
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
The degree to which invasive Acacia species affect South Africa’s livestock production has received little attention. We investigated the ecological impacts of Acacia mearnsii invasion on forage quality and quantity and on soil resources, along A. mearnsii invasion gradients, on South African rangelands and the subsequent conditions following clearing. Grazing capacity was reduced by 72% in densely invaded sites, whereas clearing improved grazing capacity by 66% relative to densely invaded sites within 5 years. In densely invaded sites total grass species basal cover was reduced by up to 42%. As a result, A. mearnsii reduced grazing capacity, from 2 to 8 ha required to support one large stock unit (ha/LSU) in uninvaded and densely invaded sites, respectively. Soil moisture content was lower in densely invaded sites compared with lightly invaded and cleared sites. Plant litter increased from 1.3% to 4.2%, carbon content of the soil increased from 2.0% to 4.0% and nitrogen concentrations increased from 0.1% to 0.2% in response to invasion by A. mearnsii. Clearing resulted in improved grazing capacity within 5 years. These results also showed that, if left uncontrolled, wattle species can reduce livestock carrying capacity within montane grasslands in South Africa.
EDITED BY James Aronson
1. Introduction
Invasion of rangelands by alien plants is a threat to livestock production in South Africa (Wise et al. Citation2012; Shackleton et al. Citation2015; Ndhlovu et al. Citation2016; Stafford et al. Citation2016). Livestock production in South Africa is predominantly rangeland-based, hence rangeland condition is crucial for sustainable forage production (Cousins Citation1999). Fast-growing invasive Acacia species use more water than grasslands and indigenous tree species and alter nutrient flows, potentially reducing the ability of rangeland ecosystems to provide forage (De Neergaard et al. Citation2005; Moyo and Fatunbi Citation2010). Australian Acacia species have the potential to convert grassland into dense woodlands through the displacement of grasses and shrubs (De Neergaard et al. Citation2005). However, little work has been done to quantify the impacts of invasive alien plant invasions on livestock production in South Africa, particularly at local scales (Ndhlovu et al. Citation2016; Stafford et al. Citation2016). This is important because it has clear implications for both land management and human wellbeing in rural areas.
Over the past years, work on understanding the impacts of invasive alien species on South African ecosystems and human wellbeing has been growing and still continues to grow (Richardson and Van Wilgen Citation2004; van Wilgen et al. Citation2008; Le Maitre et al. Citation2011). The Working for Water (WfW) programme is a national alien plant control programme run by the Department of Environment Affairs focussed on controlling the spread of invasive alien species (van Wilgen et al. Citation1998). The primary goal of the programme and the rationale behind its establishment was to secure threatened water resources through the clearing of invasive alien plants countrywide. At the same time the programme also addresses poverty through the creation of employment (van Wilgen et al. Citation1998; Jones et al. Citation2011). The project rests on a further premise that clearing of invasive alien plants will restore ecosystems to their initial conditions (van Wilgen et al. Citation2012).
Acacia mearnsii De Wilde (an introduced species of Fabaceae family from Australia) is one of the most prolific and extensive invaders of rangelands in the more mesic areas of South Africa, particularly in the Eastern Cape, KwaZulu-Natal and Mpumalanga Provinces (Nel et al. Citation2004; De Neergaard et al. Citation2005). Several studies have reported that the removal of Acacia species can facilitate re-colonisation by indigenous species and improve basal vegetation cover (Blanchard and Holmes Citation2008; Ruwanza et al. Citation2013). However, while some areas can recover following the removal of invasive alien plants without further interventions, in other areas active restoration is required to enhance ecosystem recovery (Beater et al. Citation2008; Gaertner et al. Citation2011; Le Maitre et al. Citation2011). In some cases, elevated levels of soil nutrients (Yelenik et al. Citation2007; Gaertner et al. Citation2011; Witkowski Citation2012) derived from nutrient rich litter, and N fixation in the case of legumes, can lead to the undesirable situation of reinvasion by the same and or other species after clearing (Beater et al. Citation2008; Witkowski and Garner Citation2008). Studies of the impacts of invasive alien plants and the recovery of rangelands after clearing can help quantify the damage caused by and benefits of clearing invasive alien plants (Ndhlovu et al. Citation2016). While there are several studies reporting on the benefits of clearing and the recovery of some ecosystem components (Yelenik et al. Citation2004, Citation2007; Blanchard and Holmes Citation2008; Gaertner et al. Citation2011; Witkowski Citation2012) these have not been conducted in the context of grazing capacities.
This study quantifies the impact of invasions by A. mearnsii and benefits associated with clearing operations on range condition and livestock production in the Amathole montane grassland of Stutterheim, South Africa. The objectives of the study were (i) to determine the effects of A. mearnsii density on indigenous grass species cover, and how this translates into impacts in forage quality and quantity; (ii) to determine the effects of A. mearnsii invasion on soil resources and conditions required to support grazing capacity and (iii) to determine the effects of clearing operations on grazing capacity.
2. Materials and methods
2.1. Study site
The study was conducted in the Stutterheim district of the Eastern Cape Province (32° 34ʹ 0.84”S, 27° 25ʹ 0.12”E), South Africa. Stutterheim has a 40-year mean annual rainfall of 752 mm, with most of the rain falling between October and March (O’reagain and Grau Citation1995). Average minimum and maximum temperatures range from 14 to 25°C, in summer, and 7 to 18°C, in winter. Mean annual rainfall of the period of interest (2005–2010 when clearing of invasions was undertaken) has been generally higher than the long term average (calculated for 40 years).The dominant vegetation type found here is Amathole Montane Grassland (Mucina et al. Citation2006) characterised as short grassland with a high species richness of forbs (especially Helichrysum and Senecio species). The dominant grass species include Themeda triandra (Forssk.), Sporobolus africanus (Poir.), Eragrostis chloromelus (Schrad.), E. curvula (Schrad.), Herteropogon contortus (L) and Tristachya leucothrix (Nees), while the karroid shrubs Chrysocoma ciliata (L) and Felicia filifolia (Vent.) are also prevalent. The most prominent invaders into this vegetation type include A. mearnsii (De Wild.) and Acacia dealbata (Link)(Mucina et al. Citation2006) . Geologically, the soils are derived from sedimentary rocks of the Beaufort Group and are characterised by weakly developed lithosols, solonetzic soils interspersed with red clay (Mucina et al. Citation2006). Land use within this region is dominated by livestock production with numerous cattle and sheep stud farms, commercial livestock farms as well as communally based livestock (Kopke Citation1961). Stocking rates range between 2 and 40 ha/LSU under optimal and poor conditions, respectively (Trollope Citation1986). The area also supports forestry industries that include extensive Pinus, Eucalyptus and non-native Acacia species plantations. More than 10% of the landscape has been transformed to plantations and cultivation (Kopke Citation1961).
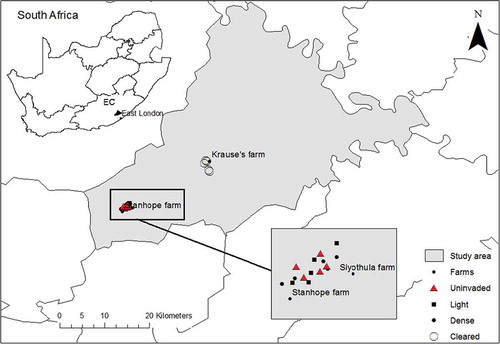
We investigated two levels of wattle tree invasion, lightly invaded and densely invaded, and compared these with uninvaded and cleared sites (). For each of these four treatments five sites were examined (n = 20 sites) on the Stanhope, Siyothula and Krause’s farms () within the Amathole montane grassland, (Mucina et al. Citation2006). Study sites were at least 200 m apart from each other. The initial clearing of A. mearnsii was done 5 years prior to assessment (cleared in 2006). Discussions with landowner confirmed the state of tree density prior to clearing as equivalent to that of densely invaded sites. Clearing operations involved the complete mechanical removal of A. mearnsii by cutting and the application of herbicide on cut stumps to prevent resprouting. Follow-up treatments were applied every year after the initial clearing, and sites were burnt after clearing to stimulate germination from the soil stored seed bank. We acknowledge that we do not have true replicates of cleared sites, as there were no other cleared sites in reasonable proximity, but it does not negate undertaking such work, but it does demand some caution in interpretation of the results. Field surveys were undertaken during the period December 2011 to February 2012.
Figure 1. Map showing location of the study site and the placement of sampling plots.
2.2. Sampling and data collection
We investigated two levels of wattle tree invasion, lightly invaded and densely invaded, and compared these with uninvaded and cleared sites (). Invaded sites (lightly and densely invaded sites) were visually assessed from Google Earth to estimate the aerial cover of wattle trees. For each of these four treatments, five sites were examined (n = 20 sites) on the Stanhope, Siyothula and Krause’s farms () within the Amathole montane grassland (Mucina et al. Citation2006). Study sites were at least 200 m apart from each other. Uninvaded sites were selected to represent the current rangeland condition of the study area. To assess the degradation gradient of the uninvaded sites we compared the rangeland condition score and forage potential (Trollope et al. Citation2006) of the uninvaded sites to that of the benchmark sites (Trollope Citation1986). The rangeland condition score of the uninvaded sites was slightly lower than that of the benchmark, but with similar forage potential. Cleared sites were selected to measure recovery of grassland ecosystem following removal of A. mearnsii. All five replicates of cleared sites were on one farm as there were no other cleared sites in reasonable proximity. We acknowledge that we do not have true replicates of cleared sites, but it does not negate undertaking such work, but it does demand some caution in interpretation of the results. To minimise the effects of pseudo-replication, sample sites were widely spread out on a large area. The limited number and size of the farms available and the need to space out sample plots restricted our sample size. Although small, some of the key variables had low variability and consequently some statistically clear and interesting results were obtained. In these instances, a larger sample size is unlikely to have changed the outcome. However, a large sample may have provided greater power in testing of treatments differences for variables with high variability. Field surveys were undertaken during the period December 2011 to February 2012. The initial clearing of A. mearnsii was done 5 years prior to assessment (cleared in 2006). Discussions with landowner confirmed the state of tree density prior to clearing as equivalent to that of densely invaded sites. Clearing operations involved the complete mechanical removal of A. mearnsii by cutting and the application of herbicide on cut stumps to prevent resprouting. Follow-up treatments were applied every year after the initial clearing, and sites were burnt after clearing to stimulate germination from the soil stored seed bank.
2.2.1. Tree cover
Tree density was determined using the point-centred quarter method (Cottam and Curtis Citation1956), as it is a more suitable method to physically sample the dense wattle stand. The point-centred quarter method collects information on density, frequency, coverage if tree species and only requires a small sampling across the woodland instead of measuring all present trees (Cottam and Curtis Citation1956). One transect of 100 m in the centre of each pot was used to record points at 10 m intervals along each transect. The distance to the nearest tree in each of the four quarters surrounding a recording point was measured and the tree (in case of a multistemmed plant, distance was measured to the stem located at the centre). There were no other tree species present in the sample plots other than A. mearnsii.
Mean distance between wattle tree stems was compared between lightly and densely invaded sites. Subsequently, we defined light invasion as greater than 3 m mean distance and dense invasion as less than 3 m mean distance. Mean distance between wattle trees was used to estimate tree density (plants ha−1) within each density. Densities were calculated by dividing the square of the mean distance (D), expressed in m2, into the area of hectare (10 000 m2/D2) (Cottam and Curtis Citation1956).
2.2.2. Vegetation sampling
At each site, three 100 m parallel transects were laid out 10 m apart. At first, transect was selected in the centre of the site and two other transects were located 10 m on each side. Botanical composition and the basal cover of the herbaceous layer were assessed using the point-to-tuft method described by Trollope (Citation1986). Along each 100 m transect, 50 recordings of species composition were made at 2 m intervals. At each 2 m interval one herbaceous species at or nearest to the point within a 30 cm radius was identified and recorded. If there was no vegetation within the 30 cm radius, the point was recorded as bare ground, to determine basal cover. Grass species were grouped as being Decreaser and Increaser species according to their response to grazing (van Oudtshoorn Citation2006). This ecological grouping system, described below, was then used to determine rangeland condition at each site (van Oudtshoorn Citation2006). Decreaser grass species are species that dominate in rangelands that are optimally utilised, but decrease in relative abundance in rangelands that are over- or underutilised. Increaser species are indicators of the poor range condition and are divided into 3 classes: Increaser I, II and III. Increaser I species are generally unpalatable climax grasses that are abundant in underutilised rangeland and under conditions of little or no herbivory. Increaser II species are grasses that dominate in over utilised rangeland. These include pioneer and sub-climax species that produce much viable seed and can thus quickly establish on new exposed ground. Increaser III species increase in abundance with selective grazing, are generally unpalatable dense climax grasses that are common in overgrazed rangeland. Herbaceous species that did not fall in the above categories (including alien grasses, sedges and forbs) were listed as ‘other’.
2.2.3. Soil sampling
The same plots and 100 m transects that were used for the vegetation assessment were used to survey soils. Ten 1 m2 quadrats were laid out at 10 m intervals along the transects. Visual soil surface assessment was conducted in each quadrat using a method derived in part from the Landscape Functional Analysis method developed by Tongway and Hindley (Citation2004). This method uses several indicators to assess the biogeochemical function of the landscape. The selected indicators for this study include plant litter, deposited material and soil surface roughness. Plant litter accumulation is strongly related to elements stored in the surface soil layers such as carbon and nitrogen. Plant litter was therefore assessed as an indicator for decomposition rates (O’Farrell et al. Citation2009). Deposited material was assessed to determine the amount of alluvium transported to and deposited in the query zone (i.e. the area being examined). Soil surface roughness is related to the ability of the surface to capture mobile resources such as water, propagules, topsoil and organic matter therefore increasing nutrient cycling and water infiltration rate. Indicators of surface roughness such as microtopography (depressions) and high grass plant density were used to assess surface roughness (Tongway and Hindley Citation2004). The amount of dung found in each quadrat was recorded as a proxy for grazing intensity and herbivore pressure. Dung deposits which fell within each quadrat were recorded, and each quadrat was given a score of 1 to 5 based on the amount of dung present. Average dung deposits were calculated for each transect and summed to give that particular plot a score (Hodgins and Rogers Citation1997; Barnes Citation2001). Water infiltration rate was measured in each quadrat to assess the soil’s ability to partition rainfall into soil water and runoff. A single ring infiltrometer was used to assess water infiltration rate by recording the amount of time taken for 25 ml of water to infiltrate the soil surface.
Within each quadrat, a soil sample was taken to a depth of 10 cm. Soil samples for each transect were grouped, providing three bulked samples for a mean value per site. Samples were sent to a commercial laboratory (BemLab, Somerset West), for moisture content and chemical analysis. Chemical dispersion was done using sodium hexametaphosphate (calgon). The soil was air dried, sieved through a 2 mm sieve and analysed for pH (1.0 M KCl), P (Bray II) and total extractable cations, namely potassium (K), calcium (Ca), magnesium (Mg) and sodium (Na) (extracted at pH = 7 with 0.2 M ammonium acetate) and organic Carbon (C) by means of the Walkley-Black method. The extracted solutions were analysed with a Varian ICP-OES optical emission spectrometer (The Non-affiliated Soil Analyses Work Committee Citation1990). Total nitrogen (N) content of soil was determined through total combustion using a Leco Truspec® CN Nitrogen Analyzer. The soil’s Cation Exchange Capacity (CEC) was determined using 0.2 M ammonium acetate method as described by The Non-affiliated Soil Analyses Work Committee (1990), where after the soil was leached with 0.2 M K2SO4. The total NH4+ was then determined using a Varian ICP-OES optical emission spectrometer as indication of CEC.
2.3. Rangeland condition scores and grazing capacity assessment
Rangeland condition refers to the state of the natural vegetation in relation to its ability to withstand soil erosion, support species diversity, provide fuel to support fire and its long-term potential to produce forage for sustained livestock production (Tainton Citation1999). Rangeland condition scores (RCS) were determined by multiplying the percentage cover of each species from each treatment with its forage factor, which is an index of the forage production potential for each species (Trollope Citation1986). Forage factors range from 0 (low potential) to 10 (high potential), (Trollope Citation1986). Rangeland condition scores for species grouped in a given ecological group were summed to give that ecological group’s contribution per treatment and the sums of all the ecological groups were added to give the rangeland condition score for each treatment. Ecological groups whose rangeland condition scores differed significantly between treatments were examined to establish which species were most affected and therefore responsible for changes in grazing capacities following A. mearnsii invasion and clearing. Grazing capacity refers to the potential of a grazeable homogeneous unit of vegetation, expressed as area of land required to maintain a single livestock unit over an extended period of time without deterioration (Tainton Citation1999).
Current grazing capacities (CGC) (potential of the vegetation in its current state to provide forage for livestock) for each treatment were determined using the estimates of rangeland condition scores (RCS) obtained for each treatment, as presented by Danckwerts (Citation1989). Rangeland condition scores obtained from different treatments were each divided by the condition score of the benchmark (CSB), as determined by Trollope (Citation1986). To drive current grazing capacity for each treatment in hectares per large stock unit (ha/LSU) the proportions obtained were multiplied by the potential grazing capacity of the benchmark (PGC) (Trollope Citation1986) to derive current grazing capacity for each treatment in hectares per large stock unit (ha/LSU). The formula was:
(Danckwerts Citation1989).
2.4. Statistical analysis
This study tested the impacts of A. mearnsii invasion on grazing capacity of montane grasslands of the Eastern Cape. Four treatments (uninvaded, lightly invaded, densely invaded and cleared) were applied to compare the impacts of A. mearnsii invasion on the mean grazing capacities of montane grasslands. Mean grazing capacities were compared between treatments (uninvaded, cleared, lightly invaded and densely invaded). Impacts of A. mearnsii invasion and clearing on soil physical properties (infiltration rate and moisture content), soil surface properties (litter cover, deposited material, surface roughness and dung density) and chemical properties (pH, P, N, C, Na, K+, Ca+, Mg+ and CEC) were evaluated by comparing mean concentrations in different treatments. Significant differences in the above variables between the treatments were taken to represent the impacts of invasion and clearing of A. mearnsii. Standard errors were used to express variation around means. The effects of different treatments on means were compared using a one way ANOVA in STATISTICA 12 (StatSoft Inc Citation2012). Data were first tested for normality using Kruskal-Wallis. Effects of different treatments on means were compared using Welch ANOVAs followed by Games-Howell post-Hoc test, as the data was not normally distributed. Differences between means were considered significant at p ≤ 0.05.
3. Results
3.1. Vegetation assessment
3.1.1. Tree density
Mean distance between tree stems on lightly and densely invaded sites was 4.0 and 2.7 m, respectively (Cottam and Curtis Citation1956). Densely invaded sites had higher tree density (1 371.74 plants ha-1). Density was much lower in lightly invaded sites (625 plants ha-1).
3.1.2. Impacts of A. mearnsii invasion and clearing on individual species basal cover
The mean distance between wattle stems on densely invaded sites was significantly shorter (F = 4.467, p < 0.001) than on lightly invaded sites. Invasion by A. mearnsii reduced species basal cover by 15 and 42% on lightly and densely invaded sites, respectively, when compared to uninvaded sites. There was no significant difference between the mean basal cover on uninvaded and lightly invaded sites. Mean herbaceous cover was improved by clearing of A. mearnsii and both cleared and uninvaded sites had higher (F = 6.62, p < 0.000) basal cover than densely invaded sites. The uninvaded and cleared sites had no bare areas recorded during the study. Bare area constituted 15% of the lightly invaded site area and nearly 60% of the area of the densely invaded sites ().
Figure 2. Mean percent basal cover ± SE for ecological groups in uninvaded (n = 5), lightly invaded, (n = 5), dense invaded (n = 5) and cleared (n = 5) sites in the Amathole Montane Grassland of Stutterheim, Eastern Cape (F = 6.62, p < 0.000) .
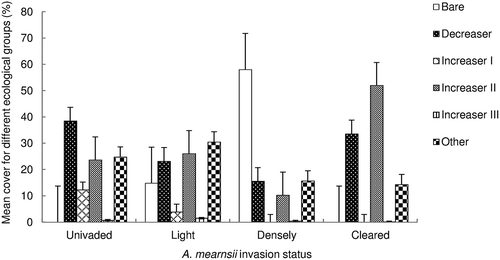
Mean percent basal cover for Decreaser species, Increaser II species, Increaser III species and species falling into the ‘other’ group () differed significantly between treatments. Mean percent basal cover of Decreaser species on uninvaded sites was higher than that on densely invaded sites. Increaser II species mean percent basal cover on cleared sites was higher than that on densely invaded sites. Increaser III species mean percent basal cover was higher on cleared sites when compared to all other categories. Cover of ‘other’ was higher on lightly invaded sites when compared to cleared sites ().
3.2. Soil assessment
3.2.1. Water infiltration and moisture content
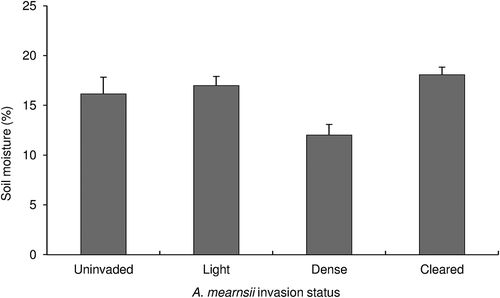
Water infiltration rate was significantly faster on lightly and densely invaded sites when compared to both uninvaded and cleared sites (). There was no significant difference between lightly and densely invaded sites. Water infiltration rate was faster on uninvaded sites when compared to cleared sites (). There was no significant difference in soil moisture content between uninvaded and lightly invaded sites and between uninvaded and densely invaded sites (). However, percentage soil moisture was significantly higher on lightly invaded sites than densely invaded sites. Soil moisture content was higher on the cleared sites than on densely invaded sites ().
Table 1. Means ± SE values of infiltration rate and layer properties in uninvaded (n = 5), lightly invaded (n = 5), dense invaded (n = 5) sites and cleared (n = 5) in the Amathole Montane Grassland of Stutterheim, Eastern Cape.
Figure 3. Average percent moisture content ± SE for difference sites: for uninvaded (n = 5), lightly invaded, (n = 5), densely invaded (n = 5) and cleared (n = 5) sites in the Amathole Montane Grassland of Stutterheim, Eastern Cape. All differences are significant at p < 0.05, Games Howell Post-Hoc test (F = 5.40, p < 0.01) .
3.2.2. Impacts of A. mearnsii invasion and clearing on soil surface properties
Plant litter cover increased with increasing density of A. mearnsii (). Litter cover was higher on densely invaded sites when compared to lightly invaded, uninvaded sites and cleared sites. Lightly invaded sites had higher litter cover than uninvaded and cleared sites. There was no significant difference between cleared and uninvaded sites (). There was a significant difference between densely invaded sites when compared to uninvaded and cleared sites (). There was no evidence of deposited material in uninvaded and cleared sites. Deposited material was more abundant on densely invaded sites than in lightly invaded sites. Soil surface roughness was less in dense invaded sites than on uninvaded, lightly invaded and cleared sites. Dung density was lowest in densely invaded sites when compared to uninvaded, lightly invaded and cleared sites ().
3.2.3. Impacts of A. mearnsii invasion and clearing on soil chemical properties
Sites that were densely invaded by A. mearnsii were slightly more acidic compared to the uninvaded and cleared sites. There was no significant difference on pH levels between densely and lightly invaded sites and between cleared and uninvaded sites (). Cleared sites had higher pH than densely invaded sites. Densely invaded sites had higher concentrations of P, C and N than uninvaded sites (), whereas, Na and Mg showed a relative decrease in densely invaded sites compared to uninvaded sites. There were no significant differences on pH, P, C, K, Ca and CEC concentrations between lightly invaded and uninvaded sites, lightly and densely invaded and lightly and cleared sites (). However, differences were noted for N which increases significantly during light invasion, as well as for Na which decreases significantly when compared to uninvaded sites. Cleared sites had higher P concentrations than uninvaded sites and higher K concentrations when compared to densely invaded, lightly invaded and uninvaded sites, whereas Na concentrations were significantly lower under cleared sites when compared to uninvaded sites. Lower concentrations of Mg were noted under cleared sites when compared to uninvaded and lightly invaded sites ().
Table 2. Mean percent content ± SE for soil chemical properties in uninvaded (n = 5), cleared (n = 5), lightly invaded (n = 5) and dense invaded (n = 5) sites in the Amathola Montane Grassland of the Eastern Cape.
3.3. Grazing capacity assessment and species condition scores underlying changes in grazing capacity
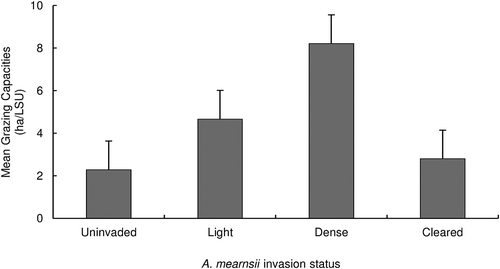
Invasion by A. mearnsii reduced grazing capacity by 56% and 72% on light and densely invaded sites, respectively, whereas grazing capacity on cleared sites had increased to levels resembling uninvaded sites ().
Figure 4. Mean grazing capacities ± SE for uninvaded (n = 5), lightly invaded, (n = 5), densely invaded (n = 5) and cleared (n = 5) sites in the Amathole Montane Grassland. All differences are significant at p < 0.05, Games Howell Post-Hoc test (F = 14.84, p < 0.05).
Means for all grass ecological groups except for species falling into the ‘other’ group (F = 4.32, p < 0.05) differed significantly in rangeland condition scores between treatments. Of the four Decreaser grass species present in the study sites (), only Themeda triandra had a significant contribution to the higher rangeland condition score in uninvaded sites when compared to lightly and densely invaded sites. Increaser II species contributed significantly (F = 4.32, p < 0.05) to the mean grazing capacity of the cleared sites, with Cynodon dactylon showing significant contributions.
Table 3. Mean range condition scores ± SE for different species in uninvaded (n = 5), lightly invaded, (n = 5), densely invaded (n = 5) and cleared (n = 5) sites in the Amathole Montane Grassland of Stutterheim, Eastern Cape.
4. Discussion
4.1. Effects of A. mearnsii invasion on rangeland condition and grazing capacity
The results demonstrate a severe decline in grazing capacity as a result of A. mearnsii invasion. The reduction in grazing capacity, from 2 to 8 ha required to support one large livestock unit on uninvaded and densely invaded sites respectively has important socio-economic and human wellbeing implications (Stafford et al. Citation2016). Acacia species have been reported to affect conditions that are essential for grazing provision, including loss of species diversity, herbaceous cover, altered soil conditions (Ndou and Ruwanza Citation2016). In this study the loss of grazing capacity in densely invaded sites is highly attributed to displacement of total basal cover. The impact on lightly invaded sites include loss of grass cover and that of high gazing value Decreaser species Themeda triandra and Sporobolus fimbriatus, which were more prominent in uninvaded sites. The reduction in herbaceous and grass cover resulting from invasion by Acacia has also been reported by several studies (Holmes Citation2005; Blanchard and Holmes Citation2008; Morris et al. Citation2008; Ruwanza et al. Citation2013). Acacia species have qualities, both above and below ground, which give them a competitive advantage over native species for growth resources. The high leaf biomass results in shading of below canopy species from essential growth resources such as light, reducing the amount of precipitation that reaches the ground (Everson et al. Citation2014). As a consequence, Acacia biomass can alter habitat suitability for indigenous vegetation by reducing light penetrability which changes the microclimate below the canopy tree canopy (Holmes and Cowling Citation1997; Morris et al. Citation2011). Furthermore, high leaf litter content deposited under tree canopy, as evident in the densely and lightly invaded sites in this study, may change the nutrient status of the soil to be more suitable for the invading species. The high root biomass of A. mearnsii is associated with high moisture and nutrient uptake (Dye and Jarmain Citation2004; Yelenik et al. Citation2007; Morris et al. Citation2011).
In this study, dense invasion reduced soil moisture by 4.97%. Because of the reported comparatively excessive water use by Acacia species (Dye and Jarmain Citation2004; Gaertner et al. Citation2011; Le Maitre et al. Citation2016), we anticipated that all invaded sites would have significantly less soil moisture. However, local conditions and factors such as rainfall during the survey period played a more influencing role. The findings of studies such like the one by Yelenik et al. (Citation2004) confirm this, finding higher soil moisture content under dense Acacia stands compared with their fynbos reference sites. Above average rainfall during the survey period for this study may have resulted in higher soil moisture in uninvaded and cleared sites. Water infiltration rate was higher in lightly and densely invaded sites than in uninvaded and cleared sites. High water infiltration rates on invaded sites could be attributed to degraded soil surface conditions and poor basal cover; this is in line with the findings by O’Farrell et al. (Citation2009). This could imply that while infiltration rate is faster on invaded sites, the soil may also dry out faster. Whereas in uninvaded sites, good cover of standing biomass slows water runoff and allows time for water to infiltrate into the soil. Similarly, Mills and Fey (Citation2004) in their study, associated water infiltration with high crust brokenness in invaded sites. Furthermore, trampling on bare soil by grazing animals can affect soil structure, and therefore water infiltration rates, and surface water runoff (Pietola et al. Citation2005; Eldridge and Whitford Citation2009; Dunne et al. Citation2011), particularly in lightly invaded sites as livestock tend to congregate here during extreme weather conditions. As anticipated, we found higher litter content in invaded sites when compared to uninvaded and cleared, with densely invaded sites having almost double the amount of plant litter on lightly invaded sites. This can be explained by the high leaf biomass produced by A. mearnsii as found by other studies (Yelenik et al. Citation2007; Gaertner et al. Citation2011; Witkowski Citation2012). High soil surface roughness on uninvaded sites was explained by presence of standing grass tussocks. The lower dung content found under densely invaded sites is an indication of low grazing animal activity (Hodgins and Rogers Citation1997; Barnes Citation2001). Indicating that the dense wattle stands can form a barrier to animal movement (De Neergaard et al. Citation2005). Low grass cover at these sites further supports the notion that these environments are avoided by grazing animals. The presence of some palatable grass species in the lightly invaded sites and higher dung content of these sites suggests that these sites are being utilised to a higher degree by animals than densely invaded sites. Acacia trees at this density do not create a barrier to animal movement, unlike densely invaded sites (De Neergaard et al. Citation2005), and may provide shade and shelter during extreme weather conditions.
The consequences of A. mearnsii invasion on soil nutrient status were more evident in the densely invaded sites than lightly invaded sites. Findings of this study concur with those of Montgomery (Citation2001), where densely invaded sites had more acidic soils than uninvaded sites. Previous studies (Yelenik et al. Citation2004; Witkowski Citation2012) have found that A. mearnsii invaded sites had higher concentrations of P, C and N. In contrast (Oelofse et al. Citation2016), C content (at 60 cm) in the grassland of Matatiele in Motseng area was reported to be higher when compared to wattle invaded sites. In the same study, Oelofse et al. (Citation2016) found no differences in C content between grassland and wattle invaded sites in the Madlangala study sites, in the same grassland type. They concluded that the lack of differences between grassland and wattle stand in Madlangala could be due to management effects and burning. Yelenik et al. (Citation2007) found no differences in K and Na concentrations under A. saligna canopy but reported higher levels of Mg and Ca when compared to the fynbos species dominated soils. Results of this study showed low levels of Mg and Na concentrations under A. mearnsii canopy, whereas there were no differences in Ca and K concentrations under all invaded treatments. In this study, the higher the concentration of P, C and N on invaded sites could be attributed to a combination of factors, including addition of these elements from nutrient rich plant litter biomass from wattle trees. This may also be an effect of the relatively short time period since clearing. Witkowski (Citation1991) and Yelenik et al. (Citation2007) associated higher levels of nutrients with the redistribution of soil nutrients by Acacia trees from deeper layers of the soil. Acacia species have been associated with high nutrient uptake when compared to native vegetation, and this might be the cause of low concentrations of some nutrient levels under A. mearnsii canopy in this study. Increased soil N concentrations under Acacia canopy have been reported by several studies and have been associated with high levels of nitrogen-rich plant litter. A. mearnsii is a leguminous tree with N2 fixing capability (Yelenik et al. Citation2004) and has the capacity to increase soil N concentrations.
4.2. Changes in rangeland condition and grazing capacity following the clearing of A. mearnsii
The results showed a considerable increase in rangeland condition and gains in grazing capacity 5 years after the clearing of A. mearnsii. These improvements could be attributed to a general increased cover of the grass layer since the removal of A. mearnsii. The increased abundance of Decreaser and Increaser II species on cleared sites indicating an increase in palatability resulted in high grazing index values (Trollope Citation1986; Van Oudtshoorn Citation2006). The improvement of herbaceous vegetation cover after Acacia clearing has been reported by several studies (Blanchard and Holmes Citation2008; Morris et al. Citation2008; Ruwanza et al. Citation2013). Clearing of A. mearnsii would have released the pressure on soil nutrients and other essential resources such moisture and light and enhanced availability of these growth resources to allow plant successional pathways to take place (Yelenik et al. Citation2004). Morris et al. (Citation2008) and Ndhlovu et al. (Citation2011) reported an increase in species richness, cover and abundance of native vegetation after complete removal of alien invasive species. They found that proper follow-up treatments after clearing contributed significantly to the recovery of indigenous vegetation.
High rainfall during our field survey period may have masked any longer-term effects of invasion on soil moisture. However, our results show that soils in cleared sites could hold soil moisture much longer than densely invaded sites. As reported by Everson et al. (Citation2014) dense wattle invasion could have intercepted much of the rainfall resulting in low soil moisture in densely invaded sites. Furthermore, high moisture content and therefore slow water infiltration rates on cleared sites could be attributed to good herbaceous cover. Clearing of trees reduces water loss from the soil by wattle trees. The absence of the wattle canopy also allows more precipitation to reach the soil surface and therefore, improves soil water availability to the grass species (Everson et al. Citation2014).
Plant litter was significantly lower on cleared sites than on invaded sites, and this could be explained by the absence of litter-producing Acacia species (Yelenik et al. Citation2004; Witkowski Citation2012). Furthermore, the absence of deposited material in both uninvaded and cleared sites could be attributed to good standing grass tussock and the absence of A. mearnsii, respectively (Mills and Fey Citation2004).
The legacy effects of A. mearnsii appear to have raised the soil fertility of cleared sites. Concentrations of soil nutrients on cleared sites were similar to that of densely invaded sites as has been reported by previous studies (Yelenik et al. Citation2004, Citation2007; Gaertner et al. Citation2011). In this study, the short period since clearing is likely to have contributed to the observed similarities between densely invaded and cleared sites. Furthermore, perhaps soil samples taken at a more shallow depth (e.g. 1–3 cm) would likely have provided more information on the soil nutrients. Soil pH and K concentrations on cleared sites were higher than on densely invaded sites and similar to that of uninvaded sites (Montgomery 2001). Soil phosphorus, organic carbon and nitrogen content were the same on invaded and cleared treatments. Corbin and D’Antonio (Citation2004) suggested that Acacia spp. have an extended legacy after clearing and this result in a lag effect with the slow release of available N from the higher N pool in the soil. Furthermore, high grass cover on cleared sites is likely to have contributed to increased soil nutrients on cleared sites by reducing water run-off, thus the loss of organic matter and nutrients.
Our study clearly demonstrates that significant grazing resources are lost under conditions of A. mearnsii invasion. Clearing of A. mearnsii can help to restore the decline in grazing capacity, but this is conditional on the autogenic recovery of the grass layer. Increased soil nutrient levels, which can remain for an extended period after clearing of A. mearnsii, could have the impact of improving grazing condition. However, considering that all cleared sites were located on the same farm, there is no way of separating the role of management effects on the successful recovery of cleared sites. We speculate that the successful recovery following alien removal could be due to well-timed-follow up treatments. Wattle seed bank sizes, disturbances during clearing, and the potential abundance of nutrients increase the potential for re-establishment of the same species, therefore careful monitoring of cleared sites is essential to ensure ecosystem recovery. The costs involved in clearing alien invasive plants is very high (Marais et al. Citation2004) and it is unrealistic to assume that poorly resourced, emerging farmers will be able to undertake such interventions. Therefore, more research is needed to understand where in the landscape restoration is mandatory and where follow-up alone is needed.
The overall implications here are that the negative impacts of invasive alien plants on grazing resource as an ecosystem services that supports livestock production and associated livelihoods are substantial and warrant the same focus that impacts of invasive alien plants on water resources have received. Furthermore, if alien plant invasions continue unabated, South Africa’s livestock production and therefore agricultural economy will most certainly be threatened by invasive alien plants. The cost of wattle clearing can be prohibitive so government interventions that assist emerging farmers in this process of clearing invasive alien plants will both enhance the speed of recovery and the chances of creating sustainable rural economies.
Acknowledgements
We thank Brian van Wilgen (SU), David Le Maitre (CSIR), Charlie Shackleton (RU) and three anonymous reviewers for their thoughtful comments, Ryan Blanchard (CSIR) for assisting with the analysis and Ilse Kotzee (CSIR) for the study site map. We also thank Dohne Agricultural Institute, Department of Agriculture staff from Stutterheim for assisting with site identification and to Asanda Yaphi for his assistance with data collection. Special thanks to the farmers and staff for allowing access to the sites. This study was funded by Department of Environmental Affairs’ Working for Water Programme and CSIR.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Barnes RFW. 2001. How reliable are dung counts for estimating elephant numbers? Afr J Ecol. 39:1–9.
- Beater MMT, Garner RD, Witkowski ETF. 2008. Impacts of clearing invasive alien plants from 1995 to 2005 on vegetation structure, invasion intensity and ground cover in a temperate to subtropical riparian ecosystem. South African J Bot. 74:495–507.
- Blanchard R, Holmes PM. 2008. Riparian vegetation recovery after invasive alien tree clearance in the Fynbos Biome. South African J Bot. 74:421–431.
- Corbin JD, D’Antonio CM. 2004. Can carbon addition increase competitiveness of native grasses? A case study from California. Restor Ecol. 12:36–43.
- Cottam G, Curtis JT. 1956. The use of distance measures in phytosociological sampling. Ecology. 37:451–460.
- Cousins B. 1999. Invisible capital : the contribution of communal rangelands to rural livelihoods in South Africa. Dev South Afr. 16:299–318.
- Danckwerts J. 1989. Monitoring of vegetation and assessment of veld condition in grassveld. In: Danckwerts J, Teague W, editors. Veld Manag East Cape. Eastern Cape: Department of Agriculture and Water Supply.
- De Neergaard A, Saarnak C, Hill T, Khanyile M, Berzosa AM, Birch-Thomsen T. 2005. Australian wattle species in the Drakensberg region of South Africa - an invasive alien or a natural resource? Agric Syst. 85:216–233.
- Dunne T, Western D, Dietrich WE. 2011. Effects of cattle trampling on vegetation, infiltration, and erosion in a tropical rangeland. J Arid Environ. 75:58–69.
- Dye P, Jarmain C. 2004. Water use by black wattle (Acacia mearnsii): implications for the link between removal of invading trees and catchment streamflow response. S Afr J Sci. 100:40–44.
- Eldridge DJ, Whitford WG. 2009. Soil disturbance by native animals along grazing gradients in an arid grassland. J Arid Environ. 73:1144–1148.
- Everson CS, Clulow A, Becker M, Watson A, Ngubo C, Bulcok H, Mengistu MG, Lorentz S, Demlie M. 2014. The long term impact of Acacia mearnsii trees on evaporation, streamflow, low flows and ground water resources. Phase II: understanding the controlling environmental variables and soil water processes over a full crop rotation. Pretoria: South African Water Research Commission.
- Gaertner M, Richardson DM, Privett SDJ. 2011. Effects of alien plants on ecosystem structure and functioning and implications for restoration: insights from three degraded sites in South African fynbos. Environ Manage. 48:57–69.
- Hodgins IW, Rogers RW. 1997. Correlations of stocking with the cryptogamic soil crust of a semi-arid rangeland in southwest Queensland. Austral Ecol. 22:425–431.
- Holmes PM. 2005. A decision-making framework for restoring riparian zones degraded by invasive alien plants in South Africa. South African J Sci. 101:553–564.
- Holmes PM, Cowling RM. 1997. The effects of invasion by acacia saligna on the guild structure and regeneration capabilities of South African Fynbos Shrublands. J Appl Ecol. 34:317–332.
- Jones C, Pech R, Forrester G, King CM, Murphy EC. 2011. Functional responses of an invasive top predator Mustela erminea to invasive meso-predators Rattus rattus and Mus musculus, in New Zealand forests. Wildl Res. 38:131–140.
- Kopke D 1961. Land use in ward one of the Stutterheim district. Rhodes University. http://contentpro.seals.ac.za/iii/cpro/DigidalItemViewPage
- Le Maitre DC, Forsyth GG, Dzikiti S, Gush MB. 2016. Estimates of the impacts of invasive alien plants on water flows in South Africa. Water. SA.42:659–672.
- Le Maitre DC, Gaertner M, Marchante E, Ens EJ, Holmes PM, Pauchard A, O’Farrell PJ, Rogers AM, Blanchard R, Blignaut J, et al. 2011. Impacts of invasive Australian acacias: implications for management and restoration. Divers Distrib. 17:1015–1029.
- Marais C, Van Wilgen BW, Stevens D. 2004. The clearing of invasive alien plants in south africa: a preliminary assessment of costs and progress. S Afr J Sc. 100:97–103.
- Mills AJ, Fey MV. 2004. Effects of vegetation cover on the tendency of soil to crust in South Africa. Soil Use Manag. 20:308–317.
- Montgomery C. 2001 July. Understanding acidic soil. Farmer's Weekly, Caxton Magazines, Johannesburg, pp. 32.
- Morris TL, Esler KJ, Barger NN, Jacobs SM, Cramer MD. 2011. Ecophysiological traits associated with the competitive ability of invasive Australian acacias. Divers Distrib. 17:898–910.
- Morris TL, Witkowski ETF, Coetzee JA. 2008. Initial response of riparian plant community structure to clearing of invasive alien plants in Kruger National Park, South Africa. South African J Bot. 74:485–494.
- Moyo HPM, Fatunbi AO. 2010. Utilitarian perspective of the invasion of Some South African Biomes by Acacia mearnsii. Glob J Environ Res. 4:6–17.
- Mucina L, Hoare DB, Lötter MC, du Preez PJ, Rutherford MC, Scott-Shaw CR, Bredenkamp GJ, Powrie LW, Scott L, Camp KGT, et al.. 2006. Grassland biome. In: Mucina L, Rutherford R, editors. Veg South Africa, Lesotho Swazil. Pretoria: South African National Biodiversity Institute; p. 338–436.
- Ndhlovu T, Milton SJ, Esler KJ. 2016. Effect of Prosopis (mesquite) invasion and clearing on vegetation cover in semi-arid Nama Karoo rangeland, South Africa. African J Range Forage Sci. 33:11–19.
- Ndhlovu T, Milton-Dean SJ, Esler KJ. 2011. Impact of prosopis (mesquite) invasion and clearing on the grazing capacity of semiarid nama karoo rangeland, south africa. Afr J Range Forage Sci. 28:129–137.
- Ndou E, Ruwanza S. 2016. Soil and vegetation recovery following alien tree clearing in the Eastern Cape Province of South Africa. Afr J Ecol. 54:460–470.
- Nel JL, Richardson DM, Rouget M, Mgidi TN, Mdzeke N, Le Maitre DC, van Wilgen BW, Schonegevel L, Henderson L, Neser S. 2004. A proposed classification of invasive alien plant species in South Africa: towards prioritizing species and areas for management action. S Afr J Sci. 100:53–64.
- O’Farrell PJ, Donaldson JS, Hoffman MT. 2009. Local benefits of retaining natural vegetation for soil retention and hydrological services. South African J Bot. 75:573–583.
- Oelofse M, Birch-Thomsen T, Magid J, de Neergaard A, van Deventer R, Bruun S, Hill T. 2016. The impact of black wattle encroachment of indigenous grasslands on soil carbon, Eastern Cape, South Africa. Biol Invasions. 18:445–456.
- O'Reagain PJ, Grau EA. 1995. Sequence of species selection by cattle and sheep on south african sourveld. J Range Manag. 48:314–321.
- Pietola L, Horn R, Yli-Halla M. 2005. Effects of trampling by cattle on the hydraulic and mechanical properties of soil. Soil Tillage Res. 82:99–108.
- Richardson DM, Van Wilgen BW. 2004. Invasive alien plants in South Africa: how well do we understand the ecological impacts? S Afr J Sci. 100:45–52.
- Ruwanza S, Gaertner M, Esler KJ, Richardson DM. 2013. Both complete clearing and thinning of invasive trees lead to short-term recovery of native riparian vegetation in the Western Cape, South Africa. Appl Veg Sci. 16:193–204.
- Shackleton RT, Le Maitre DC, Van Wilgen BW, Richardson DM. 2015. The impact of invasive alien Prosopis species (mesquite) on native plants in different environments in South Africa. South African J Bot. 97:25–31.
- StatSoft. 2012. Statistica: data analysis software system. Tulsa (OK): StatSoft Inc.
- Stafford W, Birch C, Etter H, Blanchard R, Mudavanhu S, Angelstam P, Blignaut J, Ferreira L, Marais C. 2016. The economics of landscape restoration: benefits of controlling bush encroachment and invasive plant species in South Africa and Namibia. Ecosyst Serv. 27:193–202.
- Tainton N. 1999. Veld management in South Africa. Pietermaritzburg: University of Natal Press.
- The Non-Affiliated Soil Analysis Work Committee. 1990. Handbook of Standard Soil Testing Methods for Advisory Purposes. Pretoria: Soil Science Society of South Africa.
- Tongway D, Hindley N. 2004. Landscape function analysis: a system for monitoring rangeland function. African J Range Forage Sci. 21:109–113.
- Trollope W, Trollope L, de Bruno Austin C, Held A, Emery A, Hines CJH. 2006. A fire management plan for the Okavango Delta RAMSAR site in Botswana. Gabarone: Botswana Department of Forestry and Range Resources; p. 61–79.
- Trollope WSW. 1986. Land use surveys: assessment of veld condition in the Ciskei. In: Republic of Ciskei national soil conservation strategy. Bisho: South African Department of Agriculture and Forestry. p. 1–37.
- van Oudtshoorn F. 2006. Guide to grasses of Southern Africa. 2nd ed. Pretoria: Briza Pubications.
- van Wilgen BW, Forsyth GG, Le Maitre DC, Wannenburgh A, Kotzé JDF, van den Berg E, Henderson L. 2012. An assessment of the effectiveness of a large, national-scale invasive alien plant control strategy in South Africa. Biol Conserv. 148:28–38.
- Van Wilgen BW, Le Maitre DC, Cowling RM. 1998. Ecosystem services, efficiency, sustainability and equity: South Africa’s working for water programme. Trends Ecol Evol. 13:378.
- van Wilgen BW, Reyers B, Le Maitre DC, Richardson DM, Schonegevel L. 2008. A biome-scale assessment of the impact of invasive alien plants on ecosystem services in South Africa. J Environ Manage. 89:336–349.
- Wise RM, van Wilgen BW, Le Maitre DC. 2012. Costs, benefits and management options for an invasive alien tree species: the case of mesquite in the Northern Cape, South Africa. J Arid Environ. 84:80–90.
- Witkowski AETF. 2012. Effects of invasive alien acacias on nutrient cycling in the coastal lowlands of the cape fynbos. Society. 28:1–15.
- Witkowski ETF. 1991. Effects of invasive alien acacias on nutrient cycling in the coastal lowlands of the cape fynbos. J Appl Ecol. 28:1.
- Witkowski ETF, Garner RD. 2008. Seed production, seed bank dynamics, resprouting and long-term response to clearing of the alien invasive Solanum mauritianum in a temperate to subtropical riparian ecosystem. South African J Bot. 74:476–484.
- Yelenik SG, Stock WD, Richardson DM. 2004. Ecosystem level impacts of invasive Acacia saligna in the South African fynbos. Restor Ecol. 12:44–51.
- Yelenik SG, Stock WD, Richardson DM. 2007. Functional group identity does not predict invader impacts: differential effects of nitrogen-fixing exotic plants on ecosystem function. Biol Invasions. 9:117–125.