Abstract
A new species of the Cletopsyllidae belonging to the genus Isocletopsyllus Huys and Lee, 1999 is described from the materials collected from subtidal sandy bottoms at Jeju Island and Ulleung Island, Korea. So far, only two species of the genus Isocletopsyllus, viz. I. tertius (Por, 1964) from Israel and I. quartus (Soyer, 1966) from Banyuls-sur-Mer (France) are recorded, both from the Mediterranean Ocean. The new species can be clearly distinguished from its congeners by the combination of the following characters: (1) female antennule with a large process on the outer margin of first segment, (2) caudal rami with a bulbous proximal inner expansion, and shorter than those of other species (about four times long as wide in the new species), (3) second segment of P1 exopod with a short rod-like inner seta, (4) sexual dimorphism presented in the second endopodal segment of male P2, not in exopod of P4, and (6) female P5 with a very long exopod with very short second outer seta. A table comparing the modifications of swimming legs in the family Cletopsyllidae Huys and Lee, 1999 is presented. This is the first record of this family from Korea.
Introduction
Jeju Island is the largest volcanic island off the south coast of Korea, and erupted about 2 million years ago. Its marine realm shows a high diversity of invertebrates (Je et al. Citation2002), due to its temperate and subtropical climate influenced by the warm Kuroshio current.
Although the region is known for its high faunal diversity, marine meiobenthic harpacticoid copepods from Jeju are poorly known. Song and Chang (Citation1993) first recorded five harpacticoid species of the family Harpacticidae in Jeju. Since then, only 18 species distributed in 15 genera and nine families have been recorded since 1993. All 18 species were found in different habitats of the island (Song and Chang Citation1993; Chang and Song Citation1995, Citation1997a, Citationb; Song et al. Citation1999, Citation2001, Citation2003; Nam and Lee Citation2006; Chang Citation2008) and most were found to be free-living species inhabiting algae or sediments, except for Neotachidius parvus Huys, Ohtsuka, Conroy-Dalton and Kikuchi, 2005 sensu Chang (2008) collected from brackish water, Porcellidium similis Kim and Kim, 1996 and Alteuthoides affinis Kim and Kim, 1998, associated with hermit crabs and sponges, respectively.
Of these 18 species, only eight species (Dactylopusia pauciarticulata Chang and Song Citation1997a; Paradactylopodia koreana Chang and Song Citation1997a; Parathalestris parviseta Chang and Song Citation1997b; Parastenhelia pyriformis Song, Kim and Chang 2003; Heteropsyllus coreanus Nam and Lee Citation2006; Neotachidius parvus Huys, Ohtsuka, Conroy-Dalton and Kikuchi, 2005; P. similis Kim and Kim, 1996 and A. affinis Kim and Kim, 1998) were new to science.
On the other hand, the additional specimens were collected from Ulleung Island. The small island is also a representative volcanic island off the east coast of Korea (the East Sea) and consists primarily of trachyandesite rock. Also the harpacticoid species of Ulleung Island are poorly known; only 12 species are reported from the macroalgae mainly. Since Song and Chang (Citation1993) first recorded Tigriopus japonicus Mori, 1938 and Zaus robustus Ito, 1974, subsequently 10 species were added to the fauna of Ulleung Island [Eudactylopus andrewi Sewell, 1940 and E. spectabilis (Brian, 1923) (Chang and Song 1995b), Parathalestris bulbiseta Lang, 1965, P. verrucosa Ito, 1970, P. pacificus Chislenko, 1971 and P. areolata Ito, 1972 (Chang and Song Citation1997a), Dactylopus pauciarticulata Chang and Song, Citation1997b and Paradactylopodia koreana Chang and Song, Citation1997b (Chang and Song Citation1997b), Diosaccus ezoensis Ito, 1974 (Song et al. Citation1999), and Phyllothalestris sarsi Sewell, 1940 (Song et al. Citation2001)].
In the course of benthic subtidal copepod study conducted in Jeju and Ulleung Islands during the period 2003–2006, a new species, Isocletopsyllus maximus of Cletopsyllidae, was found, and is herein described.
Materials and methods
Sediment samples were taken from Jeju Island and Ulleung Island (Korea). Specimens were fixed in 95% ethanol. Specimens were cleared and dissected in lactic acid. Dissected parts were mounted on slides in lactophenol as mounting medium. Preparations were sealed with transparent nail varnish. All drawings were prepared using an Olympus BX60 differential interference contrast microscope equipped with Nomarski optics and drawing tube.
The descriptive terminology is adopted from Huys et al. (Citation1996). Abbreviations used in the text are: ae, aesthetasc; exp, exopod; enp, endopod; P1–P6, first to sixth thoracopod; exp(enp)-1(2, 3) to denote the proximal (middle, distal) segment of a ramus. The term acrothek is used to denote the trifid seta complement found apically on the distal antennulary segment. Scale bars are in micrometers (µm).
Type series are deposited in the collections of The Natural Institute of Biological Resources (NIBR), Incheon, Korea.
Description
Order Harpacticoida Sars, 1903
Family Cletopsyllidae Huys and Willems, 1989
Genus Isocletopsyllus Huys and Lee, 1999
Isocletopsyllus maximus sp. nov. (Figures )
Figure 1. Isocletopsyllus maximus sp. nov. Female (paratype): A, habitus, dorsal; B, habitus, lateral. Scale bar=100 µm.
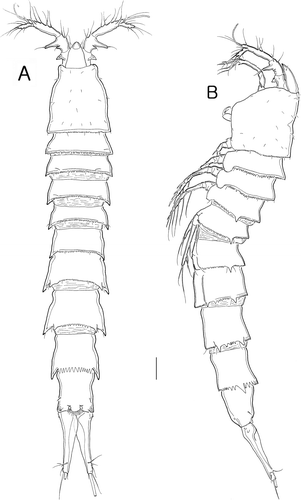
Figure 2. Isocletopsyllus maximus sp. nov. Female (paratype): A, mandible; B, maxillule; C, maxilla; D, maxilliped. Scale bar=30 µm.
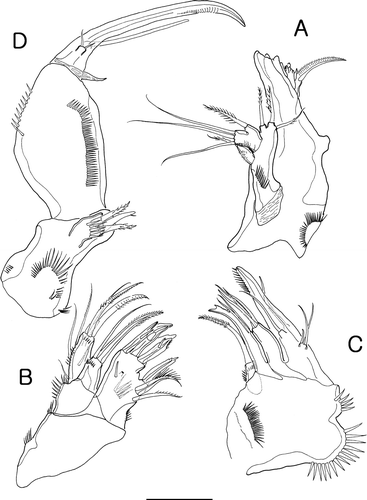
Figure 3. Isocletopsyllus maximus sp. nov. Female (paratype): A, antennule; B, antenna; C, caudal ramus, dorsal. Male (paratype): D, rostrum; E, antennule; F, 3rd segment of A1; G, 4th segment of A1; H, 5th segment of A1. Scale bars=30 µm.
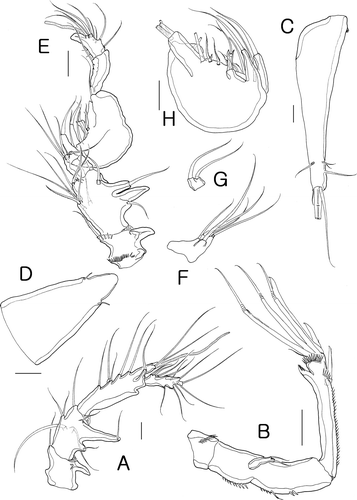
Figure 4. Isocletopsyllus maximus sp. nov. Female (paratype): A, P1; B, P2. Male (paratype): C, endopod of P2. Scale bars=30 µm.
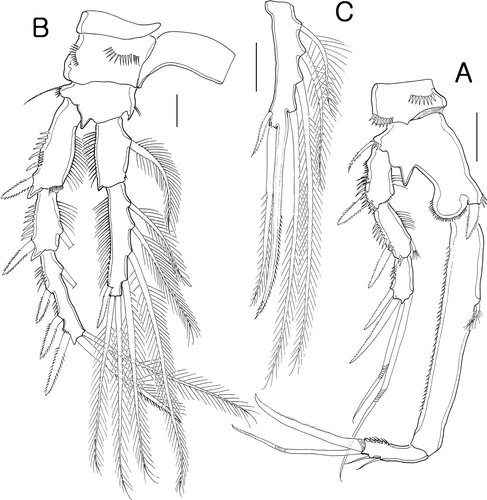
Figure 5. Isocletopsyllus maximus sp. nov. Female (paratype): A, P3; B, P4. Scale bar=30 µm.
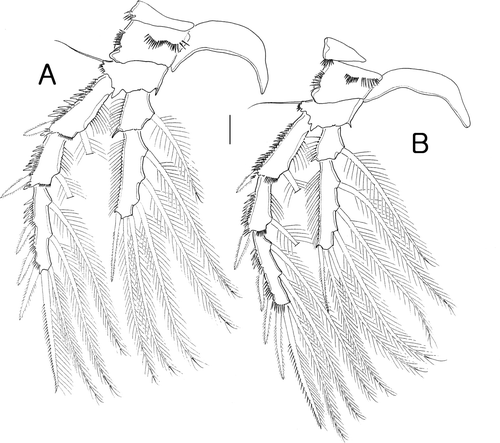
Figure 6. Isocletopsyllus maximus sp. nov. Female (paratype): A, P5. Male (paratype): B, habitus, dorsal; C, P5. Scale bars: A, C=30 µm; B=100 µm.
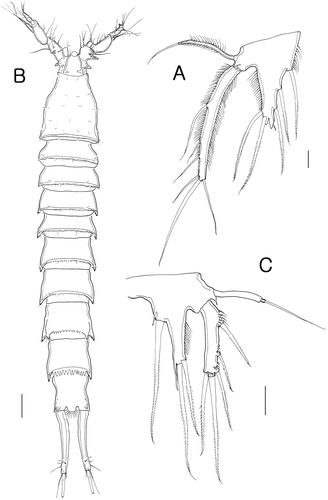
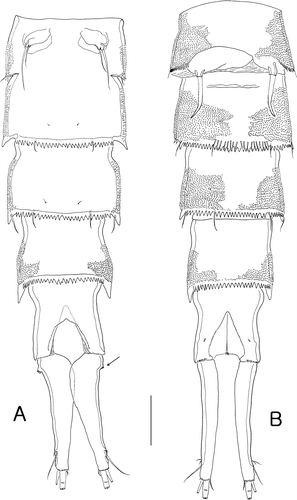
Figure 7. Isocletopsyllus maximus sp. nov. Female (paratype): A, urosome (excluding somite bearing P5). Male (paratype): B, urosome (excluding somite bearing P5). Scale=100 µm.
Type locality. Moonseom Islet, Jeju Is. South Sea of Korea (31°13′41?N, 126°33′57?E); 37 m depth, sandy bottom.
Type material. Holotype (reg. no. NIBRIV0000178485): ovigerous female preserved in alcohol; 8 July 2003, Coll. Jae Wan Choi and Dr Hyun Soo Rho.Paratypes: four males preserved in alcohol (reg. no. NIBRIV0000178486). One female and one male, dissected on 12 and 13 slides respectively, are kept in the personal research collection of the first author (SJS) (reg. no. SACOP3011-3017) – having the same collection data as the holotype.
Additional materials. Two female and one male collected from Ssangjeongcho, Ulleung Is. (37°31′20?N, 130°55′58?E), 20 m depth, sandy bottom, coll. Sung Joon Song, by SCUBA diving, 22 June 2005; three females and one male collected from Do-dong, Ulleung Is. (37°28′52?N, 130°54′34?E), 5 m depth, sandy gravel deposits, coll. Sung Joon Song using a light trap, 29 June 2005; three females collected from Moonseom Isl., Jeju Is., 30 m depth, same as type locality, coll. Sung Joon Song, 6 October 2006.
Female. Total body length 1.980 mm (mean 1.693 mm, n=6), measured from the anterior margin of the rostrum to the posterior margin of the caudal rami. Largest width measured at posterior margin of genital-double somite, 0.304 mm. Urosome gradually tapering posteriorly. Cephalothorax bell-shaped, with few integumental sensilla as figured (A, B); posterior and posterolateral margin crenulate with several sensilla. Rostrum (A,B, 3D) prominent, triangular, directed downward, with trifid tip (very weak), with pair of subapical sensilla. Pedigerous somites with a pair of hairs on dorsal surface, crenulate posteriorly as cephalothorax.
Urosome (A, 7A) 5-segmented, comprising P5-bearing somite, genital-double somite and three free abdominal somites. All urosomites with serrate posterior margin except for anal somite.
Genital-double somite (A) as long as wide, with small spinules laterally; urosomites 2 and 3 (genital-double somite) partly fused with transverse serrate surface ridge dorsally and dorsolaterally. P6 (A) represented by long outer seta and one small inner seta on small protuberance.
Anal somite (A, 7A) with concave medial outer margin; ornamented with tiny spinules ventrally. Anal operculum semicircular, with long setules along posterior margin.
Caudal rami (A) with overlapping expanded inner proximal part; about four times as long as wide; each ramus tapering distally, with several spinules on small protuberance laterally. All seven setae located in distal quarter; seta I bare, shortest, setae II–IV bare, and seta VII tri-articulate at base.
Antennule (A) 4-segmented; segment 1 smallest, with outer process; segment 2 with two outer processes; distal one about three times as long as proximal one, each one bearing one seta distally; segment 3 longest, with two lobes distally. Armature formula: 1-[1], 2-[8], 3-[8+ae], 4[11].
Antenna (B) 3-segmented, comprising coxa, allobasis and free endopodal segment. Coxa with spinular row. Allobasis as long as endopod, with several spinules on outer margin and tiny abexopodal seta midlength. Endopod ornamented with spinules along outer margin and one transverse hyaline frill subapically. Lateral armature consisting of two spines; distal armature consisting of two spines, three geniculate setae and one slender seta. Exopod represented by one segment, bearing two bare setae.
Mandible (A) with well-developed gnathobase bearing several multicuspidate teeth distally and one pinnate long seta in dorsal corner; with one spinule row on surface. Palp well-developed, biramous; basis ornamented with a surface setule row and three pinnate setae; endopod with one distal seta; exopod bilobed with three distal bare setae and one subdistal pinnate seta.
Maxillule (B). Praecoxa with few spinules on outer distal corner and posteriorly. Arthrite strongly developed, with one unipinnate inner seta, one naked surface seta and pore, and five distal spines, and one spine and one unipinnate seta subdistally. Coxal endite with one bare seta and one bipinnate seta. Palp 1-segmented, elongated with three outer setae, one small posterior seta, and three distal setae.
Maxilla (C). Syncoxa with several spinules along bilobed outer margin, and a spinular row on anterior surface; and with three endites. Proximal endite with spinules on surface and one multipinnate spine; middle endite with one modified spine fused to segment and two long setae; distal endite with two bare setae and one pinnate seta. Allobasis drawn out into pectinate claw; accessory armature consisting of oneanterior seta and one posterior seta. Endopod represented by small protuberance bearing two long and one short setae.
Maxilliped (D). Syncoxa with spinular rows as figured, with three pinnate setae distally. Basis with spinular row along palmar margin and few spinules along outer margin. Endopod represented by apically curved claw, minutely pinnate in distal third; accessory armature consisting of one long bare seta and two vestigial elements.
P1 (A). Coxa with two rows of spinules on anterior surface. Basis with spinulose process on distal margin between exopod and endopod, with bipinnate seta/spine on inner and outer corners.
Exopod 3-segmented, shorter than endopod; exp-1 with outer spinules, with long inner setules, and one outer bipinnate spine; exp-2 slightly longer than other segments, with one bipinnate outer spine and one small inner spine with long setules at its tip; exp-3 shortest, with two unipinnate long spines and two geniculate setae. Endopod 2-segmented; enp-1 about two times as long as exopod, with spinules along outer margin, with one small bipinnate rod-like seta on proximal third; enp-2 elongate, with outer spinular row, with one pectinate spine and one geniculate seta distally and two small inner setae.
P2-P4 (B, 5A,B) with wider intercoxal sclerites lacking ornamentation. Praecoxae with spinular row anterodistally except P2. Coxae with two spinular rows on anterior surface. Bases wider than long, with spinulose process on distal margin between exopod and endopod; outer distal setae bare, only P2 with a few setules on inner margin. Each rami consisting of 3-segmented exopod and 2-segmented endopod; armature formula of P2-P4 as in .
Table 1. Setal formula of swimming legs of Isocletopsyllus maximus.
P5 (A) with separate baseoendopod and exopod, both covered with long setules laterally. Baseoendopod longer than wide, with long outer basal setae arising from triarticulate setophore. Endopodal lobe extending almost to middle of exopod, with a small process distally, with 5 bipinnate setae. Exopod elongated, about 10 times as long as wide, with three bipinnate outer setae and three bare setae apically.
Male. Total body length 1.78 mm (mean 1686 mm, n=7), measured from anterior margin of rostrum to posterior margin of caudal rami. Largest width measured at posterior margin of cephalic shield, 0.26 mm.
Prosome (B) 4-segmented, comprising cephalothorax and three free pedigerous somites. Posterior margin of cephalothorax and pedigerous somites crenulate with few integumental sensillae as figured. Rostrum as in female.
Urosome (B) 6-segmented, comprising P5-bearing somite, genital somite and four abdominal somites. Posterior margin of all somites with crunulate line dorsally and ventrally, except for anal somite. Caudal rami (B, 7B) sexually dimorphic, without inner expansion, about six times as long as wide, all seven setae similar to that of female.
Antennule (E–H) 7-segmented; subchirocer, with geniculation between segment 4 and 5. Segment 1 ornamented with outer spinules, and with outer process. Segment 2 with two outer processes. Segment 5 longest and swollen. Armature formula: 1-[1 unipinnate], 2-[8 bare+3 pinnate], 3-[5 bare], 4-[2 bare], 5-[7 bare+3 modified+3 pinnate+(1+ae)], 6-[1 bare], 7-[8 bare+acrothek]. Apical acrothek consisting of two long setae and aesthetasc.
Antenna, mouth appendages, and P1, P3, P4 as in female.
P2 (C). Exopod as in female. Endopod modified, 2-segmented; enp-2 modified with four plumose inner setae, one short bipinnate outer seta; with one bipinnate seta and one modified styliform spine apically.
Fifth pair of legs (C). Rami distinct. Baseoendopodal lobe shorter than exopod, with long triarticulate outer basal seta; endopodal lobe with one long inner seta and two distal setae accompanied by a long tube-pore, outer margin with long setules. Exopod about four times as long as wide, with three outer setae and two distal setae.
Sixth pair of legs (B) asymmetric, armature consisting of one curved long spine on cylindrical process.
Etymology
The species name is derived from the Latin maximus, meaning biggest or longest, and refers to the longest body length within the family.
Discussion
Characteristic of the genus Isocletospyllus
The new species herein described from Jeju and Ulleung Islands was attributed to Isocletopsyllus Huys and Lee, Citation1999 by the presence of a crenulated posterior margin of body somites, female 4-segmented antennules, third segment of the female antennule without any posterior modification, male P5 exp with five setae/spines, and sexually dimorphic caudal rami (with a larger length:width ratio than in females). Isocletopsyllus maximus sp. nov. differs from other species of the genus in the male P2 enp-2 with a modified apical spine fused to the distal segment, instead of unilaterally pectinate spines on p4 exp-3. In addition the female of the new species has a caudal ramus with a widened inner expansion, and has a raised spinulose patch on outer margin proximally, while other species in the genus has the patch on the inner proximal margin.
Sexual dimorphism of swimming legs
Huys and Lee (Citation1999) suggested allocating genera Bathycletopsyllus [B. hexarthra] and the genus Cletopsyllus in the family Cletopsyllidae, being composed of three genera: Cletopsyllus Willey, Citation1935 [C. papillifer Willey, Citation1935; C. bacescui Marcus, Citation1976; C. rotundifera Fiers, Citation1986], Retrocalcar Huys and Lee, Citation1999 [R. brattstoemi (Geddes 1981); R. secundus (Nicholls Citation1945); R. sagamiensis (Itô Citation1971)] and Isocletopsyllus Huys and Lee, Citation1999 [I. tertius (Por Citation1964); I. quartus (Soyer Citation1966)]. The species of the family are rarely found worldwide and still unknown are the males of R. secundus, I. quartus and C. papillifera. It would be also notable that only one male of the genus Isocletopsyllus was known to science. Thus detailed description of the sexual dimorphism of I. maximus sp. nov. would help us better understand the taxonomy of the family, especially for the genus Isocletopsyllus. As shown in for the modification of male swimming legs, the third outer seta of p2 enp-2 is transformed to a styliform spine in five species, B. hexarthra, C. rotundifera, C. bacescui, R. brattstroemi, and I. maximus sp. nov. Of these species C. rotundifera has a curved spine and C. bacescui has a similar spine that is not fused at base. In p4 exp of the male three species (B. hexarthra, R. brattstoemi and I. tertius) have modified outer spines, while specimens of the new species from Jeju and Ulleung Islands do not show these modifications ().
Table 2. Modified leg, body length and their recorded localities of cletopsyllid species.
Remarks on Isocletopsyllus species
Representatives of the genus Isocletopsyllus have been recorded only from the Mediterranean. Por (Citation1964) described both sexes of I. tertius from off the Israel Mediterranean coast (on gravel bottom, 35 m deep). Isocletopsyllus tertius is different from the new species in: (1) having a setule row on the inner expansion of caudal rami proximally, (2) antennules without process anteriorly, (3) P1 exo-1 and -2 with long and thin outer spines as in I. quartus (thick spine in I. maximus), (4) male P4 exp-3 with three unipectinate spines, (5) male and femal basal seta of p5 baseoendopod without a setule line along the margins, and (6) male P5 baseoendopod with few spinules along the inner margin.
Isocletopsyllus quartus (Søyer 1966) was recorded from Banyuls-sur-Mer, France by Søyer (1966). The male and female of I. tertius are known, but only the female of I. quartus is known. The latter could not show the sexual dimorphism of the male, which is unexplored as yet. Isocletopsyllus quartus is different from the new species, I. maximus, in (1) caudal ramus with several setules on the inner expansion proximally in I. quartus, while with setules on the outer proximal margin in I. maximus sp. nov, (2) third antennular segment with 10+ae in the female of I. quartus, while 8+ae in I. maximus, (3) endopodal segment of antenna with long setules on the outer margin in I. quartus, while with short setules along the outer margin in I. maximus sp. nov., (4) syncoxa of maxilliped with two setae, while with three setae in I. maximus, (5) basis of mandible with two setae I. quartus, while with three setae in I. maximus, and (6) proximal endite of maxilla with three setae in I. quartus, but one seta in I. maximus sp. nov.
Differential diagnosis
Isocletopsyllus maximus sp. nov. can be separated from its congeners on the basis of the following aspects that are differential characters of the species: (1) elongated body in female (1.98 mm) and male (1.78 mm) (compared to 1.15 mm in female of I. quartus; 1.52 mm in female and 1.30 mm in male of I. tertius), (2) caudal ramus with a spinular field on a small outer protuberance proximally, (3) well-developed outer setule line on syncoxa of the maxilla, (4) P1 exp-2 with a short rod-like inner seta, (5) P2 enp-2 with a short outermost spine, P2 enp-2 of male with a styliform spine derived from a transformed distal inner seta, (6) male P4 exopod without modified outer spines, (7) P5 exp very long and narrow with a very short second outer seta, and (8) male baseoendopod with well-developed tube-pores, especially distally.
Biogeography
Representatives of the genus Isocletopsyllus have a comparatively elongated body and live on shallow subtidal gravel or in sandy bottoms, from 10 to 45 m depth. So far only two species of the genus collected from the Mediterranean Sea, I. quartus, have been collected along the coasts of Banyuls-sur-mer, France, and I. tertius have been collected from off the Israeli coast. The discovery of I. maximus sp. nov. in Korean waters (eastern and southern islands) would expand the range of the hitherto known biogeography for representatives of the genus Isocletopsyllus, from the Mediterranean to the Far East of Asia.
Additionally, as shown in , R. sagamiensis was also reported from the Pacific Ocean (collected from the Sagami Bay, 80 m depth) as I. maximus. Bathycletopsyllus hexarthra, C. bacescui, C. rotundifera and R. secundus were reported from the Indian Ocean; especially B. hexarthra were collected from the deepest bottom (460 m). Cletopsyllus papillifer and R. brattstoemi were reported from the Atlantic Ocean.
Key to the female species of the genus Isocletopsyllus
1. First antennular segment with outer process ------------------------------------------------------ 2First antennular segment without outer process; caudal ramus about six times as long as wide ------------------------------------------------------------------------------------- I. tertius (Por Citation1964)
2. Caudal ramus without expanded inner board; about 10 times as long as wide ------------------------------------------------------------------------------------------------- I. quartus (Soyer Citation1966)Caudal ramus with expanded inner board; about four times as long as wide ------------------------------------------------------------------------------------------------------------- I. maximus sp. nov.
Acknowledgements
We thank Mr. Jae Wan Choi for his support in collecting samples on Jeju Island. This work was supported by the Korea Research Foundation Grant funded by the Korean Government (MOEHRD)"(KRF-2005-070-C00124) and partly by a grant entitled ‘Origin of Biological Diversity of Korea: Molecular Phylogenetic Analyses of major Korean Taxa’ funded by the National Institute of Biological Resources, Korea awarded to UWH.
Related Research Data
References
- Chang , CY. 2008 . Brackish-water copepods of the family Tachidiidae (Copepoda: Harpacticoida) from South Korea . Korean J Syst Zool. , 24 ( 2 ) : 229 – 240 .
- Chang , CY and Song , SJ. 1995 . Marine harpacticoid copepods of genus Eudactylopus (Harpacticoida, Thalestridae) in Korea . Korean J Syst Zool. , 11 ( 3 ) : 379 – 388 .
- Chang , CY and Song , SJ. 1997a . Two new thalestrid harpacticoids (Copepoda,Harpacticoida, Thalestridae) from Korea . Korean J Biol Sci. , 1 ( 2 ) : 297 – 304 .
- Chang , CY and Song , SJ. 1997b . Marine harpacticoids of genus Parathalestris (Copepoda, Harpacticoida, Thalestridae) from Korea . Korean J Syst Zool. , 13 ( 3 ) : 221 – 231 .
- Fiers , F. 1986 . Amsterdam Expeditions to the West Indian islands, Report 48. Harpacticoid copepods from the West Indian islands: Laophontidae (Copepoda, Harpacticoida) . Bijdr Dierk. , 56 : 132 – 164 .
- Geddess , DC. 1981 . Marine biological investigations in the Bahamas 21. A new species of Cletopsyllus (Copepoda, Harpacticoida) . Sarsia. , 66 ( 4 ) : 287 – 291 .
- Huys , R , Gee , JM , Moore , CG and Hamond , R. 1996 . Marine brackish water harpacticoid copepods. Part 1 . Synopses of the British Fauna (New Series) , 51 : 1 – 352 .
- Huys , R and Lee , W. 1999 . On the relationships of the Normanellidae and the recognition of Cletopsyllidae grad. nov. (Copepoda, Harpacticoida) . Zool Anz. , 237 : 267 – 290 .
- Itô , T. 1971 . A new species of the genus Cletopsyllus from Sagami Bay (Harpacticoida) . Annot Zool Japon. , 44 ( 2 ) : 117 – 124 .
- Je , J-G , Koo , BJ , Lee , H-G , Kim , B-I , Shin , S-H , Lee , S-W and Lee , J-H. 2002 . Habitats and zoobenthic species diversity in the coast of Jeju Island, Korea: as a baseline study for conserving coastal and marine biological diversity . Underwater Sci Technol. , 3 ( 1 ) : 7 – 109 .
- Marcus , A. 1976 . Contributions to the study of the genus Cletopsyllus Willey 1935 (Copepoda, Harpacticoida) from the Indian Ocean . Trav Mus Hist Nat "Gr Antipa" , 17 : 39 – 49 .
- Nam , E-J and Lee , W. 2006 . Two new species of the genus Heteropsyllus (Crustacea, Copepoda, Harpacticoida) from Jeju Island, Korea and Devon, England . J Nat Hist. , 40 ( 29–31 ) : 1719 – 1745 .
- Nicholls , AG. 1945 . Marine Copepoda from western Australia. III. Littoral harpacticoids from Port Denison . J R Soc West Aust. , 29 : 1 – 16 .
- Por , FD. 1964 . A study of the Levantine and pontic Harpacticoida (Copepoda Crustacea) . Zool Verh. , 64 : 1 – 128 .
- Song , SJ and Chang , CY. 1993 . Eight harpacticoid species of Harpacticidae (Copepoda, Harpacticoida) from Korea . Korean J Syst Zool. , 9 ( 2 ) : 203 – 220 .
- Song , SJ , Yun , SG and Chang , CY. 1999 . New records on three harpacticoid copepods associated with marine macroalgae in Korea . J Fish Sci Tech. , 2 ( 2 ) : 189 – 198 .
- Song , SJ , Kim , W and Chang , CY. 2001 . Redescription of two thalestrid copepods, Phyllothalestris sarsi Sewell, 1940 and Dactylopusia falcifera Willey, 1935 (Copepoda, Harpacticoida, Thalestridae) in Korea . Korean J Syst Zool. , 17 ( 2 ) : 229 – 243 .
- Song , SJ , Kim , W and Chang , CY. 2003 . A new species of Parastenhelia (Copepoda: Harpacticoida: Parastenheliidae) from Korea . Zool Sci. , 20 : 221 – 228 .
- Soyer , J. 1966 . Copepodes Harpacticoides de Banyuls-sur-Mer. 3. Quelques formes du coralligene . Vie Milieu. , 17 ( 1-B ) : 303 – 344 .
- Willey , A. 1935 . Harpacticoid Copepoda from Bermuda. Part II . Ann Mag Nat Hist. , 10 ( 15 ) : 50 – 100 .