
ABSTRACT
Previous research addressing motivational effects on cognitive control adaptations primarily manipulated external rewards. In the present study, we examined whether achievement motivation, reflecting intrinsic motivation, modulates cognitive control adaptations. In each of two experiments, students were divided into a high achievement motivation group (HAM; Experiment 1: N = 36; Experiment 2: N = 39) and a low achievement motivation group (LAM; Experiment 1: N = 30; Experiment 2: N = 39) on the basis of the Achievement Motivation Scale. Cognitive control adaptations were assessed by sequential congruency effects (SCEs) in Flanker tasks. Using a standard Flanker task in Experiment 1 and examining response times, the HAM but not LAM participants showed evidence of cognitive control adaptations. Because SCEs in a standard Flanker task may reflect lower-level, bottom-up processes rather than cognitive control adaptations, we used an adapted version of the Flanker task in Experiment 2 that controlled for the potential influence of such processes. Experiment 2 again revealed evidence of SCEs in the HAM but not LAM participants. These results suggest that cognitive control adaptions may be modulated by achievement motivation. Such potential modulation could be taken into account when evaluating results of previous and future fundamental and applied studies on cognitive control adaptations.
Introduction
Executive or cognitive control processes enable the pursuit of goals in the face of continually changing and potentially distracting environmental stimuli (Diamond, Citation2013). These cognitive processes are crucial for many behaviours that are of clinical and societal relevance (e.g. Young et al., Citation2009) and are positively associated with (developing) physical and mental health (Diamond, Citation2013), such as mathematical and reading skills in early childhood and social functioning (Blair & Razza, Citation2007; Carlson & Moses, Citation2001). Reversely, compromised cognitive control abilities have a detrimental impact on society, being conducive to, for example, alcohol abuse, criminal versatility, and antisocial behaviour (e.g. Chamorro et al., Citation2012).
One important executive function, which is the focus of the present study, concerns the ability to prevent stimuli from interfering with performing a target task, also termed interference control (Friedman & Miyake, Citation2004; Nigg, Citation2000; Stahl et al., Citation2014). Interference control is frequently studied using the Flanker task (Eriksen & Eriksen, Citation1974). In this task, participants have to respond according to some feature of a centrally-presented stimulus. On incongruent trials, the central stimulus is surrounded by stimuli that prompt a response that conflicts with the response demanded by the central stimulus. On congruent trials, flanking and central stimuli implicate the same response. On incongruent, but not congruent trials, the participant must actively suppress attention and behavioural responding to the conflicting flanking stimuli, thus requiring interference control. This difference in interference control demand is reflected in shorter response times (RTs) and a higher response accuracy for congruent than incongruent trials.
Cognitive control processes in general, and interference control in particular, have been shown to be flexibly adjustable to meet current demands. One type of such adjustments are moment-to-moment cognitive adaptations as evidenced by sequential congruency effects (SCEs; see Duthoo, Abrahamse, Braem, Boehler, & Notebaert, Citation2014, for a review). Within the framework of a Flanker task, SCEs take the form of the difference in RT and accuracy between congruent and incongruent trials being smaller after an incongruent trial compared to after a congruent trial. This effect is also known as the Gratton effect (Gratton, Coles, & Donchin, Citation1992) and can reflect more rapid and accurate responding to an incongruent trial that is preceded by another incongruent trial (iI trial) compared to an incongruent trial that is preceded by a congruent trial (cI trial). Alternatively, or in addition, it may reflect faster and more accurate responding to a congruent trial that is preceded by another congruent trial (cC trial) compared to a congruent trial that follows an incongruent trial (iC trial).
In principle, SCEs can be explained in a number of different ways that may or may not assume top-down, cognitive control adaptation (e.g. Egner, Citation2007). A first explanation is the conflict-monitoring account (Botvinick, Braver, Barch, Carter, & Cohen, Citation2001). On this view, participants continuously monitor the stream of information for conflict. Upon detecting a conflict, such as on an incongruent trial in a Flanker task, interference control processes are up-regulated implicating enhancement of selective attention to the relevant central stimulus. Instead, after non-conflict (congruent) trials attention is widened (down-regulated control) to include non-target but congruent flanking stimuli. Attending the non-target stimuli may be beneficial for responding to the target stimulus in this case because all stimuli implicate the same response. These adaptations are assumed to be based on a reactive, relatively short-lived and automatic process (Egner & Hirsch, Citation2005). However, a second approach is the so-called repetition-expectancy account that was put forward by Gratton et al. (Citation1992). Participants are assumed to have a general expectation that two successive trials will be of the same type (congruent or incongruent), even though this might objectively not be true. After an incongruent trial, the participant's attention is narrowed to the central stimulus in anticipation of the next (incongruent) trial. Reversely, attention is widened after a congruent stimulus in anticipation of the next (congruent) trial. This type of adaptation is assumed to reflect a proactive process that takes more time to develop, requires more persistence, and is more strategic than is the case for the adjustment according to the conflict-monitoring account (Duthoo et al., Citation2014).
Each of the two preceding explanations assumes a top-down, cognitive control adaptation. However, the feature-integration or feature-priming account (e.g. Hommel, Citation2004; Mayr, Awh, & Laurey, Citation2003) explains SCEs in terms of more basic, bottom-up processes that are induced by a confound implied in a standard Flanker task. Specifically, cC and iI trial pairs implicate an exact repetition of flanker and central stimuli and responses, or a full alternation of these stimuli and responses. Instead, cI and iC trials implicate a partial repetition of stimuli (either flankers or central stimulus) and responses. As partial repetitions result in slower responses than exact repetitions and full alternations (Hommel, Proctor, & Vu, Citation2004), the classic SCEs as implied in the Gratton effect could, in principle, be explained by low-level, feature-based processes.
Cognitive control and cognitive control adaptation effects have been found to be enhanced by motivational manipulations consisting of introducing primary or secondary external rewards in most but not all studies (e.g. see Braem, Verguts, Roggeman, & Notebaert, Citation2012; Chiew & Braver, Citation2014, Citation2016; Soutschek, Stelzel, Paschke, Walter, & Schubert, Citation2015, for supporting evidence, and van Steenbergen, Band, & Hommel, Citation2009, for evidence of impaired cognitive control induced by reward). Evidence of negative effects of reward on cognitive control adaptation has been suggested to reflect the affective value of rewards (van Steenbergen et al., Citation2009). Accordingly, a conflict (incongruent) trial triggers a negative emotion (e.g. Dreisbach & Fischer, Citation2012a), which in turn motivates enhanced cognitive effort, but a reward may counteract this motive by inducing a positive emotional state. However, motivational influences induced by external rewards and emotional influences on cognitive control may reflect independent mechanisms (Chiew & Braver, Citation2011; Dreisbach & Fischer, Citation2012b). Moreover, individual differences in the motivating effects of external reward may be an additional factor modulating the effect of reward on cognitive control (Braem et al., Citation2012), but much more research on the topic is needed.
The purpose of the present two experiments was to further examine motivational and emotional influences on cognitive control adaptations. In general, research on such influences is important given the already noted large impact of cognitive control abilities on healthy functioning in many domains of daily life (e.g. Diamond, Citation2013). Variables that may modulate the flexible use of such abilities may be informative for developing tools to encourage a more efficient use of cognitive control resources. However, rather than examining external motivational variables, and possible corresponding emotional effects that were primarily examined in previous research, we focussed on an internal motivational variable in the present study. Specifically, we examined the effect of individual differences in achievement motivation on cognitive control adaptations as measured in a Flanker task.
Achievement motivation is a concept from personality psychology and refers to the extent to which an individual enjoys performing challenging cognitive tasks. This enjoyment is associated with the capacity to anticipate positive affects while performing such tasks (e.g. Nygård & Gjesme, Citation1973). Individuals with a high achievement motive base their self-regard on successfully employing and advancing their skills (Atkinson, Citation1964). These features implicate a strong intrinsic motivation to perform cognitively challenging tasks and to show a strong task engagement. This motivation could be linked to emotional consequences that in turn could affect cognitive control adaptation in a number of different ways. For example, following the conflict-monitoring account, the positive emotional state that individuals with a high achievement motivation (HAM) anticipate and experience when performing a conflict task may counteract the negative emotions elicited by incongruent trials, thereby reducing the felt need to initiate cognitive adjustments (e.g. see Schuch, Zweeings, Hirsch, & Koch, Citation2017; van Steenbergen et al., Citation2009, Citation2010). On that view, HAM individuals will show weaker SCEs compared to low-achievement (LAM) participants. The opposite prediction, that HAM participants display stronger SCEs compared to LAM participants, could be derived from previous research suggesting that cognitive control adaptations are based on the positive emotion associated with successfully resolving a conflict trial rather than the negative feeling associated with encountering a conflict (e.g. Braem et al., Citation2012; Schouppe et al., Citation2015). Specifically, resolving a conflict trial may be intrinsically rewarding and temporarily enhance the participant's motivation. Assuming that this intrinsic reward is larger for HAM than LAM participants, one may expect the former participants to display larger SCEs than the latter. After an incongruent trial and compared to LAM participants, HAM participants will experience a stronger motivation to repeat the cognitive process of enhanced selective attention that was used on that trial. This enhanced selective attention is conducive to fast correct responding on a subsequent incongruent trial. However, relative to after a congruent trial, which, if anything, is associated with adopting a broader attentional window, the enhanced selective attention is suboptimal for fast correct responding on a subsequent congruent trial.
Experiment 1
In the first experiment, we assessed the achievement motivation in a large group of students. From this group we selected subgroups of participants with the highest and lowest achievement motivation scores. All participants then performed a standard Flanker task to assess SCEs.
Method
Participants
The participants were 133 students from Northwest Normal University. They all filled in the Achievement Motivation Scale (AMS; see below). The AMS score was used for selecting and grouping participants into a high achievement motivation group (HAM; ∼ top 27% of the AMS scores) and a low achievement motivation group (LAM; ∼ bottom 27% of the AMS scores). Five students from the LAM group did not complete the Flanker task. The students in the HAM group (n = 36; 17 men; 19 women) had a mean AMS score of M = 16.75 (SD = 7.77; range: 8−33) and a mean age of M = 21.25 (SD = 2.41). The corresponding values for the students in the LAM group (n = 30; 9 men; 21 women) were M = 1.63 (SD = 5.24; range: −11−7) and M = 20.50 (SD = 1.93). The two groups differed significantly in AMS score, F(1, 64) = 82.25, p < 0.001, , but not in age, F(1, 64) = 1.90, p = 0.17,
. When we compare these mean AMS scores to those obtained from previous studies (e.g. Hagtvet & Zuo, Citation2000; Man, Nygård, & Gjesme, Citation1994, implicating a mean AMS score across studies in the range of: 8−20), the present AMS scores suggest that especially the LAM group showed a marked low achievement motivation, rather than that the HAM participants presented with a particularly high achievement motivation. All participants were right-handed and had a normal or corrected-to-normal visual acuity. The experiment was approved by the university ethics committee. The students participated voluntarily and signed an informed consent form. All participants received a small remuneration consisting of a notebook or pen after task completion.
AMS
We employed a Chinese version (Ye & Hagtvet, Citation1992) of the AMS (Gjesme & Nygård, Citation1970) to measure achievement motivation. The capacity to anticipate positive affects in achievement situations is assessed with 15 items (Ms items). Fifteen other items assess the capacity to anticipate negative affects in these situations (Mf items). Each item is answered on a 4-point Likert scale, ranging from 1 = completely disagree to 4 = completely agree. The total AMS score was computed by subtracting the total score on the Mf items from the total score on the Ms items, with a higher score reflecting a stronger achievement motivation. The coefficient of internal consistency was close to acceptable (Cronbach's α = 0.68).
Flanker task
The Flanker task was programmed in Eprime and commenced with a 500-ms presentation of a small fixation cross, presented in the centre of the screen against a grey background. The fixation cross was followed by an empty grey screen that was presented for a random duration between 300 and 500 ms. Subsequently, one of four different strings of letters was displayed for a maximum duration of 1500 ms or until a response was made (whichever came first), followed by a 1000-ms empty grey screen. The next trial started immediately thereafter. The letter string either consisted of the letters HHHHH or KKKKK (congruent trials) or the letters HHKHH or KKHKK (incongruent trials). The participant was required to respond according to the identity of the middle letter of the string. Using a standard keyboard, the participant had to press “1” with the index finger of the left hand in case of the letter H, and “0” with the index finger of the right hand in case of the letter K. The participant was asked to respond as quickly and accurately as possible and first completed one or more 16-trial practice blocks, each consisting of eight quasi-randomly intermixed congruent and incongruent trials, until correctly responding on at least 85% of the trials. The actual task was initiated immediately thereafter. The main task consisted of four 33-trial blocks. The trials were quasi-randomly presented (for each participant the same sequence) in such a way that, across the task (= 128 trial pairs), there were 32 trials of each of the cC, cI, iC, and iI trial pairs. The participant could have a short break between each trial block and the task lasted approximately 15 min.
Procedure
All initial 133 students first filled in the AMS. The students that were selected from this larger pool based on their relatively high or low AMS score performed the Flanker task 3−4 days after completion of the AMS. Both the questionnaire and the task were completed in a quiet laboratory room. The Flanker task was performed in small groups of 4−5 students and each student had a separate computer to do the task.
Data analysis
The mean RT, based on correct trials and RTs > 100 ms, and proportion of trials with a correct response obtained from the Flanker task were each subjected to a repeated measures analysis of variance (RM-ANOVA), with Group (HAM vs. LAM) as between-subject factor, and Trial n-1 congruency (congruent vs. incongruent trials) and Trial n congruency (congruent vs. incongruent) as within-subject factors. Significant interactions were followed-up by simple main- or interaction-effect analyses. Because of the unequal distribution of male and female participants in the two groups, we also performed the RM-ANOVAs with gender as covariate; these analyses yielded the same pattern of results as those described below and will not be reported. Moreover, additional analyses using proportional and log-transformed RTs (to control for possible effects of overall differences in response speed between the two groups) revealed the same pattern of results as those reported below based on the uncorrected RTs. A p value of < 0.05 was adopted as criterion for statistical significance in all analyses; effect sizes were expressed as partial eta-squared. Next to these null hypothesis significance testing (NHST) analyses, we performed Bayesian tests using JASP (Computer software; Version 0.8.1.1) to quantify the strength of the evidence for the alternative compared to the null hypothesis in terms of the Bayes Factor (BF; see Jeffreys, Citation1961), with BF10 > 100 indicating decisive, BF10 between 30 and 100 very strong, BF10 between 10 and 30 strong, BF10 between 3 and 10 substantial, and BF10 between 1 and 3 anecdotal evidence in favour of the alternative hypothesis, respectively. A BF10 between 0.33 and 1 indicates anecdotal, and BF10 between 0.10 and 0.33 substantial evidence in support of the null hypothesis. In these analyses with default prior scales, we contrasted the alternative model consisting of the interaction term or main effect of critical interest against the null model consisting of the (remaining) main, interaction, or no effects. We took the data as evidence for the alternative model if both the NHST revealed a significant effect, and the BF10 suggested at least substantial evidence for the model. Finally, we performed correlation analyses to test the association between achievement motivation and SCEs. For this purpose we expressed SCEs magnitude using the RT data (which gave the clearest results in this and the following experiment; see below) in terms of the difference between percentage congruency effects after congruent and after incongruent trials: [((cI-cC)/cC)-((iI-iC)/iC)]*100 (e.g. see also Ambrosi, Lemaire, & Blaye, Citation2016); the larger the value on this measure, the stronger the SCEs). To further differentiate between the two possible sources of SCEs, we also computed the cI-iI and iC-cC difference scores. For each of these differences scores, a high score reflects a strong adaptation effect. Next, we computed the non-parametric correlation (Spearman's rho) between the score on the AMS and each of the three measures of adaptation.
Results
RTs
The left panel of displays the groups’ mean RT for the different trial pairs. The figure suggests that the HAM participants showed stronger SCEs than the LAM students.
Figure 1. Mean (±SEM) RT (left) and proportion of trials with a correct response (right) for the different trial pairs in Experiment 1 that were defined by the congruency (con = congruent; incon = incongruent) of the current (Trial n) and preceding (Trial n-1) trial, separately for participants with a high achievement motivation (HAM) and a low achievement motivation (LAM).
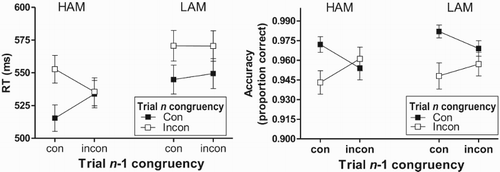
ANOVA revealed a significant Trial n congruency effect, F(1, 64) = 85.83, p < 0.001, , BF10 = 4.87e + 11 (null model including Group, Trial n-1 congruency, and their interaction), and a Trial n-1 congruency × Trial n congruency effect, F(1, 64) = 14.13, p < 0.001,
, BF10 = 796.21 (null model including Group, and Trial n-1 congruency, Trial n congruency and their interaction with Group). Of primary importance, the Group × Trial n-1 congruency × Trial n congruency interaction was also significant, F(1, 64) = 8.27, p = 0.005,
, BF10 = 25.9. A separate Trial n-1 congruency × Trial n congruency ANOVA on the data from the HAM participants revealed a significant Trial n-1 congruency × Trial n congruency interaction, F(1, 35) = 16.26, p < 0.001,
, BF10 = 10489.5, reflecting a weaker congruency effect after a previous incongruent trial (iC vs. iI, p = 0.76, BF10 = 0.3) than after a previous congruent trial (cC vs. cI, F(1, 35) = 59.85, p < 0.001,
, BF10 = 1.3e + 6). This differential congruency effect was due to both cC trials evoking faster responding than iC trials, F(1, 35) = 12.16, p = 0.001,
, BF10 = 22.8, and iI evoking faster responding than cI trials, F(1, 35) = 11.00, p = 0.002,
, BF10 = 14.9. For the LAM participants, the Trial n-1 congruency × Trial n congruency ANOVA only revealed a significant effect for Trial n congruency, F(1, 29) = 39.89, p < 0.001,
, BF10 = 6.7e + 7, reflecting faster responding on current congruent than incongruent trials, regardless of the congruency of the preceding trial, F(1, 29) < 1, p = 0.36,
, BF10 = 0.32, for the Trial n × Trial n-1 interaction. The significant Group × Trial n-1 congruency × Trial n congruency interaction was also examined by analysing the effect of Group for each of the trial-pair types. These analyses revealed (marginally) significantly faster responding for the HAM than LAM participants on cC and iI trial pairs, Fs(1, 64) > 3.89, ps < 0.054,
, BFs10 > 1.2, but not on cI and iC trial pairs, ps > 0.25, BFs10 < 0.4.
Response accuracy
The right panel of shows the mean proportion of trials with a correct response for each of the trial-pair types and groups. ANOVA only revealed a significant main effect of Trial n congruency, F(1, 64) = 13.43, p = 0.001, , BF10 = 215.5 (BF against null model including Group, Trial n-1 congruency, and Group × Trial n-1 congruency effects) reflecting an overall larger accuracy for current congruent compared to incongruent trials, and a significant Trial n-1 congruency × Trial n congruency interaction, F(1, 64) = 18.79, p < 0.001,
, BF10 = 99.6 (BF against null model containing all effects except the target and Group × Trial n-1 congruency × Trial n congruency interaction effects) (other ps > 0.22; F < 1, p = 0.33,
, BF10 = 0.4, for the most critical three-way interaction). The interaction reflected a smaller congruency effect for trials preceded by an incongruent trial (iC vs. iI, p = 0.75, BF10 = 0.2) than for trials preceded by a congruent trial (cC vs. cI, F(1, 65) = 23.88, p < 0.001,
, BF10 = 5466.4). This differential congruency effect was due to both cC trials evoking more accurate responses than iC trials, F(1, 65) = 10.70, p = 0.002,
, BF10 = 18.0 and iI trials evoking more accurate responses than cI trials, F(1, 65) = 6.67, p = 0.01,
, BF10 = 3.4.
Correlation analyses
Spearman's rho was significant for the association between the AMS score and SCE magnitude (ρ = .28, p = .02), and near significant for that between the AMS and cI-iI difference scores (ρ = .24, p = .056). The association between the AMS and iC-cC difference scores was not significant (ρ = .15, p = .24; see Supplementary Material Figure 1 for scatterplots).
Discussion
The present experiment revealed that, when examining RTs in a standard Flanker task, participants with a relatively high achievement motivation showed clear evidence of SCEs, whereas those with a relatively low achievement motivation did not. Both achievement motivation groups showed clear SCEs when considering the performance accuracy data. In terms of correlations, we found an association between the score on the achievement motivation questionnaire and RT-based SCE magnitude, which was primarily based on the association with the cI versus iI trial type performance contrast.
The facts that group differences in the association between achievement motivation and SCEs was significant when examining RTs but not accuracy, are in line with other studies showing that, at least in Flanker tasks, critical differences may emerge when analysing RT but not accuracy data (e.g. Lamers & Roelofs, Citation2011; Schuch et al., Citation2017; van Steenbergen, Band, & Hommel, Citation2010). In the present study, both groups displayed SCEs when analysing the accuracy data for the different trial types. However, the RTs suggest that the LAM participants needed more time to reach this similar pattern of accuracy rates for the different trial pair types than the HAM participants did. This was most strongly the case for the cC and iI trials pairs. These two trial pairs are most critically involved in producing the Gratton effect: widening attention (i.e. including flankers) to facilitate responding on cC trials, and narrowing attention (i.e. blending out distracting flankers) on iI trials.
Whether the group difference in SCEs for the RT data implicates a difference in cognitive control adaptation depends on how to interpret SCEs from interference-control tasks in general. As outlined in the introduction, following the feature-integration or feature-priming account, SCEs in a standard Flanker task may be entirely explained in terms of low-level, bottom-up processes based on differential priming effects associated with full versus partial stimulus repetitions or alternations. Therefore, in Experiment 2 we again assessed the effect of achievement motivation on SCEs in a Flanker tasks, but now while eliminating the potential influence of these low-level processes.
Experiment 2
In Experiment 2, we adopted a version of the Flanker task that controls for repetition confounds inherent to traditional 4-stimuli/2-response Flanker tasks, while at the same time avoiding contingency biases that may be introduced when using an unequal number of congruent and incongruent trials (Duthoo et al., Citation2014). Specifically, we adapted the Flanker task with coloured circle stimuli used by Kim and Cho (Citation2014) to a letter Flanker task similar to that in Experiment 1. The question of interest was whether, using this task, we could replicate the main result of Experiment 1 of differential SCEs as a function of achievement motivation.
Method
Participants
The participants were 143 healthy undergraduate university students from Northwest Normal University. All students first finished the AMS, as in Experiment 1. From this large group, we selected 27% of the participants with the highest and lowest AMS scores as the HAM and LAM groups, respectively. The HAM group (n = 39; 14 men; age: M = 21.21 years, SD = 1.79) had a mean AMS score of M = 7.18 (SD = 5.11; range: 2−26); the LAM group (n = 39; 14 men; age: M = 20.72 years, SD = 1.64) had a mean AMS score of M = −6.92 (SD = 8.03; range: −37−0). The difference in AMS score was highly significant, F(1, 76) = 85.57.85, p < 0.001, . Although the mean AMS score for both the HAM and LAM groups was lower than that for the corresponding groups in Experiment 1, the absolute difference in AMS score between the HAM and LAM groups was similar in both experiments (Experiment 1: 15.12; Experiment 2: 14.10) and a Group × Experiment ANOVA on the AMS score did not reveal a significant interaction effect, F < 1. Moreover, selecting only participants from the present experiment that were matched by a participant from Experiment 1 with an identical AMS score, while applying the same criterion for group membership as in Experiment 1, resulted in the same pattern of results as those reported below. These findings suggest that any differences in results between the two experiments were due to task differences rather than to differences in AMS scores. All participants were right-handed and had a normal or corrected-to-normal visual acuity. The experiment was approved by the Northwest Normal University Ethics Committee.
AMS
The AMS was as described in Experiment 1.
Flanker task
The Flanker task was as described in Experiment 1 except for the following. Next to the four trial types involving the letters H and K (Set 1), we used an additional set of four trial types (Set 2) using the letters D and U (congruent: DDDDD and UUUUU; incongruent: DDUDD and UUDUU). Stimuli from Set 1 were consistently presented on even trials; those from Set 2 on even trials (strict alternating presentation). Each of the 8 possible trial-pair (transition) types that constituted cC trials (i.e. with arrow representing “is followed by”, HHHHH→DDDDD, DDDDD→KKKKK, UUUUU→HHHHH, etc.) was presented an equal number of times within each block of trials. The same was true for cI (i.e. DDDDD→KKHKK, KKKKK→UUDUU, KKKKK→DDUDD, etc.), iC (i.e. DDUDD→KKKKK, HHKHH→DDDDD, KKHKK→UUUUU, etc.), and iI trial pairs (i.e. KKHKK→DDUDD, UUDUU→HHKHH, DDUDD→KKHKK, etc.). The main test was initiated after a practice block of 16 trials (8 congruent and 8 incongruent trials, 8 trials from each of Sets 1 and 2), which was repeated until the participant responded correctly on at least 85% of the trials. The main task consisted of four blocks of 33 trials each. Within each block, the first trial was not used for analysis and the remaining 32 trials were quasi-randomly selected. For each participant we used the same quasi-random order so as to ensure that all trial types (letter combinations) and trial-pair types (cC, iC, cI, and iI) were presented equally often, and the same trial-pair type was not presented more than two times in a row. Most importantly, the strict alternating presentation ensured a complete control of feature repetitions: none of the letters, neither flanker nor target, occurring on trial n-1 was repeated on trial n. Moreover, none of the flanker or target stimuli was predictive of the nature of the target stimulus on the subsequent trial. As in Experiment 1, each trial started with a 500-ms presentation of a fixation cross, which was followed by an empty screen for a variable duration between 300 and 500 ms.
Thereafter, one of eight possible letter strings was shown for a maximum of 1500 ms or until a response was made, whichever came first. Finally, an empty screen was presented for 1000 ms. Participants were instructed to press the “G”, “H”, “J”, and “K” key on a standard keyboard using the fingers of their dominant hand if the middle (target) letter was H, K, D, and U, respectively. Each of the keys was covered by a sticker displaying the corresponding target letter. The task lasted about 15 min.
Procedure and data analysis
The procedure and data analysis were as described for Experiment 1.
Results and discussion
shows the mean RT and response accuracy associated with each of the different types of trial pair for the HAM and LAM groups.
Figure 2. Mean (±SEM) RT (left) and proportion of trials with a correct response (right) for the different trial pairs in Experiment 2 that were defined by the congruency (con = congruent; incon = incongruent) of the current (Trial n) and preceding (Trial n-1) trial, separately for participants with a high achievement motivation (HAM) and a low achievement motivation (LAM).
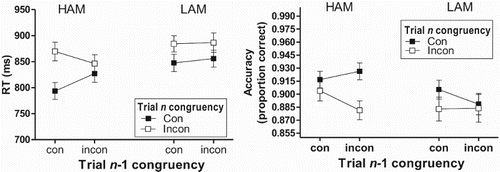
RTs
A Group × Trial n-1 congruency × Trial n congruency ANOVA using the RT data revealed significant effects of Trial n congruency, F(1, 76) = 93.23, p < 0.001, , BF10 = 7.36e + 17, the Trial n-1 congruency × Trial n congruency interaction, F(1, 76) = 40.73, p < 0.001,
, BF10 = 365.75, and the Group × Trial n-1 congruency × Trial n congruency interaction, F(1, 76) = 26.68, p < 0.001,
, BF10 = 54.15 (other ps > 0.10). The latter interaction motivated an ANOVA for each group separately. For the HAM group, this analysis revealed a significant interaction effect, F(1, 38) = 87.73, p < 0.001,
, BF10 = 285782.93 (null model only including main effects), next to a main effect of Trial n congruency, F(1, 38) = 62.44, p < 0.001,
, BF10 = 2.71e + 11. The interaction reflected a smaller congruency effect after an incongruent trial (RT difference between iI and iC: mean = 19.13, SD = 36.94) than after a congruent trial (cI minus cC: mean = 76.18, SD = 46.85). This differential congruency effect reflected faster responding on cC than iC trials, F(1, 38) = 48.73, p < 0.001,
, BF10 = 291406.05, and on iI than cI trials, F(1, 38) = 12.92, p < 0.001,
, BF10 = 30.52. The Trial n-1 congruency × Trial n congruency ANOVA using the data from the LAM group only revealed a main Trial n congruency effect, F(1, 38) = 32.80, p < 0.001,
, BF10 = 8290.44, reflecting faster responding on current congruent than incongruent trials regardless of the congruency of the previous trial, F(1, 38) < 1, p = 0.45, BF10 = 0.26, for the Trial n-1 congruency × Trial n congruency interaction. The significant Group × Trial n-1 congruency × Trial n congruency interaction was also examined by analysing the effect of Group for each of the trial-pair types. These analyses only revealed significantly faster responding for the HAM than LAM participants on cC trial pairs, Fs(1, 76) = 5.42 p = 0.02,
, although Bayesian analysis only revealed anecdotal evidence for this difference, BF10 = 2.36. The group difference was not significant for each of the other trial-pair types, ps > 0.12, BFs10 < 0.70.
Response accuracy
ANOVA using the accuracy data revealed a main effect of Trial n congruency, F(1, 76) = 10.16, p = 0.002, , BF10 = 70.57, which reflected an overall higher accuracy on current congruent (mean = 0.91) than incongruent (mean = 0.89) trials. The Group × Trial n-1 congruency × Trial n congruency interaction was also significant, F(1, 76) = 5.48, p = 0.02,
, but the evidence for a model containing the interaction term against a null model with only the main effects was only anecdotal, BF10 = 1.57. A subsequent Trial n-1 congruency × Trial n congruency ANOVA using the data from the HAM group also only revealed very strong evidence for an effect of Trial n congruency, F(1, 38) = 8.40, p = 0.006,
, BF10 = 86.77, whereas the evidence for a Trial n-1 congruency × Trial n congruency interaction was again anecdotal, F(1, 38) = 6.81, p = 0.01,
, BF10 = 1.46. For the LAM group, the corresponding ANOVA failed to reveal any significant effects, ps > .13, maximum BF10 = .55.
Despite the lack of convincing evidence of differential SCEs in the two groups when considering the accuracy data, we assessed whether the larger SCEs for the HAM than LAM participants when looking at the RT data were significantly affected by a speed-accuracy trade-off. First, we assessed whether there were any signs of such trade off to begin with by computing Spearman's correlations between mean RT and accuracy for each of the trial pair types. If anything, these analyses revealed negative (but non-significant) correlations (higher RTs [slower responses] meaning lower accuracy; ρ = −0.09, p = 0.46 for cC, ρ = 0.01, p = 0.94 for iC, ρ = −0.19, p = 0.09 for cI, and ρ = −0.18, p = 0.11 for iI), suggesting no speed-accuracy trade off. Second, we tested whether the critical Group × Trial n-1 congruency × Trial n congruency interaction for the RT data (reflecting stronger SCEs in the HAM than LAM group) was still significant when controlling for accuracy on the different trial types. A Group × Trial n-1 congruency × Trial n congruency analysis of covariance, with the mean accuracy on cC, cI, iC, and iI trial types as covariates still revealed very strong evidence for a three-way interaction effect, F(1, 72) = 25.93, p < 0.001, , BF10 = 48.21. The interaction again reflected faster responding on cC than iC trials for the HAM, F(1, 34) = 4.94, p = .03,
, BF10 = 344747.61, but not the LAM participants, F < 1, p = 0.38, BF10 = 0.38. Likewise, responding was faster on iI than cI trials for the HAM, F(1, 34) = 8.58, p = .006,
, BF10 = 30.08, but not LAM students, F < 1, p = 0.77, BF10 = 0.26. These results suggest that the interpretation of stronger SCEs for the HAM compared to LAM group derived from the RT analyses was not compromised by the pattern of responding seen for the accuracy data.
Correlation analyses
Spearman's rho was significant for the association between AMS score and RT-based SCE magnitude (ρ = .58, p < .001). The association between the AMS and cI-iI difference score just failed to be significant (ρ = .21, p = .06), whereas the association between the AMS and the iC-cC difference score was significant (ρ = .34, p = .002; see Supplementary Material Figure 1 for the corresponding scatterplots).
In this experiment we used a Flanker task that controlled for lower-level processes that potentially could have affected the SCEs in Experiment 1. As in Experiment 1, for the RT but not accuracy data, we again found reliable evidence for SCEs in the HAM but not LAM participants. Moreover, if anything the correlation analyses revealed an even stronger association between the AMS score and magnitude of (components of) SCEs than was the case in Experiment 1.
General discussion
The present two experiments provide evidence that the extent to which students flexibly adapt cognitive control processes, specifically those implied in interference control, is dependent on their achievement motivation. This evidence was found in Experiment 1 in the form of differential standard SCEs in a standard Flanker task. In Experiment 2, we again used a Flanker task but removed potential confounds by lower processes that are not dependent on cognitive control. We again found evidence for differential moment-to-moment adaptation effects that were dependent on achievement motivation. In both experiments, these adaptations took the form of faster responding to a congruent trial when that trial was preceded by a congruent rather than incongruent trial, and by faster responding to an incongruent trial when that trial had been preceded by an incongruent rather than congruent trial. The positive association between achievement motivation and SCEs was further supported by correlation analyses.
In the introduction we outlined three different accounts of SCEs, the conflict-monitoring, repetition-expectancy, and feature integration accounts. Research suggests that each of the processes described by these accounts may independently contribute to performance in a given conflict task (e.g. Scherbaum, Frisch, Dshemuchadse, Rudolf, & Fischer, Citation2016) but that the extent to which they are involved may be dependent on specific task parameters, such as number of feature repetitions and inter-stimulus interval (ITI) or response-to-stimulus interval (RSI; Duthoo et al., Citation2014). For Experiment 1, we cannot exclude that the standard SCEs found for the HAM group were not partly or even fully due to feature integration or feature-priming effects. However, if such lower-order processes were responsible, it remains to be explained why and how different levels of achievement motivation should affect such processes rather than the adaptation of cognitive control. The task in Experiment 2 removed possible influences of lower-order, bottom-up processes. The achievement-motivation-based differential SCEs found in this experiment suggest a difference in the use of some cognitive control adaptation process, either of the reactive type (conflict-monitoring account) or proactive type (repetition-expectancy account).
One possible explanation of the effect of achievement motivation could be that the present SCEs for the HAM participants were based on the proactive, expectancy-driven process. Accordingly, the HAM participants put more effort into the task than the LAM participants did in terms of more voluntarily and strategically employing expectancy-based control processes. However, in general, expectancy-based adaptations are held to be primarily involved if expectancies concerning the repetition of congruent and incongruent trials are relatively strong, for example by providing explicit cues that inform the participant about the probability of such sequences (Duthoo et al., Citation2014). The experimental design in both experiments implicated equal probabilities of stimulus type repetition and alternation and, therefore, was not conducive to the establishment of strong expectations. Moreover, although the RSI in the present tasks was relatively long (between 1500 and 2000 ms), which might speak to the involvement of the more prolonged expectancy-based processes, this interval is still within the range at which the relatively more short-lived retroactive processes have been shown to be effective (e.g. Egner, Ely, & Grinband, Citation2010).
An alternative mechanism underlying the enhanced cognitive adaptation seen in the HAM individuals may be based on effects that emotions in general have on cognitive control, and adaptations thereof (Chiew & Braver, Citation2011). By definition HAM, but not LAM, individuals anticipate and experience a positive emotion when performing cognitive tasks. Induction of a positive affect has been suggested to improve cognitive flexibility (e.g. Isen, Daubman, & Nowicki, Citation1987), which in turn might be linked to increased dopamine release, affecting brain areas that are critical for cognitive control (e.g. Ashby, Isen, & Turken, Citation1999). Accordingly, because of their more positive affective state, HAM participants display more flexible cognitive control in the form of SCEs than the LAM participants. However, this line of reasoning would be more in line with a proactive rather than reactive account of the present cognitive control adaptations. This is because, if anything, a positive emotion would counteract any negative feelings induced by conflict (incongruent) trials and thereby decreasing the tendency to adapt attentional processes, as assumed to take place in the framework of the conflict-monitoring account (see van Steenbergen et al., Citation2009, Citation2010). Hence, if we dismiss a strong involvement of proactive processes based on the arguments outlined above, we have to look for yet an alternative explanation of the present achievement motivation effects.
A third explanation focusses on motivational rather than (potential) affective effects of a high or low achievement motivation. As outlined in the introduction, there is evidence that resolving a conflict trial may be intrinsically rewarding and can temporarily enhance the participant's motivation (Braem et al., Citation2012; Schouppe et al., Citation2015). If this motivating effect is stronger for HAM than LAM participants, it could be assumed that, after having solved an incongruent trial, the former participants experience a much stronger motivation to repeat the cognitive process that had just been used, be that in the form of a modulation of selective attention or some effort-related process. Note that this line of reasoning assumes that these motivational effects occur very rapidly and automatically, similar as is assumed within the context of the conflict-monitoring account. However, clearly more research is needed to test these assumptions, for example in research that systematically combines intrinsic and extrinsic motivational and affective manipulations.
In principle, SCEs differences based on achievement motivation might result from, or be mediated by, potential differences in executive functions or general IQ rather than from or by differences in achievement motivation (but see Keye, Wilhelm, Oberauer, & Van Ravenzwaaij, Citation2009). For example, a high achievement motivation might be the result of repeatedly having been successful in solving cognitive tasks in general, which in turn is positively associated with cognitive capacities. However, in both studies we used participants that were randomly sampled from one pool of university students and the two groups performed equally well on the tasks in terms of overall response accuracy. Moreover, in an unpublished study we assessed non-verbal fluid IQ using Raven's Advanced Progressive Matrices Test (RAPM; Raven, Court, & Raven, Citation1977) in two selected groups (out of a 175 students, also from Northwest Normal University) with a high (HAM: 17.02) and low (LAM: −.5.5) AMS score. If anything, these two groups had even more extreme AMS scores than the students in the present Experiments 1 and 2 did, but they did not significantly differ in the proportion of correct answers on the RAPM items, M = 0.58 for the HAM and M = 0.57 for the LAM group, F < 1. Moreover, additional correlation analyses assessing the association between the participant's IQ and SCE magnitude indices in this experiment also did not reveal any significant effects. However, future studies should include measures of executive functioning (next to measures of general IQ) to explicitly assess potential differences in cognitive control abilities that might be associated with differences in achievement motivation.
The present results are not in line with those reported in a recent study (Feldman & Freitas, Citation2016). Feldman and Freitas used a Stroop task to measure interference control and conflict adaptations in which they controlled for stimulus repetition and stimulus-contingency confounds. Unlike in our two experiments, in their study they did not find significant associations between a measure of conflict adaptation on one hand and a measure of executive functioning and diverse self-report measures of self-regulation on the other. The self-reports also included the Grit Scale (Duckworth, Peterson, Matthews, & Kelly, Citation2007), which measures the tendency for perseverance for achieving long-term goals, for which they only found a modest and nonsignificant correlation with conflict adaptation (r = .12). However, although “grit” may be related to the AMS, we would like to suggest that the latter scale may be more directly linked to affects associated with performing cognitive tasks. Given the proposed interaction between affective and motivational variables on one hand and SCEs on the other, the AMS might be more sensitive to show associations with SCEs than the Grit Scale. However, clearly more research is needed to explicitly test these assumptions, by including other personality scales in future research.
Conclusion
The present experiments suggest that achievement motivation is one of the factors that may modulate the adaptation of cognitive control that is necessary to deal with current environmental demands. Such modulation may have implications for the interpretation of the results of previous studies and the design of future studies examining motivational and emotional influences on cognitive control adaptations. A further implication of this modulation could be that changing achievement motivation by promoting more positive thoughts regarding one's ability to deal with cognitively demanding tasks in general might also enhance the flexible use of cognitive strategies for the purpose of moment-to-moment adaptations to current cognitive demands.
Supplementary Material Figure 1
Download MS Word (163.7 KB)Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Ambrosi, S., Lemaire, P., & Blaye, A. (2016). Do young children modulate their cognitive control? Sequential congruency effects across three conflict tasks in 5-to-6 year olds. Experimental Psychology, 63(2), 117–126. doi: 10.1027/1618-3169/a000320
- Ashby, F. G., Isen, A. M., & Turken, A. U. (1999). A neuropsychological theory of positive affect and its influence on cognition. Psychological Review, 106(3), 529–550. doi: 10.1037/0033-295X.106.3.529
- Atkinson, J. W. (1964). Introduction to motivation. New York: Van Nostrand.
- Blair, C., & Razza, R. P. (2007). Relating effortful control, executive function, and false belief understanding to emerging math and literacy ability in kindergarten. Child Development, 78(2), 647–663. doi: 10.1111/j.1467-8624.2007.01019.x
- Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108(3), 624–652. doi: 10.1037/0033-295X.108.3.624
- Braem, S., Verguts, T., Roggeman, C., & Notebaert, W. (2012). Reward modulates adaptations to conflict. Cognition, 125(2), 324–332. doi: 10.1016/j.cognition.2012.07.015
- Carlson, S. M., & Moses, L. J. (2001). Individual differences in inhibitory control and children’s theory of mind. Child Development, 72(4), 1032–1053. doi: 10.1111/1467-8624.00333
- Chamorro, J., Bernardi, S., Potenza, M. N., Grant, J. E., Marsh, R., Wang, S., & Blanco, C. (2012). Impulsivity in the general population: A national study. Journal of Psychiatric Research, 46(8), 994–1001. doi: 10.1016/j.jpsychires.2012.04.023
- Chiew, K. S., & Braver, T. S. (2011). Positive affect versus reward: Emotional and motivational influences on cognitive control. Frontiers in Psychology, 2, 279. doi: 10.3389/fpsyg.2011.00279
- Chiew, K. S., & Braver, T. S. (2014). Dissociable influences of reward motivation and positive emotion on cognitive control. Cognitive, Affective, & Behavioral Neuroscience, 14(2), 509–529. doi: 10.3758/s13415-014-0280-0
- Chiew, K. S., & Braver, T. S. (2016). Reward favors the prepared: Incentive and task-informative cues interact to enhance attentional control. Journal of Experimental Psychology: Human Perception and Performance, 42(1), 52–66. doi: 10.1037/xhp0000129
- Diamond, A. (2013). Executive functions. Annual Review of Psychology, 64, 135–168. doi: 10.1146/annurev-psych-113011-143750
- Dreisbach, G., & Fischer, R. (2012a). Conflicts as aversive signals. Brain and Cognition, 78(2), 94–98. doi: 10.1016/j.bandc.2011.12.003
- Dreisbach, G., & Fischer, R. (2012b). The role of affect and reward in the conflict-triggered adjustment of cognitive control. Frontiers in Human Neuroscience, 6, 342. doi: 10.3389/fnhum.2012.00342
- Duckworth, A. L., Peterson, C., Matthews, M. D., & Kelly, D. R. (2007). Grit: Perseverance and passion for long-term goals. Journal of Personality and Social Psychology, 92(6), 1087–1101. doi: 10.1037/0022-3514.92.6.1087
- Duthoo, W., Abrahamse, E. L., Braem, S., Boehler, C. N., & Notebaert, W. (2014). The heterogeneous world of congruency sequence effects: An update. Frontiers in Psychology, 5, 716. doi: 10.3389/fpsyg.2014.01001
- Egner, T. (2007). Congruency sequence effects and cognitive control. Cognitive, Affective, & Behavioral Neuroscience, 7(4), 380–390. doi: 10.3758/CABN.7.4.380
- Egner, T., Ely, S., & Grinband, J. (2010). Going, going, gone: Characterizing the time-course of congruency sequence effects. Frontiers in Psychology, 1, 154. doi: 10.3389/fpsyg.2010.00154
- Egner, T., & Hirsch, J. (2005). The neural correlates and functional integration of cognitive control in a Stroop task. NeuroImage, 24(2), 539–547. doi: 10.1016/j.neuroimage.2004.09.007
- Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16(1), 143–149. doi: 10.3758/BF03203267
- Feldman, J. L., & Freitas, A. L. (2016). An investigation of the reliability and self-regulatory correlates of conflict adaptation. Experimental Psychology, 63(4), 237–247. doi: 10.1027/1618-3169/a000328
- Friedman, N. P., & Miyake, A. (2004). The relations among inhibition and interference control functions: A latent-variable analysis. Journal of Experimental Psychology: General, 133(1), 101–135. doi: 10.1037/0096-3445.133.1.101
- Gjesme, T., & Nygård, R. (1970). Achievement Motivation Scale. Oslo: University of Oslo.
- Gratton, G., Coles, M. G. H., & Donchin, E. (1992). Optimizing the use of information: Strategic control of activation of responses. Journal of Experimental Psychology: General, 121(4), 480–506. doi: 10.1037/0096-3445.121.4.480
- Hagtvet, K. A., & Zuo, L. (2000). Conceptual and empirical components of an internal domain study: An illustration in terms of the achievement motives scale. Scandinavian Journal of Educational Research, 44(1), 49–78. doi: 10.1080/713696664
- Hommel, B. (2004). Event files: Feature binding in and across perception and action. Trends in Cognitive Sciences, 8(11), 494–500. doi: 10.1016/j.tics.2004.08.007
- Hommel, B., Proctor, R. W., & Vu, K.-P. (2004). A feature-integration account of sequential effects in the Simon task. Psychological Research, 68(1), 1–17. doi: 10.1007/s00426-003-0132-y
- Isen, K. A., Daubman, K. A., & Nowicki, G. P. (1987). Positive affect facilitates creative problem solving. Journal of Personality and Social Psychology, 52(6), 1122–1131. doi: 10.1037/0022-3514.52.6.1122
- Jeffreys, H. (1961). Theory of probability (3rd ed.). Oxford, UK: Oxford University Press.
- Keye, D., Wilhelm, O., Oberauer, K., & Van Ravenzwaaij, D. (2009). Individual differences in conflict-monitoring: Testing means and covariance hypothesis about the Simon and the Eriksen Flanker task. Psychological Research Psychologische Forschung, 73(6), 762–776. doi: 10.1007/s00426-008-0188-9
- Kim, S., & Cho, Y. S. (2014). Congruency sequence effect without feature integration and contingency learning. Acta Psychologica, 149, 60–68. doi: 10.1016/j.actpsy.2014.03.004
- Lamers, M. J. M., & Roelofs, A. (2011). Attentional control adjustments in Eriksen and Stroop task performance can be independent of response conflict. Quarterly Journal of Experimental Psychology, 64(6), 1056–1081. doi: 10.1080/17470218.2010.523792
- Man, F., Nygård, R., & Gjesme, T. (1994). The achievement motives scale (AMS): Theoretical basis and results from a first try-out of a Czech form. Scandinavian Journal of Educational Research, 38(3–4), 209–218. doi: 10.1080/0031383940380304
- Mayr, U., Awh, E., & Laurey, P. (2003). Conflict adaptation effects in the absence of executive control. Nature Neuroscience, 6, 450–452. doi: 10.1038/nn1051
- Nigg, J. T. (2000). On inhibition/disinhibition in developmental psychology: Views from cognitive and personality psychology and a working inhibition taxonomy. Psychological Bulletin, 126(2), 220–246. doi: 10.1037/0033-2909.126.2.220
- Nygård, R., & Gjesme, T. (1973). Assessment of achievement motives: Comments and suggestions. Scandinavian Journal of Educational Research, 17(1), 39–46. doi: 10.1080/0031383730170104
- Raven, J. C., Court, J. H., & Raven, J. (1977). Manual for Raven’s progressive matrices and vocabulary scales. London: H. K. Lewis.
- Scherbaum, S., Frisch, S., Dshemuchadse, M., Rudolf, M., & Fischer, R. (2016). The test of both worlds: Identifying feature binding and control processes in congruency sequence tasks by means of action dynamics. Psychological Research. doi: 10.1007/s00426-016-0823-9
- Schouppe, N., Braem, S., De Houwer, J., Silvetti, M., Verguts, T., Ridderinkhof, K. R., & Notebaert, W. (2015). No pain, no gain: The affective valence of congruency conditions changes following a successful response. Cognitive and Affective Behavioral Neuroscience, 15(1), 251–261. doi: 10.3758/s13415-014-0318-3
- Schuch, S., Zweeings, J., Hirsch, P., & Koch, I. (2017). Conflict adaptation in positive and negative mood: Applying a success-failure manipulation. Acta Psychologica, 176, 11–22. doi: 10.1016/j.actpsy.2017.03.005
- Soutschek, A., Stelzel, C., Paschke, L., Walter, H., & Schubert, T. (2015). Dissociable effects of motivation and expectancy on conflict processing: An FMRI study. Journal of Cognitive Neuroscience, 27(2), 409–423. doi: 10.1162/jocn_a_00712
- Stahl, C., Voss, A., Schmitz, F., Nuszbaum, M., Tüscher, O., Lieb, K., & Klauer, K. C. (2014). Behavioral components of impulsivity. Journal of Experimental Psychology: General, 143, 850–886. doi: 10.1037/a0033981
- van Steenbergen, H., Band, G. P. H., & Hommel, B. (2009). Reward counteracts conflict adaptation. Psychological Science, 20(12), 1473–1477. doi: 10.1111/j.1467-9280.2009.02470.x
- van Steenbergen, H. V., Band, G. P. H., & Hommel, B. (2010). In the mood for adaptation: How affect regulates conflict-driven control. Psychological Science, 21(11), 1629–1634. doi: 10.1177/0956797610385951
- Ye, R., & Hagtvet, K. A. (1992). The assessment and analysis of the achievement motives. Psychological Development and Education (China), 2, 14–17.
- Young, S. E., Friedman, N. P., Miyake, A., Willcutt, E. G., Corley, R. P., Haberstick, B. C., & Hewitt, J. K. (2009). Behavioral disinhibition: Liability for externalizing spectrum disorders and its genetic and environmental relation to response inhibition across adolescence. Journal of Abnormal Psychology, 118(1), 117–130. doi: 10.1037/a0014657