Abstract
Estimating the habitat potential of inland forest patches for birds requires the modeling of species–area relationships, or relationships between habitat size and numbers of bird species in each patch. The accurate estimation of species–area relationships significantly reduces the effort required to recognize the number of species living in each patch. The objective of this study was to estimate the relationship between forest patch size and bird species diversity in Dangjin County, in northwest South Korea, based on the sizes of inland forest patches. KOMPSAT-2 images were obtained and ortho-rectified to construct a map of the target forest patches. The numbers of birds per patch were surveyed four times: August 2008, September 2008, February 2009 and May 2009. Regression models were derived to explain the relationships between the numbers of bird species and patch size. A model that was derived using data from all four observation periods had the highest coefficient of determination (R 2). According to these models, the numbers of bird species at first increased linearly with increasing patch size; however, the curve then plateaued. Our model including observations from four seasons will be useful for estimating the numbers of bird species in other inland forest patches in South Korea.
In island biogeography, habitat area and isolation are considered to be the primary determinants of the number of biological species. Since MacArthur and Wilson (Citation1967) first proposed the theory of island biogeography, many biogeographers have investigated the relationship between islands and the number of biological species present on islands, and concluded that the area of an island has a large influence on the number of species present on the island (Rafe et al. Citation1985; Michael et al. Citation2002).
As various land covers and land use types can be present on an island, resources for the survival of species are not necessarily dependent only on the size of the island. However, it is expected that the larger an island is, the more species it would attract as larger islands generally have more resources necessary for the survival of species than smaller islands.
Frank Preston (Citation1962), an early investigator of species–area relationships, divided these relationships into two types: samples (a census of a contiguous habitat, also called ‘mainland’ species–area relationships) and isolates (a census of discontiguous habitats, such as islands, also called ‘island’ species–area relationships) (http://en.wikipedia.org/wiki/Species-area_curve). Using Preston's concepts, it may be possible to apply species–area relationships to inland areas. A forest patch is a landscape element that can potentially substitute for an island habitat when investigating species–area relationships. In landscape ecology, a forest is recognized as a small patch in a whole landscape and is interactively related to other patches. Each forest patch can therefore be ecologically connected or isolated depending on the configuration or size of the matrix (Jongman and Pungetti Citation2004; Platt and Lill Citation2006). Furthermore, the size of forest patches and the distance between forest patches are sometimes associated with the size of an island and the degree of isolation in the theory of island biogeography. For birds, however, the isolation of forest patches does not appear to play a major role, due to birds’ ability to fly long distances. Instead of that, larger forests could contain more resources such as food and cover that are attractive to birds. Furthermore, more species are predicted to be present in larger forest patches, which harbor protected inner regions and buffer zones to mitigate environmental pressure from the outside.
The objective of this study was to estimate the diversity of bird species depending on forest size for evaluating whether there is a relationship between the number of species and the size of the forest patch in inland area. This relationship can be an indicator for estimating or managing the level of biodiversity of a forest patch in an urban area.
Materials and methods
Study area
Dangjin is a rapidly developing city in South Korea located at about 36°53′34″N 126°38′10″E; total population has gradually increased, and 11,639 moved to this city in 2007 to get a job (Dangjin county office Citation2009). Two hundred and fifty companies move into Dangjin every year. Due to the rapid pace of urbanization, many forest patches have been destroyed or divided into smaller patches, or converted to other land uses. Even though Dangjin has developed rapidly and its population has increased significantly, many forest patches are still densely distributed because they occur in low-elevation areas and/or on slopes. Each forest patch is potentially a forest habitat for birds, and a forest patch is a continuous one fragmented by any road or artificial one. The term ‘patch’ is a landscape-ecological term, denoting a unique grain within a landscape (Begon et al. Citation2008). We chose to investigate 21 forest patches distributed in Dangjin county in northeast Chungnam province, South Korea, to identify the species–area relationships for birds in these patches. The forest patches chosen were near the national road across the county, while one forest patch each on Ami and Dabul mountains was included. Most of the other forest patches were satellite patches near much larger forest patches ().
Figure 1. Twenty-one forest patches investigated in Dangjin county.
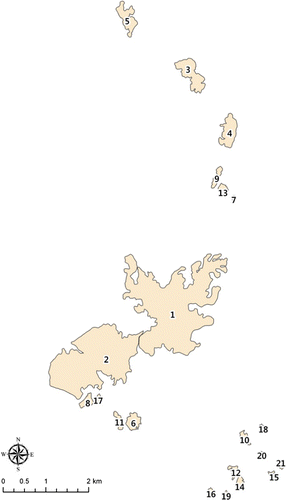
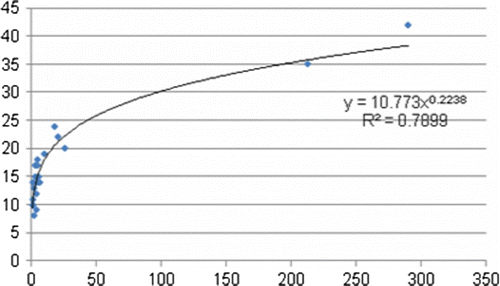
Figure 2. Species–area curve derived from integrating the four-season bird survey data.
The forest patches selected in this study were of different sizes (). Different patch sizes could potentially result in different types of birds in each patch. This landscape-ecological perspective can be used to model the species–area curve relationships for birds. In terms of the targets for modeling, we used two patches on two large mountains that were larger than 2 km2, and three forest patches that were smaller than 1 ha in size.
Table 1. Area of forest patches in the study area.
Bird survey data
Birds were surveyed in 21 forest patches using a line transaction method. Bird species were classified by naked eye observation and their vocalizations. Binoculars (Nikon 8x30) were used for bird observations. Bird surveys were carried out by walking through each forest at a speed of about 2 km per hour, and birds within 25 m on either side of the observer were counted. The number of birds per forest patch was surveyed four times: in August 2008, September 2008, February 2009 and May 2009. Two additional forest patches were selected and bird species in each patch were observed twice in February 2010 and May 2010 to evaluate the validity of the species–area relationship models derived from the survey of 21 forest patches.
Species–area curves
This species–area relationship can be formalized as a power function as follows (Connor and McCoy Citation1979; Crawley and Harral Citation2001):
which is linearized by logarithmic transformation to give
where S is the number of bird species, A is the size of islands, and c and z are constants that can be estimated by regression analysis. The term ‘c’ generally reflects the influence of geographic variation on species diversity, and the z-value is the slope obtained by linear regression when the species–area curves are log-transformed. The slope, z, depends on the overlap in the species composition of the localities used to draw the curve. When localities share many species, there is a ‘slow’ increase in the number of species with area, and z will be relatively small. Alternatively, when localities share just a few species, there is a ‘faster’ increase in the number of species with area, and z will be relatively high (Michael et al. Citation2002). Rosenzweig (Citation1995) and after him Brown and Lomolino (Citation1998) established three different categories of species–area curves: between biogeographical provinces, within biogeographical provinces and of islands or isolated habitat patches. The typical values of z are 0.5–1, 0.1–0.2 and 0.2–0.5, respectively (Triantis et al. Citation2003).
Estimating species number depending on the size of forest patches
To estimate the number of bird species that immigrate or dwell in a forest according to the size of the forest patch size, we performed ortho-rectification of high spatial resolution imagery, classified vegetation types by image interpretation and field surveys, monitored bird species, and estimated species–area curves.
Satellite images were analyzed and field surveys were conducted simultaneously to characterize the plant communities present in the study area. High spatial resolution images were obtained using the KOMPSAT-2 sensor (Korean Multi-purpose Satellite) operated by the Korea Aerospace Research Institute (KARI) at Daejeon, South Korea. KOMPSAT-2 images with a resolution of 1 m were collected twice on 16 February 2007 and 10 June 2007. The images were taken in four channels ranging from 450 nm to 900 nm (blue: 450–520 nm; green: 520–600 nm; red: 630–690 nm; NIR: 760–900 nm). A digital elevation model (DEM) and coordinate data were used for ortho-rectification of the KOMPSAT-2 images. The coordinate data were collected from a digital topographic map produced by the National Geographic Information Institute (NGII). The ortho-rectification of KOMPSAT-2 image was performed using Leica Photogrammetry Suite 9.1 software (Leica geospatial imaging, Norcross, Ga.)
Boundaries of the vegetation cover were digitized on the ortho-rectified KOMPSAT-2 images on a computer. Polygons describing vegetation communities were vectorized using an on-screen digitizing method, and classified into three vegetation groups: deciduous forest, coniferous forest, and mixed stand forest, by visual investigation. Boundary data were overlapped with KOMPSAT-2 images using the Arc Map 9.3 program (ESRI, California, USA), and then printed out to use as a base map for mapping dominant vegetation communities within the target forest patches. Through field surveys, boundaries of the dominant vegetation communities were mapped on computer using geographic information system (GIS) technology. Polygons representing the extent of dominant vegetation communities were subdivided by using the data editing module in Arc Map 9.3, and information on the communities was input into an attribute table.
Results
Bird species survey
A total of 57 bird species were observed in 21 forest patches (). Ami mountain, the largest mountain in Dangjn county, contained 42 bird species, while patch No. 16 with a patch size of 1.60 ha was found to have only eight species. Many different bird species were observed on Ami mountain. Tits, Parus spp., the long-tailed tit, Aegithalos caudatus, the rustic bunting, Emberiza rustica, and the yellow-throated bunting, Emberiza elegans, were often observed at the forest boundary, while species including the Indian cuckoo, Cuculus micropterus, and thrushes, Turdus spp., were observed in inner forest patches on the mountain. Birds including great tit, Parus major, brown-eared bulbul, Hypsipetes amaurotis, and rufous turtle dove, Streptopelia orientalis, which were also found in patches less than 1 ha in size, e.g. patch numbers 19, 20 and 21, indicated edge species preferring forest boundaries to an inner space ().
We performed seasonal monitoring of birds in this study because of the immigration in spring of summering birds. These summering birds start to emigrate from about late August, and are all gone by October. In September and October, resident birds and a few migrant birds were observed in the forest patches. In the winter season from November to February, wintering birds replaced the migrant birds in forest patches.
Mountain birds, including summering birds, were observed during the surveys performed in May and August. In May, resident and summering birds as well as migrant birds were observed. The major summering birds observed were the dollarbird, Eurystomus orientalis, black-naped oriole, Oriolus chinensis, eastern crowned willow warbler, Phylloscopus coronatus, hoopoe, Upupa epops, and migrant birds included the rufous-tailed robin, Luscinia sibilans, olive-backed pipit, Anthus hodgsoni, and chestnut bunting, Emberiza rutila. In August, many species of resident birds such as the rufous turtle dove, Streptopelia orientalis, vinous-throated parrotbill, Paradoxornis webbianus, great tit, Parus major, marsh tit, Parus palustris, brown-eared bulbul, Hypsipetes amaurotis, and tree sparrow, Passer montanus, were found in forest patches. Moreover, summering bird species such as White's thrush, Turdus dauma, the blue-and-white flycatcher, Cyanoptila cyanomelana, and the pale thrush, Turdus pallidus, were observed. When the season changed from summer to autumn, the number of the summering birds decreased and migrant birds such as the yellow-browned wabler, Phylloscopus inornatus, appeared.
During February, most wintering birds prepare to emigrate from the Korea peninsula. Representative wintering birds observed in February were the rustic bunting, Emberiza rustica, hawfinch, Coccothraustes coccothraustes, brambling, Fringilla montifringilla, and goldcrest, Regulus regulus.
Models relating bird species to forest patch size
Equation Equation1 defines the relationship between forest patch size and the number of bird species. Using this equation, we calculated the area–species relationship using a power function. Equations Equation2–Equation5 are the area–species relationships derived from surveys in August (equation Equation2), September (equation Equation3), February (equation Equation4) and May (equation Equation5). Equation Equation6 reflects the observations from all four seasons.
The models had reasonable credibility, and we found a significant relationship between patch size and species diversity across the year with the highest coefficient of determination (R 2) and significant coefficients (). The R 2 values from the models for August and September were 0.47 and 0.48, respectively, while the models for May and February had much higher R 2 values of 0.74 and 0.68, respectively. Many summering birds start to migrate in August and September, but at this time it is still early for migrant birds or wintering birds to settle down. However, in May summering birds settle down while in February wintering birds are not yet ready to migrate. A more credible model was obtained from integration of data from all four seasons, although it is clear from our data that the species–area relationship of birds is affected to some extent by seasonal variation.
Table 2. Coefficients and significance for the regression models in this study.
The z-value, which determines the rate of increase in the number of species with increasing area, showed wide variation depending on the season. In this study, the z-values for August and May were 0.27 and 0.28, respectively, while the z-values for September and February were 0.31 and 0.22, respectively.
Simple investigation of verifying the species–area relationships for birds
To investigate the validity of the species–area relationships obtained in this study, two forest patches, occupying 64 ha and 92 ha, were surveyed in February and May 2010. In February, 13 species of birds were surveyed in the 64-ha forest patch, and 14 species of birds were observed in the 92-ha forest patch, while the species–area curve for February (equation Equation4) estimated nine species for the 64-ha forest, and 10 species for the 92-ha forest. This indicates that the species–area curve for February underestimated the number of species. The survey in May found 17 species in the 64-ha forest and 16 species in the 92-ha forest, while the model predicted 15 species for the 64-ha forest and 17 species for the 92-ha forest. The model for May therefore showed a good fit between the number of observed species and the number of estimated species. However, this model slightly underestimated the number of species according to forest patch size.
Discussion
Different preferences of forest bird species for food or cover were estimated to cause different frequencies in forest patches: a higher frequency in a large patch, a higher frequency in a small patch, and frequency not dependent on the size of forest. Landscape perspectives, land use types and other characteristics appearing in a landscape mosaic can be a good inducement sometimes, while some of them can be a barrier for birds.
Though there was seasonal variation, the general trend was that the number of species increased as the forest patch size increased. However, after a certain point, the curve of the relationship between the number of species and forest patch size plateaued as the patch size increased. The models derived in this study have aspects similar to those reported for species–area relationships, but probably underestimate the number of species in forest patches. All the curves, except for that based on survey data from September, underestimated the increase in the number of species with increasing patch size. Through the simple investigation of verifying the species–area relationship, we could also recognize that the relationship of bird species to forest size had a power curve, not a linear relationship. The models suggested in this study were related to other approaches in island biogeography. The models analyzed in other studies showed a power curve as we suggested in this study. In Particular, the z-values are consistent with the previously reported z-values for bird species in forest patches of 0.23 (Woodhouse Citation1981), 0.27 (Moore and Hooper Citation1975), and 0.13 (Rafe et al. Citation1985).
In the case of small sizes of forest patches, it was difficult to determine the relationship between the number of species and forest patch size in some of the small forest patches. For example, the graphs derived from the surveys in August (equation Equation2) and September (equation Equation3) indicated that some small patches had a small number of species and other small patches had large number of species, while equation Equation4 showed a similar number of species in forest patches of different sizes. Such a phenomenon is known as the small island effect, which means that there is no increase in the number of species with increasing island size (Triantis et al. Citation2003). The small island effect can be estimated to be derived from a variety of condition within forest patches including vegetation types, shape perspectives and so on. Our model, including observations from four seasons, will be useful for estimating the number of bird species in other inland forest patches, and also the relationship between bird species number and forest patch size would be an important indicator of how many species could live in a certain size of forest, and of what we should do to increase the number of bird species in inland forest patches.
Acknowledgements
This research was supported by the Yeungnam University research grants in 2009
Related Research Data
References
- Begon , M , Townsend , CR and Harper , JL. 2008 . Ecology: from individuals to ecosystems , 167 Oxford : Blackwell .
- Brown , JH and Lomolino , MV. 1998 . Biogeography , Sunderland, MA : Sinauer associates .
- Connor , EF and McCoy , ED. 1979 . The statistics and biology of the species–area relationship . Am Nat. , 113 : 791 – 833 .
- Crawley , MJ and Harral , JE. 2001 . Scale dependence in plant biodiversity . Science. , 291 : 864 – 868 .
- Dangjin county office . 2009 . Statistical yearbook . p. 55 – 62 .
- Jongman , R and Pungetti , G. 2004 . Ecological networks and greenways: concept, design, implementation , 7 – 33 . Cambridge : Cambridge University Press .
- MacArthur , RH and Wilson , EO. 1963 . An equilibrium theory of insular biogeography . Evolution. , 17 : 273 – 387 .
- MacArthur , RH and Wilson , EO. 1967 . The theory of island biogeography , Princeton, NJ : Princeton University Press .
- Michael , DC , Vázquez , DP and Sanders , NJ. 2002 . Species–area curves, homogenization and the loss of global diversity . Evol Ecol Res. , 4 : 457 – 464 .
- Moore , NW and Hooper , MD. 1975 . On the number of bird species in British woods . Biol Conserv. , 1 : 2010 – 207 .
- Platt , A and Lill , A. 2006 . Occurrence of breeding bird species in urban parks: effects of park structure and broad-scale variables . Urban Ecosys. , 9 : 83 – 97 .
- Preston FW . 1962 . The canonical distribution of commonness and rarity: Part I . Ecology 43 : 185 – 215 and 431–432 .
- Rafe , RW , Usher , MB and Jefferson , RG. 1985 . Birds on reserves: the influence of area and habitat on species richness . J Appl Ecol. , 22 : 327 – 335 .
- Rosenzweig , ML . 1995 . Species diversity in space and time , New York, NY : Cambridge University Press .
- Sylvén , M. 1978 . Interspecific relations between sympatrically wintering common buzzards Buteo buteo and rough-legged buzzards Buteo lagopus . Ornis Scand. , 9 : 197 – 206 .
- Triantis , KA , Mylonas , M , Lika , K and Vardinoyannis , K. 2003 . A model for the species-area-habitat relationship . J Biogeogr. , 30 : 19 – 27 .
- Woodhouse , MEJ. 1981 . A description of a species-area relationship: a theoretical analysis and a discussion of its broader implications [M.Sc. thesis] , University of York .