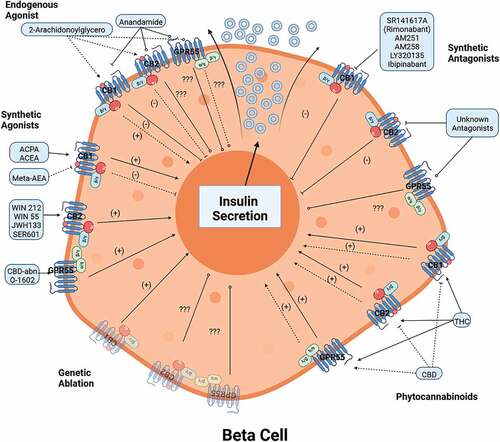
Figure 3. Modulation of insulin secretion with natural agonists, synthetic agonists and antagonists, and genetic deletion of CB receptors: (+) stimulation and enhance, (-) inhibition, (???) unknown effect or incomplete description. AEA (Anandamide), ACPA (arachidonyl cyclopropylamine), ACEA (arachidonyl-2-chloroethylamine), abn-CBD (abnormal Cannabidiol), CBD (Cannabidiol) and THC (Tetrahydrocannabinol).
After the psychoactive effects of delta-9-tetrahydrocannabinol in brain, CB1 receptor functions were discovered in basal ganglia, hippocampus, hypothalamus, cerebellum, olfactory cortex, septum, amygdala, brainstem, nerve terminals, spinal cord nodes and periaqueductal gray matter. The effects are associated to food intake modulation, motor coordination and pain sensitivity.Citation8–12 CB1 receptor activity inhibits neurotransmitters release in the presynaptic terminals, such as glutamate, dopamine or GABA, mobilizing vesicles or regulating ion channels activity.Citation11,Citation12 2-AG synthesis is provided by DAGL located at postsynaptic neurons, and its inactivation by MAGL activity is present in presynaptic neurons, establishing a retrograde mechanism of action.Citation13–15 In a similar way, synthesis and degradation of AEA by NAPE-PLD and FAAH are present in the plasmatic membrane of postsynaptic neurons, plus the high expression of CB1 receptor, suggests a local predominance of this system.Citation16,Citation17 CB2 receptor is highly expressed in immune cells, and in lower proportion at neurons from cerebellum and brainstem, liver, pancreas, muscle and adipose tissue.Citation18 It has been suggested the participation of a membrane transporter for ECs, not yet characterized, could facilitate release and uptake at both synapse membranes, and promote intracellular signaling mechanisms.Citation19,Citation20
CB1 and CB2 receptors are expressed in several peripheral tissues, executing physiological and pharmacological responses according to specific interactions with other proteins. Mostly CB receptors, as homo or heterodimers, are coupled to Gi/o proteins, inhibit adenylate cyclase (AC), regulate the activity of Ca2+ and K+ ion channels, promote mitogen-activating protein kinase (MAPK) signaling with p42/44 MAPK, p38 MAPK and c-JUN N-terminal kinase, and regulate intracellular location of specific nuclear transcription factors.Citation21,Citation22 Some cells present CB1 and CB2 receptors associated to Gs proteins, inducing changes in cAMP, intracellular Ca2+ streams, and increase phospholipases activity.Citation23
Orphan GPR55 receptor is expressed in some tissues involved in vascular physiology and nutrients metabolism. Some refer to GPR55 as CB3 receptor after specific binding to AEA, cannabidiol (CBD), abnormal cannabidiol (abn-CBD), virodhamine, palmitoylethanolamide and oleoylethanolamide.Citation24,Citation25 GPR55 activity is coupled to Gα12/13 proteins, associated with RhoA, cdc42 and rac1 activation. Its interaction with Gαq proteins induces changes in intracellular Ca2+ and extracellular signal-regulated kinase ERK ½ phosphorylation.Citation26,Citation27
ECS plays a key role in the energy metabolism and homeostasis of lipids and glucose through hypothalamic and mesolimbic pathways, and in adipocytes, liver and pancreatic islets as peripheral pathways.Citation28–32 At physiological concentrations, agonists and antagonists of CB receptors have shown specific short-effects on food ingestion, glucose and lipid metabolism; chronic or pharmacological exposure correlates to cellular dysfunction, ECS dysregulation, and the development of metabolic and eating disorders including abdominal obesity, dyslipidemia, metabolic syndrome and diabetes.Citation31,Citation32 In this review we spotlight these effects in pancreatic islets.
The ECS function in pancreatic islets
The presence and activity of the ECS in pancreatic islets have been reported in different models: cell lines, primary culture of intact islets or dispersed cells; in histological slides of pancreas obtained from rats, mice, and humans, under healthy, obesity of diabetic conditions.Citation33–45 Gene expression and localization of cannabinoid receptors, synthesis/degradation enzymes, and the local ECs production have been determined by different approaches, including immunochemestry, immunoflorescence, conventional PCR and Real-Time PCR, leading to controversial and inconclusive results: the inherent characteristics of mice, rats and human islets are well documented under different ages and in diabetes development.Citation33–45 The use of antibodies from diverse sources recognizes different epitopes that contributes to the inexact location of each ECS element. Apparently, the expression of receptors and enzymes is associated to specific culture conditions, tissue extraction, and the health state of patients, donors, or animals. The shared conclusion leads to a dynamic ECS expression and activity in islets, close-related to the changes in glucose input, diet and the development of diabetes.Citation43,Citation44
In beta-cell lines and islets isolated from mice and rat, CB1 and CB2 receptors activated with AEA from physiological (10–100 nM) to pharmacological (1 μM) concentrations, or with synthetic agonists, potentiate glucose-stimulated insulin secretion after increasing intracellular Ca2+ concentration, but reducing cyclic AMP production.Citation34,Citation35,Citation42 In opposition, there are evidences that AEA and 2-AG decrease insulin secretion with variable effects on intracellular calcium.Citation37,Citation38,Citation41 The conditions and origin of cell culture, ligands concentration, time of exposure, experimental approaches, even the source of reagents could be the reasons of these disparities, evoking opposite results; it is probable that some agonists change to inverse agonists, or under specific conditions CB receptors present different G-proteins coupling.Citation18–20,Citation46–49 Under physiological circumstances, ECS is associated to food ingestion and energy balance,Citation28–32 including insulin and glucagon secretions as modulators of glucose homeostasis and metabolism. The over-stimulation of CB receptors and the increasing presence of ECs in blood has been correlated with hyperinsulinemia, hyperglycemia and lipogenesis, suggesting negative effects on islets function.Citation33,Citation45,Citation50 Our hypothesis is ESC present a role as paracrine regulator: under fasting state, glucagon secretion is promoted among with a discrete basal insulin secretion. Since CB1 receptor and enzymes for synthesis and degradation of AEA are mainly expressed in islets, probably this endocannabinoid control basal insulin secretion in beta-cells. CB2 receptor and enzymes involved in 2-AG production are low expressed in islets, probably in correlation to the percent of alpha-cells, suggesting glucagon secretion could be regulated by CB1 and CB2 receptors.Citation51,Citation52 Considering evidences of GPR55 activity involved in insulin secretion,Citation47,Citation53–55 the ECS role in islets suggests an integral and paracrine regulation of metabolic homeostasis, therefore, a potential therapeutic option for diabetes. Solid evidences suggest LH-21 and abnormal cannabidiol as pharmacological strategy to improve beta-cell function in human and mouse islets, using different mechanism depend and independent of GPR55 interaction ().Citation55
Islets exposed to endogenous CB receptors agonists
In beta-cell lines and dispersed islets from mice, short-term activation of CB1 receptor by AEA reduced glucose-induced insulin secretion, but cytosolic Ca2+ oscillations were inhibited only in 50% of the cells, suggesting that CB1r is not present in all islet cells.Citation37,Citation38 2-AG exposure presented a similar effect, related to the permissive pulsatile insulin release.Citation40 In opposition, pancreatic islets from rat and beta-cell lines treated for 1 or 24 hours with pharmacological concentrations of AEA, increased basal and glucose-stimulated insulin secretion.Citation35,Citation42 2-AG promotes cAMP production, elevation in intracellular Ca2+, thus induces insulin secretion and decreases basal glucagon secretion.Citation33,Citation42–44 The explanation for these contrary observations is not clear, authors suggest different expression of ECS elements within islets, possibly a random distribution and activity, even between the same cell lineage.Citation43,Citation51,Citation52 AEA and 2-AG binding to CB receptors are well-described in neurons and other cell types, but there are not specific studies in pancreatic islets, suggesting a possible cross interaction and activation using pharmacological doses.Citation18,Citation20 In obese and diabetic models, AEA and 2-AG production inside the islets is increased, probably induce an over-activation of the ECS, leading to interference in metabolic mechanisms involved in insulin secretion (decrease in cAMP, changes in gene expression, insulin resistance).Citation33,Citation45,Citation56 The mechanism of CB1r action suggested for beta-cells is the formation of heterotrimeric complex with insulin receptor and Giα protein, which results in inhibition of insulin signaling by autophosphorylation.Citation50
Islets treated with synthetic CB receptors agonists
The development of pharmacological analogs and agonists, first for CB1 receptor and then for CB2 receptor, has been a useful in the study of mechanisms of action, binding and biological effects.Citation1,Citation2,Citation5,Citation45,Citation51,Citation52,Citation56–58 CB1 receptor agonist methanandamide increased intracellular Ca2+ oscillations in 31.3% of the beta-cells tested (10 out of 32), similarly to the secondary effect of AEA.Citation37 Selective agonists for CB1 receptor, arachidonyl cyclopropylamine (ACPA) and arachidonyl-2-chloroethylamine (ACEA), as well as JWH-015 for CB2 receptor, have been usually studied in isolated islets from animal models and humans.Citation43,Citation50–52,Citation56–59 At short exposure, ACEA increases the release of intracellular Ca2+ by activating the phosphatidyl-inositol-triphosphate (IP3) receptor in the endoplasmic reticulum, however the effect on insulin secretion is not clear since some studies demonstrate an inhibition, while others propose a significant increase of basal secretion.Citation51,Citation58 Isolated islets from mice and human chronically exposed to ACEA induced an adaptable ECS response, probably involving a process of saturation and self-regulation of receptors’ expression, meanwhile acute exposition decreased insulin secretion and apoptosis.Citation51,Citation52,Citation56 Acute and chronic effects of JWH-015 were most evident in alpha-cells, evoking increase in glucagon secretion, content, and gene expression.Citation51,Citation52
Islets exposed to synthetic CB receptors antagonists
The synthetic compound SR141716A, also called rimonabant, is a highly selective antagonist for CB1 receptor, suggested for the treatment of overweight, obesity and diabetes, but suspended until today.Citation42,Citation60,Citation61 Clinical studies demonstrated rimonabant reduced significantly adipose mass on obese and diabetic patients, increasing adiponectin expression, lipolysis, and decreasing pro-inflammatory and other cardiovascular risk factors.Citation60–64 The study of rimonabant in obesity (RIO) showed several benefits in obese and non-obese patients: 20 mg/day treatment improved carbohydrate and lipid metabolisms in peripheral tissues, mainly skeletal muscle, adipose tissue and liver.Citation63 The effects on insulin secretion and sensitivity induced a significant reduction in glycated hemoglobin percent and LDL level.Citation63
In Zucker fatty rats and mice with diet-induced metabolic syndrome, chronic rimonabant treatment improved oral glucose tolerance and insulin resistance. Specific analysis of pancreatic islets correlated insulin expression, content and secretion with slowing beta-cell dysfunction: 60% reduction on islet-cell surface (in these models, the islet mass and area increases in correlation with dysfunction), and 50% less disorganized islets, with a normal distribution of cells.Citation60,Citation64,Citation65
However, rimonabant was discontinued after undesired side effects reported, including dizziness, psychiatric disorders, symptoms of depression, anxiety, and the development of suicidal intention.Citation66,Citation67 In the last decades two different CB1 receptor antagonists have been synthetized, BAR-1 and Ibipinabant.Citation42,Citation68–70 In isolated pancreatic islets from rats, BAR-1 increased insulin, glucokinase, PDX-1 and CB1r gene expression, and basal insulin secretion was modified, in opposition to AEA effect.Citation42 BAR-1 treatment of prediabetic and diabetic mice reduced oral glucose tolerance without effect on HbA1c and triglycerides; glucose-stimulated insulin secretion was enhanced, but gene expression for insulin, glucagon, PDX-1 and some elements of ECS showed significant decrease.Citation68 Streptozocin-induced diabetic mice presented a discrete increase in islets mass and insulin-positive cells after BAR-1 exposure, suggesting a protective effect against apoptosis.Citation68 Ibipinabant is another rimonabant analog tested in animal models that improved glucose tolerance, insulin sensitivity, weight loss, triglycerides in blood, and induced a significant reduction in beta-cell lost.Citation69,Citation70
In the last decade, AM251, AM281 and LY320135 were proposed as synthetic antagonists or reverse agonists for cannabinoid receptors. Isolated human and mouse islets short-term exposed to AM251 increased beta-cell proliferation after cytokines stimulation.Citation46,Citation47,Citation71 As a CB1 receptor antagonist AM251 exerts stimulatory effect on glucose-stimulated insulin secretion in isolated islets from mice and human, but as GPR55 agonist enhances GLP-1 receptor activity, and therefore, potentiates insulin secretion.
Genetic ablation of CB1 receptor in islets
Obese mice and rats with modified gene expression or knock-out for CB1 receptor have reduced food intake and body fat deposits, showed a sustained loss weight, reversible changes in several metabolites, and increased plasmatic levels of leptin, insulin and adiponectin.Citation47,Citation48,Citation56,Citation72,Citation73 In mice with specific CB1 receptor -/- knock-out in pancreatic beta-cells, food intake was diminished while a significant increase in fasting insulin and insulin secretion during a glucose-lipid challenge was observed.Citation73 Isolated islets from these animals increased glucose-stimulated insulin secretion, preceded by elevated cAMP levels. Gene ablation also modified beta-cells viability, induced a discrete proliferation, and protection against diet-induced inflammation, supporting the interaction with insulin receptor and other signaling pathways.Citation22,Citation56,Citation73–75 These results suggest CB1 receptor has a negative regulatory function in beta-cell as mediator of inflammation under metabolic stress condition. Pancreatic beta-cells and hepatocytes express a specific isoform of CB1 receptor; exposure to the reverse agonist JD-5037 increase insulin secretion associated to cAMP production, suggesting it as a potential therapeutic target for metabolic disorders.Citation48
Islets exposed to natural phytocannabinoids
The use of some Cannabis derivatives as alternative treatment for diabetic patients is still on debate. Chronic marijuana diabetic users usually present low fasting insulin levels, reduced HOMA-IR index, adequate waist circumference, and low prevalence of complications.Citation3,Citation4,Citation76,Citation77 Diabetic rats treated chronically with 3 to 5 mg/day of THC presented improvement in glucose tolerance, total cholesterol, HDL and LDL, small decrease in body weight, increased total area of islets, and upregulation of gene expression for CB1 receptor, glucose transporter GLUT2, uncoupling protein UCP2, and protein kinase B.Citation49,Citation78 These studies suggest THC treatment could improve hyperlipidemia, hyperglycemia, and islet dysfunction.Citation25,Citation26 However, we assume that therapeutic effects of THC should be promoted as agonist of GPR55 receptor, after CB1 receptor presents oversaturation at nanomolar range. Unpublished data from our group suggest chronic exposure to THC increases insulin secretion in isolated islets, and improves oral glucose tolerance test in prediabetic mice, but decrease gene expression meanwhile islet structure remains unchanged (García-Luna and Vilches-Flores, 2023). To date, there are no direct evidences of the effect of THC on human islets.
Other receptors and signaling pathways related to ECS
CB1 receptor agonists AEA, N-arachidonoyl dopamine and N-arachidonylglycine can activate the transient potential receptor V1 (TRVP-1) or GPR18 receptor in pancreatic islets, elevate intracellular Ca2+ and potentiate insulin secretion, through coupling with the PI-PLC pathway and insulin receptor signaling.Citation38,Citation58,Citation74 CB1 receptor coupled to heterodimers of Gαi proteins interacts with insulin receptor through direct binding to the tyrosine-kinase domain, affecting the activation loop; consequently, phosphorylation of pro-apoptotic protein Bad is reduced, leading to apoptotic activity.Citation50,Citation56 CB1 receptor blockade or reduced expression improves insulin signaling by increase in cAMP production, leading to improvements on hyperglycemia, cell survival, and islet cells proliferation.Citation72 Other evidence demonstrate ECs regulate CB1 receptor interaction with focal adhesion kinase (FAK), mediating insulin exocytosis process by cytoskeleton rearrangement.Citation48,Citation79 In addition, CB1 receptor can interact with the mammalian target of rapamycin complex 1 (mTORC1) signaling pathway; in isolated islets, antagonists for CB1 and CB2 receptors decreased glucose-stimulated insulin secretion and mTOR activity, meanwhile in vivo they induced glucose intolerance and insulin resistance.Citation75
ECS expressed in adipose tissue, liver and pancreatic islet present a complex interaction and coordination between them to regulate glucose homeostasis. Treatment with CB2 receptor agonists JWH-015 and SER 601 induces changes in gene expression of the ESC elements, in liver, adipose tissue and pancreatic islets from mice; insulin secretion, glucose and lipid metabolisms, cell signaling, and immune responses in diabetic animal models are improved.Citation31,Citation33,Citation40,Citation41,Citation45,Citation51,Citation52,Citation68,Citation80 The function of GPR55 as part of the ECS within pancreatic islet is still under study, pointing to different pharmacological strategies will be target in future. Synthetic agonists abbnormal cannabidiol (abn-CBD) and O-1602 improve the function and viability of beta-cells from humans and rats, potentiate glucose-stimulated insulin secretion through increase cAMP and intracellular Ca2+, reduce cytokine-induced apoptosis, cytotoxic effects, and increase AKT phosphorylation ().Citation53–55
Implications of the ECS in the development of pathologies associated with the islet
Under physiological conditions the ECS in pancreatic islets participates integrally in the regulation of insulin secretion, according to food intake and in close coordination with other peripheral organs. We have exposed how CB receptors on islets, modulated by agonists and antagonists, are close-related to glucose metabolism, synthesis, secretion and action of insulin. ECs high production and the over-activation of cannabinoid receptors have important implications in white adipose tissue, hepatocytes and pancreatic islets, inducing deficiencies in hormonal secretion, hyperglycemia and metabolism imbalance.Citation29–33,Citation45,Citation62 Impaired or over-activated ECS presents a direct correlation with weight gain, decreased insulin sensitivity, hyperinsulinemia, glucose intolerance and hyperlipidemia, contributing to the development of obesity and type-2 diabetes mellitus.Citation35,Citation39–42,Citation52,Citation60,Citation64–66
The chronic hyperactivity of CB receptors and the elevated EC production have been associated with significant alterations in insulin secretion, dysfunction of pancreatic islets and type-2 diabetes progression. It is not clear the therapeutic role of THC, cannabidiol and other agonists of the cannabinoid receptors in islets, but evidence suggest a complex interaction and adaptative response over chronic exposure. CB1 receptor antagonists are proposed as therapeutic alternative to delay islet cell dysfunction, reduce obesity and improve insulin sensitivity, decrease blood triglyceride levels and weight, through insulin- and glucagon-mediated actions.Citation49,Citation61,Citation63,Citation77
Acknowledgments
This review and our research line is supported by PAPIIT-DGAPA UNAM, grant project IN225819.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Di Marzo V. Targeting the endocannabinoid system: to enhance or reduce? Nat Rev Drug Discov. 2008;7(5):438–455. doi:10.1038/nrd2553. PMID: 18446159.
- Di Marzo V. New approaches and challenges to targeting the endocannabinoid system. Nat Rev Drug Discov. 2018;17(9):623–639. doi:10.1038/nrd.2018.115. PMID: 30116049.
- Bellocchio L, Inchingolo AD, Inchingolo AM, Lorusso F, Malcangi G, Santacroce L, Scarano A, Bordea IR, Hazballa D, D’Oria MT, et al. Cannabinoids drugs and oral health-from recreational side-effects to medicinal purposes: a systematic review. Int J Mol Sci. 2021;22:8329. doi:10.3390/ijms22158329. PMID: 34361095.
- Grotenhermen F, Müller-Vahl K. The therapeutic potential of cannabis and cannabinoids. Dtsch Arztebl Int. 2012;109(29–30):495–501. doi:10.3238/arztebl.2012.0495. PMID: 23008748.
- Bisogno T, Sepe N, Melck D, Maurelli S, De Petrocellis L, Di Marzo V. Biosynthesis, release and degradation of the novel endogenous cannabimimetic metabolite 2-arachidonoylglycerol in mouse neuroblastoma cells. Biochem J. 1997;322(Pt 2):671–677. doi:10.1042/bj3220671. PMID: 9065792.
- Bisogno T, Maurelli S, Melck D, De Petrocellis L, Di Marzo V. Biosynthesis, uptake, and degradation of anandamide and palmitoylethanolamide in leukocytes. J Biol Chem. 1997;272(6):3315–3323. doi:10.1074/jbc.272.6.3315. PMID: 9013571.
- Bisogno T, Howell F, Williams G, Minassi A, Cascio MG, Ligresti A, Matias I, Schiano-Moriello A, Paul P, Williams EJ, et al. Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain. J Cell Biol. 2003;163:463–468. doi:10.1083/jcb.200305129. PMID: 14610053.
- Herkenham M, Lynn AB, Little MD, Johnson MR, Melvin LS, de Costa Br, Rice KC, de Costa BR. Cannabinoid receptor localization in brain. Proc Natl Acad Sci USA. 1990;87(5):1932–1936. doi:10.1073/pnas.87.5.1932. PMID: 2308954.
- Tsou K, Brown S, Sañudo-Peña MC, Mackie K, Walker JM. Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience. 1998;83(2):393–411. doi:10.1016/s0306-4522(97)00436-3. PMID: 9460749.
- Freund TF, Katona I, Piomelli D. Role of endogenous cannabinoids in synaptic signaling. Physiol Rev. 2003;83(3):1017–1066. doi:10.1152/physrev.00004.2003. PMID: 12843414.
- Schlicker E, Kathmann M. Modulation of transmitter release via presynaptic cannabinoid receptors. Trends Pharmacol Sci. 2001;22(11):565–572. doi:10.1016/s0165-6147(00)01805-8. PMID: 11698100.
- Chevaleyre V, Castillo PE. Heterosynaptic LTD of hippocampal GABAergic synapses: a novel role of endocannabinoids in regulating excitability. Neuron. 2003;38(3):461–472. doi:10.1016/s0896-6273(03)00235-6. PMID: 12741992.
- Dinh TP, Carpenter D, Leslie FM, Freund TF, Katona I, Sensi SL, Kathuria S, Piomelli D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc Natl Acad Sci USA. 2002;99(16):10819–10824. doi:10.1073/pnas.152334899. PMID: 12136125.
- Okamoto Y, Morishita J, Tsuboi K, Tonai T, Ueda N. Molecular characterization of a phospholipase D generating anandamide and its congeners. J Biol Chem. 2004;279(7):5298–5305. doi:10.1074/jbc.M306642200. PMID: 14634025.
- Cravatt BF, Giang DK, Mayfield SP, Boger DL, Lerner RA, Gilula NB. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature. 1996;384(6604):83–87. doi:10.1038/384083a0. PMID: 8900284.
- Egertová M, Cravatt BF, Elphick MR. Comparative analysis of fatty acid amide hydrolase and cb(1) cannabinoid receptor expression in the mouse brain: evidence of a widespread role for fatty acid amide hydrolase in regulation of endocannabinoid signaling. Neuroscience. 2003;119(2):481–496. doi:10.1016/s0306-4522(03)00145-3. PMID: 12770562.
- Ligresti A, Morera E, Van Der Stelt M, Monory K, Lutz B, Ortar G, Di Marzo V. Further evidence for the existence of a specific process for the membrane transport of anandamide. Biochem J. 2004;380(Pt 1):265–272. doi:10.1042/BJ20031812. PMID: 14969584.
- Van Sickle MD, Duncan M, Kingsley PJ, Mouihate A, Urbani P, Mackie K, Stella N, Makriyannis A, Piomelli D, Davison JS, et al. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science. 2005;310:329–332. doi:10.1126/science.1115740. PMID: 16224028.
- Baggelaar MP, Maccarrone M, van der Stelt M. 2-Arachidonoylglycerol: a signaling lipid with manifold actions in the brain. Prog Lipid Res. 2018;71:1–17. doi:10.1016/j.plipres.2018.05.002. PMID: 29751000.
- Brailoiu GC, Oprea TI, Zhao P, Abood ME, Brailoiu E. Intracellular cannabinoid type 1 (CB1) receptors are activated by anandamide. J Biol Chem. 2011;286(33):29166–29174. doi:10.1074/jbc.M110.217463. PMID: 21719698.
- Turu G, Hunyady L. Signal transduction of the CB1 cannabinoid receptor. J Mol Endocrinol. 2010;44(2):75–85. doi:10.1677/JME-08-0190. PMID: 19620237.
- Nithipatikom K, Gomez-Granados AD, Tang AT, Pfeiffer AW, Williams CL, Campbell WB. Cannabinoid receptor type 1 (CB1) activation inhibits small GTPase RhoA activity and regulates motility of prostate carcinoma cells. Endocrinology. 2012;153(1):29–41. doi:10.1210/en.2011-1144. PMID: 22087025.
- Abadji V, Lucas-Lenard JM, Chin C, Kendall DA. Involvement of the carboxyl terminus of the third intracellular loop of the cannabinoid CB1 receptor in constitutive activation of Gs. J Neurochem. 1999;72(5):2032–2038. doi:10.1046/j.1471-4159.1999.0722032.x. PMID: 10217281.
- Baker D, Pryce G, Davies WL, Hiley CR. In silico patent searching reveals a new cannabinoid receptor. Trends Pharmacol Sci. 2006;27(1):1–4. doi:10.1016/j.tips.2005.11.003. PMID: 16318877.
- Pertwee RG. GPR55: a new member of the cannabinoid receptor clan? Br J Pharmacol. 2007;152(7):984–986. doi:10.1038/sj.bjp.0707464. PMID: 17876300.
- Ryberg E, Larsson N, Sjögren S, Hjorth S, Hermansson NO, Leonova J, Elebring T, Nilsson K, Drmota T, Greasley PJ. The orphan receptor GPR55 is a novel cannabinoid receptor. Br J Pharmacol. 2007;152(7):1092–1101. doi:10.1038/sj.bjp.0707460. PMID: 17876302.
- Lauckner JE, Jensen JB, Chen HY, Lu HC, Hille B, Mackie K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc Natl Acad Sci USA. 2008;105(7):2699–2704. doi:10.1073/pnas.0711278105. PMID: 18263732.
- Di Marzo V, Ligresti A, Cristino L. The endocannabinoid system as a link between homoeostatic and hedonic pathways involved in energy balance regulation. Int J Obes (Lond). 2009;33(2):S18–24. doi:10.1038/ijo.2009.67. PMID: 19528974.
- Pagotto U, Marsicano G, Cota D, Lutz B, Pasquali R. The emerging role of the endocannabinoid system in endocrine regulation and energy balance. Endocr Rev. 2006;27(1):73–100. doi:10.1210/er.2005-0009. PMID: 16306385.
- Viveros MP, de Fonseca Fr, Bermudez-Silva FJ, McPartland JM, de Fonseca F. Critical role of the endocannabinoid system in the regulation of food intake and energy metabolism, with phylogenetic, developmental, and pathophysiological implications. Endocr Metab Immune Disord Drug Targets. 2008;8(3):220–230. doi:10.2174/187153008785700082. PMID: 18782018.
- Matias I, Di Marzo V. Endocannabinoids and the control of energy balance. Trends Endocrinol Metab. 2007;18(1):27–37. doi:10.1016/j.tem.2006.11.006. PMID: 17141520.
- Bermudez-Silva FJ, Cardinal P, Cota D. The role of the endocannabinoid system in the neuroendocrine regulation of energy balance. J Psychopharmacol. 2012;26(1):114–124. doi:10.1177/0269881111408458. PMID: 21824982.
- Matias I, Gonthier MP, Orlando P, Martiadis V, De Petrocellis L, Cervino C, Petrosino S, Hoareau L, Festy F, Pasquali R, et al. Regulation, function, and dysregulation of endocannabinoids in models of adipose and beta-pancreatic cells and in obesity and hyperglycemia. J Clin Endocrinol Metab. 2006;91:3171–3180. doi:10.1210/jc.2005-2679. PMID: 16684820.
- Li C, Bowe JE, Jones PM, Persaud SJ. Expression and function of cannabinoid receptors in mouse islets. Islets. 2010;2(5):293–302. doi:10.4161/isl.2.5.12729. PMID: 21099327.
- Li C, Jones PM, Persaud SJ. Cannabinoid receptors are coupled to stimulation of insulin secretion from mouse MIN6 beta-cells. Cell Physiol Biochem. 2010;26(2):187–196. doi:10.1159/000320527. PMID: 20798502.
- Flores LE, Alzugaray ME, Cubilla MA, Raschia MA, Del Zotto HH, Román CL, Suburo AM, Gagliardino JJ. Islet cannabinoid receptors: cellular distribution and biological function. Pancreas. 2013;42(7):1085–1092. doi:10.1097/MPA.0b013e31828fd32d. PMID: 24005231.
- Juan-Picó P, Fuentes E, Bermúdez-Silva FJ, Javier Díaz-Molina F, Ripoll C, Rodríguez de Fonseca F, Nadal A. Cannabinoid receptors regulate Ca(2+) signals and insulin secretion in pancreatic beta-cell. Cell Calcium. 2006;39(2):155–162. doi:10.1016/j.ceca.2005.10.005. PMID: 16321437.
- Juan-Pico P, Ropero AB, Tuduri E, Quesada I, Fuentes E, Bermúdez-Silva FJ. Regulation of glucose-induced [Ca2+]i signals by cannabinoid CB1 and CB2 receptors in pancreatic α- and β-cells within intact islets of Langerhans. Obe Metab. 2009;5:1–9.
- Bermúdez-Siva FJ, Serrano A, Diaz-Molina FJ, Sánchez Vera I, Juan-Pico P, Nadal A, Fuentes E, Rodríguez de Fonseca F. Activation of cannabinoid CB1 receptors induces glucose intolerance in rats. Eur J Pharmacol. 2006;531(1–3):282–284. doi:10.1016/j.ejphar.2005.12.016. PMID: 16423347.
- Bermudez-Silva FJ, Sanchez-Vera I, Suárez J, Serrano A, Fuentes E, Juan-Pico P, Nadal A, Rodríguez de Fonseca F. Role of cannabinoid CB2 receptors in glucose homeostasis in rats. Eur J Pharmacol. 2007 22;565(1–3):207–211. doi:10.1016/j.ejphar.2007.02.066. PMID: 17499236.
- Bermúdez-Silva FJ, Suárez J, Baixeras E, Cobo N, Bautista D, Cuesta-Muñoz AL, Fuentes E, Juan-Pico P, Castro MJ, Milman G, et al. Presence of functional cannabinoid receptors in human endocrine pancreas. Diabetologia. 2008;51(3):476–487. doi:10.1007/s00125-007-0890-y. PMID: 18092149.
- Vilches-Flores A, Delgado-Buenrostro NL, Navarrete-Vázquez G, Villalobos-Molina R. CB1 cannabinoid receptor expression is regulated by glucose and feeding in rat pancreatic islets. Regul Pept. 2010;163(1–3):81–87. doi:10.1016/j.regpep.2010.04.013. PMID: 20451564.
- Anderson RL, Randall MD, Chan SL. The complex effects of cannabinoids on insulin secretion from rat isolated islets of Langerhans. Eur J Pharmacol. 2013;706(1–3):56–62. doi:10.1016/j.ejphar.2013.02.034. PMID: 23499687.
- Nakata M, Yada T. Cannabinoids inhibit insulin secretion and cytosolic Ca2+ oscillation in islet beta-cells via CB1 receptors. Regul Pept. 2008;145(1–3):49–53. doi:10.1016/j.regpep.2007.08.009. PMID: 17884194.
- Starowicz KM, Cristino L, Matias I, Capasso R, Racioppi A, Izzo AA, Di Marzo V. Endocannabinoid dysregulation in the pancreas and adipose tissue of mice fed with a high-fat diet. Obesity (Silver Spring). 2008;16(3):553–565. doi:10.1038/oby.2007.106. PMID: 18239598.
- Li C, Bowe JE, Huang GC, Amiel SA, Jones PM, Persaud SJ. Cannabinoid receptor agonists and antagonists stimulate insulin secretion from isolated human islets of Langerhans. Diabetes Obes Metab. 2011;13(10):903–910. doi:10.1111/j.1463-1326.2011.01422.x. PMID: 21564460.
- Ruz-Maldonado I, Liu B, Atanes P, Pingitore A, Huang GC, Choudhary P, Persaud SJ. The cannabinoid ligands SR141716A and AM251 enhance human and mouse islet function via GPR55-independent signalling. Cell Mol Life Sci. 2020;77(22):4709–4723. doi:10.1007/s00018-019-03433-6. PMID: 31925452.
- González-Mariscal I, Krzysik-Walker SM, Doyle ME, Liu QR, Cimbro R, Santa-Cruz Calvo S, Ghosh S, Cieśla Ł, Moaddel R, Carlson OD, et al. Human CB1 receptor isoforms, present in hepatocytes and β-cells, are involved in regulating metabolism. Sci Rep. 2016;6:33302. doi:10.1038/srep33302. PMID: 27641999.
- Levendal RA, Schumann D, Donath M, Frost CL. Cannabis exposure associated with weight reduction and β-cell protection in an obese rat model. Phytomedicine. 2012;19(7):575–582. doi:10.1016/j.phymed.2012.02.001. PMID: 22421529.
- Kim W, Doyle ME, Liu Z, Lao Q, Shin YK, Carlson OD, Kim HS, Thomas S, Napora JK, Lee EK, et al. Cannabinoids inhibit insulin receptor signaling in pancreatic β-cells. Diabetes. 2011;60:1198–1209. doi:10.2337/db10-1550. PMID: 21346174; PMCID: PMC3064093.
- Vilches-Flores A, Hauge-Evans AC, Jones PM, Persaud SJ. Chronic activation of cannabinoid receptors in vitro does not compromise mouse islet function. Clin Sci (Lond). 2013;124(7):467–478. doi:10.1042/CS20120447. PMID: 23078523.
- Vilches-Flores A, Franklin Z, Hauge-Evans AC, Liu B, Huang GC, Choudhary P, Jones PM, Persaud SJ. Prolonged activation of human islet cannabinoid receptors in vitro induces adaptation but not dysfunction. BBA Clin. 2016;5:143–150. doi:10.1016/j.bbacli.2016.03.009. PMID: 27114924.
- McKillop AM, Moran BM, Abdel-Wahab YH, Flatt PR. Evaluation of the insulin releasing and antihyperglycaemic activities of GPR55 lipid agonists using clonal beta-cells, isolated pancreatic islets and mice. Br J Pharmacol. 2013;170(5):978–990. doi:10.1111/bph.12356. PMID: 23992544.
- Liu B, Song S, Ruz-Maldonado I, Pingitore A, Huang GC, Baker D, Jones PM, Persaud SJ. GPR55-dependent stimulation of insulin secretion from isolated mouse and human islets of Langerhans. Diabetes Obes Metab. 2016;18(12):1263–1273. doi:10.1111/dom.12780. PMID: 27561953.
- Ruz-Maldonado I, Pingitore A, Liu B, Atanes P, Huang GC, Baker D, Alonso FJ, Bermúdez-Silva FJ, Persaud SJ. LH-21 and abnormal cannabidiol improve β-cell function in isolated human and mouse islets through GPR55-dependent and -independent signalling. Diabetes Obes Metab. 2018;20(4):930–942. doi:10.1111/dom.13180. PMID: 29205751.
- Kim W, Lao Q, Shin YK, Carlson OD, Lee EK, Gorospe M, Kulkarni RN, Egan JM. Cannabinoids induce pancreatic β-cell death by directly inhibiting insulin receptor activation. Sci Signal. 2012;5(216):ra23. doi:10.1126/scisignal.2002519. PMID: 22434934.
- Pertwee RG. Inverse agonism and neutral antagonism at cannabinoid CB1 receptors. Life Sci. 2005;76(12):1307–1324. doi:10.1016/j.lfs.2004.10.025. PMID: 15670612.
- De Petrocellis L, Marini P, Matias I, Moriello AS, Starowicz K, Cristino L, Nigam S, Di Marzo V. Mechanisms for the coupling of cannabinoid receptors to intracellular calcium mobilization in rat insulinoma beta-cells. Exp Cell Res. 2007;313(14):2993–3004. doi:10.1016/j.yexcr.2007.05.012. PMID: 17585904.
- Hillard CJ, Manna S, Greenberg MJ, DiCamelli R, Ross RA, Stevenson LA, Murphy V, Pertwee RG, Campbell WB. Synthesis and characterization of potent and selective agonists of the neuronal cannabinoid receptor (CB1). J Pharmacol Exp Ther. 1999;289(3):1427–1433. PMID: 10336536.
- Ravinet Trillou C, Arnone M, Delgorge C, Gonalons N, Keane P, Maffrand JP, Soubrie P. Anti-obesity effect of SR141716, a CB1 receptor antagonist, in diet-induced obese mice. Am J Physiol Regul Integr Comp Physiol. 2003;284(2):R345–353. doi:10.1152/ajpregu.00545.2002. PMID: 12399252.
- Jbilo O, Ravinet-Trillou C, Arnone M, Buisson I, Bribes E, Péleraux A, Pénarier G, Soubrié P, Le Fur G, Galiègue S, et al. The CB1 receptor antagonist rimonabant reverses the diet-induced obesity phenotype through the regulation of lipolysis and energy balance. FASEB J. 2005;19:1567–1569. doi:10.1096/fj.04-3177fje. PMID: 16009704.
- Kunos G, Osei-Hyiaman D, Liu J, Godlewski G, Bátkai S. Endocannabinoids and the control of energy homeostasis. J Biol Chem. 2008;283(48):33021–33025. doi:10.1074/jbc.R800012200. PMID: 18694938.
- Scheen AJ, Finer N, Hollander P, Jensen MD, Van Gaal LF; RIO-Diabetes Study Group. Efficacy and tolerability of rimonabant in overweight or obese patients with type 2 diabetes: a randomised controlled study. Lancet. 2006;368:1660–1672. doi:10.1016/S0140-6736(06)69571-8. PMID: 17098084.
- Duvivier VF, Delafoy-Plasse L, Delion V, Lechevalier P, Le Bail JC, Guillot E, Pruniaux MP, Galzin AM. Beneficial effect of a chronic treatment with rimonabant on pancreatic function and beta-cell morphology in Zucker fatty rats. Eur J Pharmacol. 2009;616(1–3):314–320. doi:10.1016/j.ejphar.2009.05.024. PMID: 19482020.
- Getty-Kaushik L, Richard AM, Deeney JT, Krawczyk S, Shirihai O, Corkey BE. The CB1 antagonist rimonabant decreases insulin hypersecretion in rat pancreatic islets. Obesity (Silver Spring). 2009;17(10):1856–1860. doi:10.1038/oby.2009.234. PMID: 19644453.
- Janero DR, Makriyannis A. Cannabinoid receptor antagonists: pharmacological opportunities, clinical experience, and translational prognosis. Expert Opin Emerg Drugs. 2009;14(1):43–65. doi:10.1517/14728210902736568. PMID: 19249987.
- Nathan PJ, O’Neill BV, Napolitano A, Bullmore ET. Neuropsychiatric adverse effects of centrally acting antiobesity drugs. CNS Neurosci Ther. 2011;17(5):490–505. doi:10.1111/j.1755-5949.2010.00172.x. PMID: 21951371.
- Nava-Molina L, Uchida-Fuentes T, Ramos-Tovar H, Fregoso-Padilla M, Rodríguez-Monroy MA, Vega AV, Navarrete-Vázquez G, Andrade-Jorge E, Villalobos-Molina R, Ortiz-Ortega R, et al. Novel CB1 receptor antagonist BAR-1 modifies pancreatic islet function and clinical parameters in prediabetic and diabetic mice. Nutr Diabetes. 2020;10:7. doi:10.1038/s41387-020-0110-0. PMID: 32132523.
- Chorvat RJ, Berbaum J, Seriacki K, McElroy JF. JD-5006 and JD-5037: peripherally restricted (PR) cannabinoid-1 receptor blockers related to SLV-319 (Ibipinabant) as metabolic disorder therapeutics devoid of CNS liabilities. Bioorg Med Chem Lett. 2012;22(19):6173–6180. doi:10.1016/j.bmcl.2012.08.004. PMID: 22959249.
- Rohrbach K, Thomas MA, Glick S, Fung EN, Wang V, Watson L, Gregory P, Antel J, Pelleymounter MA. Ibipinabant attenuates β-cell loss in male Zucker diabetic fatty rats independently of its effects on body weight. Diabetes Obes Metab. 2012;14(6):555–564. doi:10.1111/j.1463-1326.2012.01563.x. PMID: 22268426.
- Shin H, Han JH, Yoon J, Sim HJ, Park TJ, Yang S, Lee EK, Kulkarni RN, Egan JM, Kim W. Blockade of cannabinoid 1 receptor improves glucose responsiveness in pancreatic beta cells. J Cell Mol Med. 2018;22(4):2337–2345. doi:10.1111/jcmm.13523. PMID: 29431265.
- González-Mariscal I, Krzysik-Walker SM, Kim W, Rouse M, Egan JM. Blockade of cannabinoid 1 receptor improves GLP-1R mediated insulin secretion in mice. Mol Cell Endocrinol. 2016;423:1–10. doi:10.1016/j.mce.2015.12.015. PMID: 26724516.
- González-Mariscal I, Montoro RA, Doyle ME, Liu QR, Rouse M, O’Connell JF, Santa-Cruz Calvo S, Krzysik-Walker SM, Ghosh S, Carlson OD, et al. Absence of cannabinoid 1 receptor in beta cells protects against high-fat/high-sugar diet-induced beta cell dysfunction and inflammation in murine islets. Diabetologia. 2018;61:1470–1483. doi:10.1007/s00125-018-4576-4. PMID: 29497784.
- Ikeda Y, Iguchi H, Nakata M, Ioka RX, Tanaka T, Iwasaki S, Magoori K, Takayasu S, Yamamoto TT, Kodama T, et al. Identification of N-arachidonylglycine, U18666A, and 4-androstene-3,17-dione as novel insulin secretagogues. Biochem Biophys Res Commun. 2005;333:778–786. doi:10.1016/j.bbrc.2005.06.005. PMID: 15967412.
- Bermudez-Silva FJ, Romero-Zerbo SY, Haissaguerre M, Ruz-Maldonado I, Lhamyani S, El Bekay R, Tabarin A, Marsicano G, Cota D. The cannabinoid CB1 receptor and mTORC1 signalling pathways interact to modulate glucose homeostasis in mice. Dis Model Mech. 2016;9(1):51–61. doi:10.1242/dmm.020750. PMID: 26563389.
- Ewell TR, Abbotts KSS, Williams NNB, Butterklee HM, Bomar MC, Harms KJ, Rebik JD, Mast SM, Akagi N, Dooley GP, et al. Pharmacokinetic investigation of commercially available edible marijuana products in humans: potential influence of body composition and influence on glucose control. Pharmaceuticals (Basel). 2021;14:817. doi:10.3390/ph14080817. PMID: 34451914.
- Sidney S. Marijuana use and type 2 diabetes mellitus: a review. Curr Diab Rep. 2016 Nov;16(11):117. doi:10.1007/s11892-016-0795-6. PMID: 27747490.
- Coskun ZM, Bolkent S. Biochemical and immunohistochemical changes in delta-9-tetrahydrocannabinol-treated type 2 diabetic rats. Acta Histochem. 2014;116(1):112–116. doi:10.1016/j.acthis.2013.05.013. PMID: 23845579.
- Malenczyk K, Jazurek M, Keimpema E, Silvestri C, Janikiewicz J, Mackie K, Di Marzo V, Redowicz MJ, Harkany T, Dobrzyn A. CB1 cannabinoid receptors couple to focal adhesion kinase to control insulin release. J Biol Chem. 2013;288(45):32685–32699. doi:10.1074/jbc.M113.478354. PMID: 24089517.
- Zhang X, Gao S, Niu J, Li P, Deng J, Xu S, Wang Z, Wang W, Kong D, Li C. Cannabinoid 2 receptor agonist improves systemic sensitivity to insulin in high-fat diet/streptozotocin-induced diabetic mice. Cell Physiol Biochem. 2016;40(5):1175–1185. doi:10.1159/000453171. PMID: 27960161.

