
Abstract
Abrin, obtained from the seeds of Abrus precatorius plant, is a potent toxin belonging to the family of type II ribosome-inactivating proteins. Recently, a recombinant vaccine consisting of the A subunits of abrin and its homolog Abrus precatorius agglutinin (APA) was demonstrated to protect mice from abrin lethality. Toward identifying neutralizing epitopes recognized during this response, we generated monoclonal antibodies against the proposed vaccine candidate. One antibody, namely A7C4, the corresponding epitope of which was found to be distal to the active site of the enzymatic A chain, prevented abrin-mediated toxicity on cells and abrin-induced lethality in mice but did not inhibit the catalytic activity of the A chain. The in vivo protection conferred by monoclonal antibody A7C4 highlights the potential use of this antibody as a promising immunotherapeutic.
Abbreviations
| mAb | = | Monoclonal antibody |
| ABA | = | Abrin A chain |
| APA | = | Abrus precatorius agglutinin |
| APAA | = | Abrus precatorius agglutinin A chain |
| Ig | = | Immunoglobulin |
| kDa | = | kilo Dalton |
| LD50 | = | 50% |
| lethal | = | dose |
| RIP | = | Ribosome inactivating protein |
| ELISA | = | Enzyme-linked immunosorbent assay |
Introduction
Plant toxins hold considerable potential as bioweapons for military or terrorist use.Citation1 Abrin, of the plant Abrus precatorius is one of the most lethal plant toxins known. Owing to its high toxicity, relative ease of purification and accessibility, it is considered a biohazardous agent.Citation2 Abrin belongs to the family of type II ribosome inactivating proteins (RIPs). It is a heterodimeric protein consisting of the A chain which harbors the N-glycosidase activity and B chain which is a galactose-specific lectin.Citation3,4 The B chain binds to the cell surface terminal galactose residues and facilitates the entry of the toxin inside the cell. The A chain, depurinates the universally conserved α-sarcin loop of the 28S rRNA and inhibits protein synthesisCitation5 that activates the unfolded protein response leading to apoptosis.Citation6,7
Despite the high toxicity, treatment for abrin poisoning is symptomatic.Citation8 Currently there are no antidotes available for managing abrin poisoning. Certain chemical compounds such as N-acetyl-L-cysteine and Trolox which prevented abrin-induced cell death have been proposed for management of abrin intoxication, although the effects of these compounds in vivo have not been examined.Citation2 On the other hand, passive immunization with neutralizing antibodies has proven to be a specific and effective mode of defense against poisoning by several biological toxins.Citation9-12 The only known neutralizing monoclonal antibody (mAb) namely D6F10 for abrin was reported from our laboratory.Citation13 This mAb was found to inhibit the enzymatic activity of the abrin A chain in vitro and also conferred protection to mice in vivo.Citation13,14 Toward mapping the epitope corresponding to this mAb, at first various overlapping truncated forms of the abrin A chain were constructed and examined for binding to the mAb. We could conclude that the epitope lay in the amino acids 1-123 of ABA. Further truncations resulted in collapse of the protein structure.Citation14 Therefore, a chimeric protein consisting of 1-123 amino acids of abrin A chain (ABA) and 124-175 amino acids of the A chain of Abrus precatorius agglutinin (APAA) was constructed.Citation14 The recombinant construct retained the folding domains of ABA, enabling us to carry out mutagenesis studies to identify the key amino residues. Since the chimera included the epitope corresponding to the mAb D6F10 it was explored as a potential vaccine candidate (unpublished data). Although demonstrated to be effective against administration of lethal doses of abrin in mice, the epitopes corresponding to the response against the chimera are not known. In an effort to delineate the neutralizing epitopes on abrin, we established hybridoma using the splenocytes from mice immunized with the chimera and characterized the mAbs. In this study we report one mAb, namely A7C4 which inhibited toxin activity on cells and more importantly, in mice. Toward understanding the mechanism of immunoneutralization of abrin by this antibody, we mapped the core epitope recognized by mAb A7C4. Interestingly, the mAb A7C4 neither prevented the binding of abrin to cell surface nor did it inhibit the enzymatic activity of ABA as determined in cell free system, thereby suggesting that mAb A7C4 rescues abrin toxicity by a different mechanism.
Results
Reactivity of mAb A7C4 to different RIPs
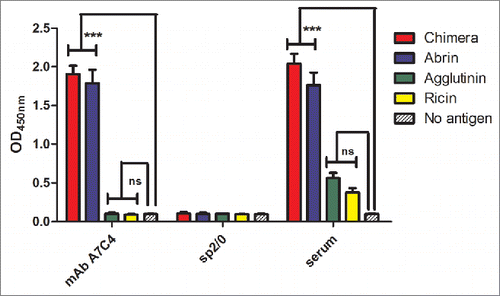
Twelve mAbs were generated to the ABA-APAA chimera. Of these the mAb A7C4 was selected based on its ability to rescue cells from abrin-mediated cytotoxicity. MAb A7C4 bound to abrin but not to APA in ELISA suggesting that the epitope of mAb A7C4 lies within 1 to 123 amino acids of abrin (). The mAb did not bind to the closely related type II RIP ricin, demonstrating that mAb A7C4 recognizes an epitope unique to abrin.
Figure 1. Reactivity of mAb A7C4 to abrin, agglutinin and ricin. Wells of ELISA plates were coated with 500 ng/100 µl per well of any one of the proteins, chimera, abrin, APA or ricin and incubated with 100 µl of the hybridoma culture supernatants for 2 h, followed by incubation with the secondary antibody-HRP conjugate. After adding the TMB substrate and stopping the reaction with 1M H2SO4, readings were taken at 450 nm. Polyclonal mouse antiserum to the chimera, and no-antigen wells served as the positive and negative control respectively. Each bar represents the mean ±SEM. Statistical analysis was carried out using One way ANOVA followed by Turkey's multiple comparison test (*p < 0.05).
Neutralization of abrin-induced cytotoxicity by mAb A7C4
The apoptotic population in cells treated with either abrin alone or along with mAb A7C4 was measured. HeLa (adherent culture) and Jurkat (suspension culture) cell lines were taken for this study. The choice of the two was based on the studies carried out earlier in our laboratory to understand the mechanism of abrin neutralization by mAb D6F10Citation14 and the signaling triggered by abrin.Citation7 As shown in the percentage of the apoptotic population in the samples treated with abrin along with mAb A7C4 was negligible compared to the controls in both cell lines demonstrating the ability of mAb A7C4 to rescue cells from abrin-mediated cytotoxicity. Moreover, mAb A7C4 when added to cells along with abrin, rescued cells from abrin-mediated inhibition of protein synthesis in a dose dependent manner ().
Figure 2. Neutralization of abrin cytotoxicity by mAb A7C4. HeLa / Jurkat cells were treated for 36 hr with abrin alone, abrin with mAb A7C4, or abrin with a control antibody and stained with propidium iodide as described in Materials and Methods. The apoptotic population was then measured using FACScan. Each bar represents the mean ±SEM.Statistical analysis was carried out using One way ANOVA followed by Turkey's multiple comparison test (*p < 0.05).
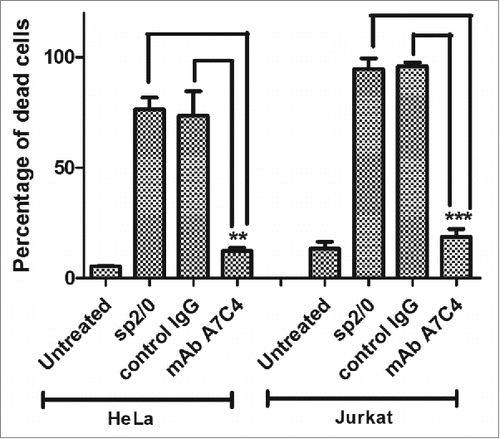
Figure 3. Rescue from abrin-mediated inhibition of protein synthesis in cells by mAb A7C4. HeLa cells were cultured in the presence of abrin alone or along with mAb A7C4 for 7 hr after which the cells were starved of leucine for 1 hr. Cells were then pulsed with [3H]-leucine and processed as described earlier. Each bar represents the mean ±SEM. Statistical analysis was carried out using One way ANOVA followed by Turkey's multiple comparison test (*p < 0.05).
![Figure 3. Rescue from abrin-mediated inhibition of protein synthesis in cells by mAb A7C4. HeLa cells were cultured in the presence of abrin alone or along with mAb A7C4 for 7 hr after which the cells were starved of leucine for 1 hr. Cells were then pulsed with [3H]-leucine and processed as described earlier. Each bar represents the mean ±SEM. Statistical analysis was carried out using One way ANOVA followed by Turkey's multiple comparison test (*p < 0.05).](/cms/asset/4b55b38f-cd3a-4bec-986d-91b4137cd3c6/khvi_a_1067741_f0003_b.gif)
Rescue from abrin-induced lethality in mice by mAb A7C4
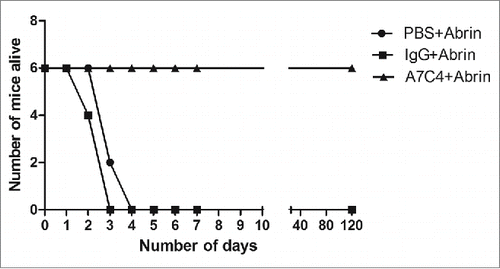
In order to assess the in vivo protective efficacy of mAb A7C4, mice were injected with 5 × LD50 of abrin intraperitoneally subsequent to administration of mAb A7C4 or a control IgG1 antibody. The mice were observed closely Administration of mAb A7C4 rescued 100% of mice from lethality (), while all the mice in other groups died within 4 days of abrin challenge.
Figure 4. In vivo protection from abrin-mediated lethality conferred by mAb A7C4. 6-8 weeks old BALB/c mice were injected intraperitoneally with 5xLD50 abrin after administration of mAb A7C4 or a control IgG1 antibody or PBS and their survival monitored.
Epitope mapping
mAb A7C4 does not bind APA in ELISA () nor Western blot (data not shown) suggesting that the core epitope of mAb A7C4 lies in the abrin part of the chimera, i.e 1-123 amino acids of abrin A chain. Further, the binding of mAb A7C4 to the overlapping truncated proteins, ABA1-100 and ABA76-175 and also the full length ABA was tested in Western blot. As shown in , mAb A7C4 recognized the full length ABA but neither of the truncated fragments. This suggested that the mAb A7C4 recognizes a conformational epitope. The refolding of ABA on the nitrocellulose membrane, as reported earlier with few other proteins,Citation15 appears to be a possible explanation for this observation.
Figure 5. Mapping the epitope of mAb A7C4. Different truncated forms of ABA and mutated forms of the chimera as depicted, were expressed as GST-tagged proteins in E.coli BL21 pLys strain and the lysate electrophoresed on 12.5% SDS-PAGE, transferred to nitrocellulose paper and immunoblotted with mAb A7C4 or with anti-GST antibody. The rest of the procedure followed was as described under ‘Materials and Methods ’.
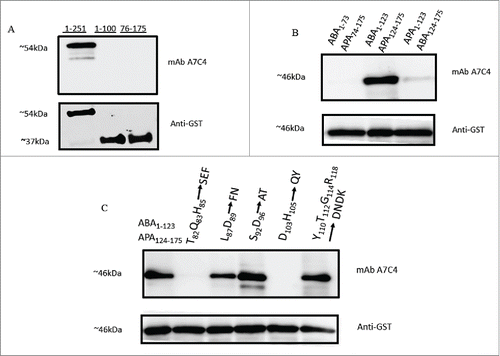
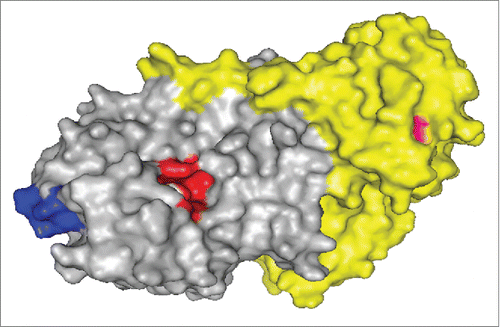
Though ABA and APAA share 67% sequence identity, Citation16 the observation that mAb A7C4 does not recognize the sequence of APAA suggests that the core epitope comprises majorly of the amino acid sequences that are unique to ABA only. The absence of binding to the construct APA1-123ABA124-175 () in which the first 123 amino acids of the abrin chimera (ABA1-123APA124-175) are replaced with the sequence of APAA, further corroborates this assertion. In line with this argument, the absence of binding of mAb A7C4 to the chimeric construct ABA1–73APA74–175 () would mean that the amino acids present in the region 74-123 which are unique to ABA are a part of the epitope corresponding to the mAb A7C4. Following this rationale, the ability of mAb A7C4 to recognize different mutant forms of chimera in which the amino acids unique to ABA have been replaced with the corresponding ones in the sequence of APAA, was then tested. As shown in , the mAb did not bind to the mutants T82Q83H85 to SEF and D103H105 to QY, thereby showing clearly that the amino acids Thr82, Gln83, His85, Asp103 and His105 are crucial for the binding of mAb A7C4 to abrin. depicts the epitopic region on the surface of abrin, the active site cleft and one of the galactose binding region.
Figure 6. Schematic representation of the epitopic region of mAb A7C4. The epitopic region (blue) of mAb A7C4 is depicted with respect to the active site cleft (red) on the surface of abrin (gray: abrin A chain; yellow: abrin B chain). The residue Asn-160 near one of galactose binding pockets of the B chain is represented in pink. The image was generated using open-source PyMOL software (PDB code: 1ABR).
MAb A7C4 does not inhibit the enzymatic activity of abrin A chain
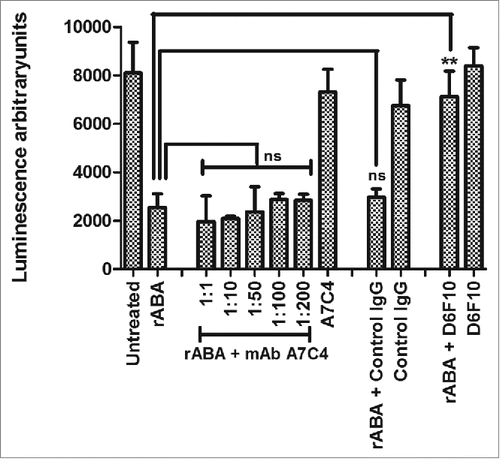
To determine whether mAb A7C4 neutralizes abrin toxicity by possibly inducing conformational changes in its enzymatic region, a cell-free system for translation of the luciferase mRNA to a functional protein was used to compare the enzymatic activity of abrin in the presence of the mAb A7C4. Synthesis of an active luciferase was detected by the luminescence observed upon addition of luciferin substrate. As expected, there was significantly lower luminescence observed in the presence of ABA () due to inhibition of protein synthesis. Presence of mAb A7C4 did not rescue the inhibition, thereby establishing that the mAb A7C4 does not interfere with the enzymatic activity of abrin unlike the earlier reported abrin-specific mAb D6F10.Citation14
Figure 7. Enzymatic activity of abrin in the presence of mAb A7C4. Rabbit reticulocyte lysate containing amino acids and luciferase mRNA was treated with either 200 pM recombinant ABA (rABA) alone or in the presence of varying molar concentrations of the mAb A7C4 for 1 h at 37°C followed by addition of the luciferin substrate. The rescue of inhibition of protein synthesis in the test samples by the mAbs was evaluated as the luminescence obtained in comparison with the rABA control. The figure represents data obtained from 3 different experiments. Similar treatments were performed with abrin in the presence of a control IgG1 antibody (200 fold molar excess over abrin). Treatment with abrin in the presence of mAb D6F10 (200 fold molar excess over abrin) served as a positive control.
MAb A7C4 does not affect the cell surface binding of abrin
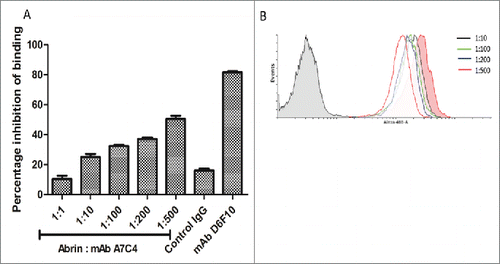
FACScan analysis was carried out to determine the extent of cell surface binding of labeled abrin in the presence of mAb A7C4. As shown in , there was no shift in the fluorescence peak in the presence of mAb even at concentrations as high as 200 fold molar excess of mAb A7C4 over abrin. These results demonstrate that mAb A7C4 does not neutralize by inhibiting the attachment of abrin to cell surface.
Figure 8. Binding of abrin on cell surface in the presence of mAb A7C4. (A) HeLa cells were treated with either Alexa 488- labeled abrin alone or along with different concentrations of mAb A7C4 for 1 hr, washed twice with ice cold PBS and analyzed by FACScan. Similar treatment with a control IgG1 antibody and mAb D6F10 (1:200 molar ratio of abrin : antibody) was performed to serve as negative and positive controls respectively. (B) Histogram of the representative of triplicate treatments performed is depicted.
Abrin internalizes in cells bound to mAb A7C4
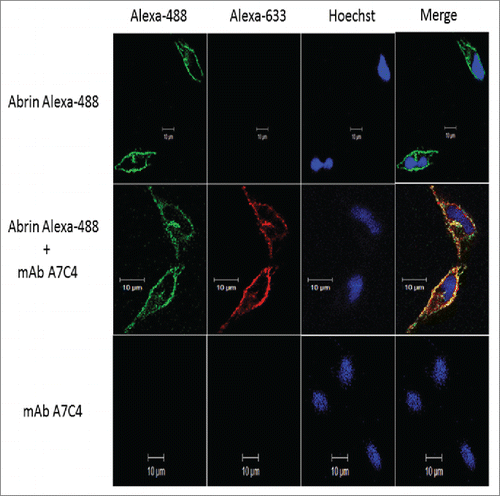
To examine whether mAb A7C4 inhibits the entry of abrin inside cells, we analyzed the cells treated with abrin and the mAb by confocal microscopy. Validating our previous observation from flow cytometric analysis (), abrin was found to bind to the cell surface in the presence of the mAb (). Furthermore, abrin internalized in the cells along with the mAb A7C4 thereby demonstrating that the mAb does not interfere with the entry of abrin in cells. MAb A7C4 was found to localize on the cell surface and internalize but only when bound to abrin. Taken together these results suggest that the mAb A7C4 might function intracellularly to rescue abrin-toxicity.
Figure 9. Internalization of abrin in cells in the presence of mAb A7C4. Hela cells were treated with Alexa 488-labeled abrin alone or mAb A7C4 alone or a mixture of both (1:100 molar ratio of abrin:mAb) for 1 hr and fixed with paraformaldehyde. Mab A7C4 was detected using Alexa-688 labeled anti-mouse IgG and the nuclei stained with Hoechst-33342 dye and visualized under Zeiss LSM 510 Meta Laser Confocal Scanning microscope (Carl Zeiss Foundation, Germany).
Discussion
The knowledge of neutralizing epitopes on lethal toxins is crucial for designing strategies to counter the toxicity. Abrin is a highly lethal protein obtained from the seeds of Abrus precatorius with a toxicity higher than its sister protein ricin.Citation17 Recently a chimeric protein comprising of the A subunits of abrin (ABA) and its homolog Abrus precatorius agglutinin (APA) was proposed in our laboratory as a potential vaccine candidate (unpublished data). Mice immunized with the abrin chimera were protected completely from lethal doses of abrin, attracting interest in studying the epitopes, in particular, neutralizing epitopes recognized by antibodies elicited by this protein. B cell hybridoma were established from mice immunized with this chimeric protein and the mAbs generated were screened for their ability to rescue cells from abrin-mediated cytotoxicity. The experiment led to the identification of the neutralizing mAb A7C4. Of additional interest is the protection conferred by passive immunization of mice with mAb A7C4 prior to administration of lethal dose of abrin which urged us to delineate the epitope of this antibody.
Studies to determine the reactivity of mAb A7C4 to the RIPs abrin and APA demonstrated that in spite of 67% sequence identity between ABA and APAA,Citation16 the core epitope of mAb A7C4 is restricted to amino acid sequence unique to abrin A chain within 1- 123 amino acids. Moreover, mAb A7C4 did not bind to the chimeric construct ABA1–73APA74–175 suggesting that the core epitope lies within 74–123 amino acids of abrin. Mutational analysis of the amino acids in this region clearly established that the amino acids Thr82, Gln83, His85, Asp103 and His105 are essential for the binding of A7C4 to abrin.
Abrin A chain is an RNA-N-glycosidase which plays a major role in inhibiting protein synthesisCitation3,4 leading to apoptosis.Citation18 Therefore, we analyzed the enzymatic actvitity of ABA in the presence of mAb A7C4. Visualization of the epitopic region on the surface of ABA showed that these residues are not proximal to the active site. Lack of inhibition of the enzymatic activity of ABA in the presence of mAb A7C4 validated this observation. Previously, the only known neutralizing antibody (mAb D6F10) to ABA was demonstrated to inhibit the binding of the toxin to the cell surface at higher concentrations.Citation14 mAb A7C4 failed to inhibit the attachment of abrin to the cell surface, corroborating well with the fact that the epitope of this antibody is present away from the 2 galactose binding pockets of the B chain. Furthermore the presence of internalized antibody in the cells suggests a possible intracellular neutralization mechanism. Antibodies to ricin, a sister protein of abrin, have been demonstrated to exert neutralization by hindering cell surface attachment of ricinCitation19 or inhibiting the catalytic activity of the A chainCitation20 or interfering with the reduction of the holotoxin to A and B chains.Citation21 It is less likely that mAb A7C4 acts via the last mechanism as the epitopic region is present away from the interface between the A and B chains. All the observations mentioned above strongly suggest an alternate mechanism of neutralization of abrin by the mAb A7C4. It is quite possible that mAb A7C4 alters the intracellular trafficking of abrin to the cytosol as observed earlier in case of neutralizing antibodies to ricinCitation22,23 although experimental evidence is required to confirm this hypothesis.
Taken together, we have reported a mAb, namely A7C4, to an ABA-APAA chimera, which inhibits abrin toxicity in cells as well as in vivo in mice. Abrin is a highly lethal toxin and its' potential use in biowarfare is a cause of major concern.Citation14 Neutralizing antibodies have been passively administered to manage several toxins.Citation24-26 The ability of mAb A7C4 to inhibit abrin toxicity in vivo ascertains its potential for therapy. So far only one neutralizing epitope on abrin has been reported.Citation14 This was found to be in proximity to the active site cleft of the A subunit and hence the antibody to this epitope was observed to interfere with the enzymatic activity of abrin.Citation14 In contrast, the epitope of mAb A7C4 is positioned away from the active site. This demonstrates that antibodies to abrin A chain which do not interfere with the N-glycosidase activity or the cell surface binding are still effective in neutralizing abrin toxicity. Studies are underway to elucidate the mechanism by which mAb A7C4 acts to inhibit abrin toxicity.
Materials and Methods
Reagents
Dulbecco's modified Eagle's medium(DMEM), Iscove's modified Dulbecco's medium(IMDM), Roswell Park Memorial Institute (RPMI-1640), propidium iodide (PI), bovine serum albumin (BSA) and RNase A were purchased from Sigma Aldrich (St.Louis, MO). Glutamax, Trypsin-EDTA and Fetal bovine serum (FBS) were obtained from Gibco BRL, USA. Isopropyl thiogalactoside (IPTG) was purchased from Fermentas (USA). In vitro translation kit was obtained from Promega Pvt. Ltd (Singapore). All the sterile plastic-ware was from Nunc™ (Denmark).
Cell lines
HeLa cells were cultured in DMEM, Sp2/0 myeloma and hybridoma cells in IMDM, and JR4 clone of Jurkat cells (human T-cell line) in RPMI-1640 at 37°C in a humidified chamber with 5% CO2. All the media were supplemented with 10% (v/v) FBS, 1mM Glutamax, 100 I.U./ml penicillin, 100 µg streptomycin, 50 µg gentamycin and 5 U/ml nystatin.
Establishment of chimera specific hybridoma
The protocol used for establishing hybridoma generating antibodies specific to the abrin chimera was essentially the same as that described by Gangatirkar et al.Citation27 Female BALB/c mice (6-8 weeks old) were administered 40 µg of the abrin chimera emulsified in Freund's complete adjuvant. Two booster doses of 20 µg of the antigen in Freund's incomplete adjuvant were given at 21 day intervals after which the animals were rested for a month. Three days prior to sacrificing, the mice were injected intraperitoneally with 200 µg of the antigen in PBS. The splenocytes were collected, washed and fused with Sp2/0 myeloma cells in the ratio 5:1 (splenocytes:Sp2/0) using PEG 4000 (Merck, Rahway, NJ). The fused cells were plated in medium supplemented with 20% (v/v) FBS, 50 µM β-mercaptoethanol and HAT (10 mM hypoxanthine, 40 µM aminopterin and 1.6 mM thymidine) and 12 days later tested for secretion of abrin chimera-specific antibodies using indirect ELISA. The positive clones were expanded and subcloned to monoclonality using the method of limiting dilution.
Plant proteins and antibodies
Abrin and APA were purified from the mature seeds as described earlier.Citation16,28 Monoclonal antibodies were purified from the corresponding hybridoma culture supernatants using Protein A-Sepharose affinity chromatography29 and the concentration of the purified proteins estimated by Lowry's method.
Indirect ELISA
ELISA plates were coated with proteins (500 ng/100 µl per well) in phosphate buffer, pH 7.2 containing 150 mM NaCl (PBS) overnight at RT. The unoccupied sites were blocked with 0.5% (w/v) gelatin in PBS for 1 hr at RT. Hybridoma culture supernatants (100 µl) or purified antibody was added and incubated for 1.5 hr at RT, followed by washing with PBS containing 0.05 % Tween-20 and subsequent incubation with secondary antibody (rabbit anti-mouse antibody conjugated to horse radish peroxidase (HRP), Dako, Denmark) for 45 min. The plates were washed and the enzyme activity retained in the well was determined with 100 μl substrate 3,3′,5,5′-tetra methyl benzidine (TMB) - H2O2. The reaction was stopped by the addition of 50 μl 2N H2SO4. The absorbance was measured at 450 nm using an ELISA reader.
Propidium iodide staining of abrin treated cells
HeLa cells, 0.5 million / 500 µl medium / well were plated in a 12 well plate and allowed to adhere overnight. Abrin (10 ng/ml) alone or along with hybridoma culture supernatant (100 µl) containing antibodies was added and incubated for 36 hr after which the cells were harvested, washed twice with PBS and fixed using 70% ethanol overnight. After washing, the cells were resuspended in 300 µl PBS containing propidium iodide (20 µg/ml) and RNase (50 µg/ml) and incubated for 2 hr at 42 °C. The cells were then washed with PBS and the proportion of hypodiploid population was estimated using fluorescence activated cell scan. The percentage of dead cell population in different samples was calculated considering the value obtained for abrin treated cell as 100%. Similar treatments were carried out with Jurkat cells, but treated with a lower dose of abrin (1ng/ml).
Analysis of protein synthesis in cells
HeLa cells, 0.2 million / 200 µl /well were plated in 24 well plate and allowed to adhere overnight. Abrin (10 ng/ml) alone or along with varying concentrations of antibodies was added to cells and incubated for 7 hr. The medium was replaced with 250 µl of leucine-free RPMI (US Biological). Two hr later the cells were pulsed with 0.3 µCi [3H]-leucine (BRIT, India) for 1 hr and the total protein was precipitated using 5% (w/v) trichloroacetic acid at 4°C overnight. The precipitates were then washed with 20% ethanol, dried and dissolved in 200 µl of 1% SDS in 0.1 N NaOH and the radioactivity incorporated was measured in a liquid scintillation counter (Beckman Coulter Inc.., USA).
In vivo rescue from abrin-induced lethality
Experiments using mice were performed with strict adherence to the guidelines established by the Indian Institute of Science Animal Ethics Committee with prior approval for the experimental protocol. Mice were housed in 12 hr night-day cycle at controlled temperature (24°C) and humidity and food ad libitum. Three groups of BALB/c mice were recruited, 6 mice per group. Mice were injected intraperitoneally with 200 fold molar excess of antibodies over abrin or equal volume of PBS. After 90 min, abrin (5 × LD50) was injected into the mice by the same route. The mice were monitored closely.
Expression and immunoblot analysis of recombinant proteins
Generation and expression of the chimeric constructs, overlapping truncation and mutant forms of the chimeric abrin have been described elsewhere.Citation14 Lysates of E.coli cells induced for the expression of the desired proteins (using IPTG) were subjected to electrophoresis on 12.5% SDS-polyacrylamide gel under denaturing and reducing conditions and then transferred onto nitrocellulose membrane. The membrane was incubated for 2 hr with 5% (w/v) skimmed milk in PBS and subsequently incubated with hybridoma culture supernatants containing the primary antibodies (A7C4 or anti-GST antibody) for 2 hr. Following this incubation, the membranes were washed and incubated with secondary antibody for 45 min and then washed and developed using ECL kit (Millipore).
Luciferase assay for analyzing protein synthesis in cell-free system
Abrin (400 pM) alone or along with different concentrations of mAbs was added to rabbit reticulocyte lysate (RRL kit, Promega) cocktail containing total amino acids, RNase inhibitor and luciferase mRNA and incubated for 1 hr at 37°C. After the incubation, the reaction mixture was mixed with luciferin substrate and the extent of the enzyme luciferase synthesized was determined by measuring the luminescence in a luminometer.
Fluorescence activated cell scan (FACScan) analysis for studying the cell surface bound abrin
HeLa cells (0.2 million) were incubated with Alexa-488 (Invitrogen, USA) labeled abrin (250 ng/ml) alone or in the presence of different concentrations of mAbs for 1 hr on ice after which the cells were washed twice with ice-cold PBS and analyzed using FACScan (Becton Dickinson). Median fluorescence values of cells treated with labeled abrin along with or without mAbs were compared.
Confocal microscopy
0.02 million HeLa cells were plated on coverslips and allowed to adhere at 37°C. Normal goat serum (1:50 dilution) was used to block the Fc receptors on cells for 1 hr after which the cells were treated with abrin (Alexa-488 labeled) alone or in the presence of mAb A7C4 (1:100 molar ratio of abrin:mAb) or mAb A7C4 alone for 1 hr at 37°C. The cells were washed thrice with PBS and fixed with 4% paraformaldehyde followed by further washes with PBS. The fixed cells were incubated with Alexa-633 labeled anti-mouse IgG (Invitrogen, USA) for 1 hr and washed again with PBS. The nuclei were stained with Hoescht-33342 (5 µg/ml) for 7 min followed by further washes with PBS. The coverslip with the cells were mounted on slides using 10 µl mounting solution (90% glycerol containing Mowiol antifade, Calbiochem, USA) and visualized under Zeiss LSM 510 Meta Laser Confocal Scanning microscope (Carl Zeiss Foundation, Germany). The images were analyzed using Zeiss LSM image browser.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
MSK acknowledges the Department of Science and Technology, Government of India, for the KVPY fellowship, we thank the Department of Biotechnology, Government of India for the FACS and confocal facilities, and Shruthi KS for the technical assistance.
Funding
We thank the Department of Biotechnology, Government of India for financial support.
References
- Zhang X, Kuča K, Dohnal V, Dohnalová L, Wu Q, Wu C. Military potential of biological toxins. J Appl Biomed 2014; 12:63-77; http://dx.doi.org/10.1016/j.jab.2014.02.005
- Saxena N, L R PV, B B AS, Bhutia YD. Protective effects of certain pharmaceutical compounds against abrin induced cell death in Jurkat cell line. Int Immunopharmacol 2014; 21:412-25; PMID:24938881; http://dx.doi.org/10.1016/j.intimp.2014.05.028
- Hartley MR, Lord JM. Cytotoxic ribosome-inactivating lectins from plants. Biochim Biophys Acta 2004; 1701:1-14; http://dx.doi.org/10.1016/j.bbapap.2004.06.004
- Nielsen K, Boston RS. RIBOSOME-INACTIVATING PROTEINS: A Plant Perspective. Annu Rev Plant Physiol Plant Mol Biol 2001; 52:785-816; PMID:11337416; http://dx.doi.org/10.1146/annurev.arplant.52.1.785
- Stirpe F, Bailey S, Miller SP, Bodley JW. Modification of ribosomal RNA by ribosome-inactivating proteins from plants. Nucleic Acids Res 1988; 16:1349-57; PMID:3347493; http://dx.doi.org/10.1093/nar/16.4.1349
- Narayanan S, Surolia A, Karande AA. Ribosome-inactivating protein and apoptosis: abrin causes cell death via mitochondrial pathway in Jurkat cells. Biochem J 2004; 377:233-40; PMID:13129434; http://dx.doi.org/10.1042/BJ20030797
- Mishra R, Karande AA. Endoplasmic Reticulum Stress-Mediated Activation of p38 MAPK, Caspase-2 and Caspase-8 Leads to Abrin-Induced Apoptosis. PLoS ONE 2014; 9:e92586; PMID:24664279; http://dx.doi.org/10.1371/journal.pone.0092586
- Dickers KJ, Bradberry SM, Rice P, Griffiths GD, Vale JA. Abrin poisoning. Toxicol Rev 2003; 22:137-42; PMID:15181663; http://dx.doi.org/10.2165/00139709-200322030-00002
- Colombatti M, Johnson VG, Skopicki HA, Fendley B, Lewis MS, Youle RJ. Identification and characterization of a monoclonal antibody recognizing a galactose-binding domain of the toxin ricin. J Immunol 1987; 138:3339-44; PMID:2437188
- Nowakowski A, Wang C, Powers DB, Amersdorfer P, Smith TJ, Montgomery VA, Sheridan R, Blake R, Smith LA, Marks JD. Potent neutralization of botulinum neurotoxin by recombinant oligoclonal antibody. Proc Natl Acad Sci U S A 2002; 99:11346-50; PMID:12177434; http://dx.doi.org/10.1073/pnas.172229899
- Burnett JC, Henchal EA, Schmaljohn AL, Bavari S. The evolving field of biodefence: therapeutic developments and diagnostics. Nat Rev Drug Dis 2005; 4:281-97; PMID:15803193; http://dx.doi.org/10.1038/nrd1694
- Tzipori S, Sheoran A, Akiyoshi D, Donohue-Rolfe A, Trachtman H. Antibody therapy in the management of shiga toxin-induced hemolytic uremic syndrome. Clin Microbiol Rev 2004; 17:926-41, table of contents; PMID:15489355; http://dx.doi.org/10.1128/CMR.17.4.926-941.2004
- Surendranath K, Karande AA. A neutralizing antibody to the a chain of abrin inhibits abrin toxicity both in vitro and in vivo. Clin Vacc Immunol 2008; 15:737-43; PMID:18353919; http://dx.doi.org/10.1128/CVI.00254-07
- Bagaria S, Ponnalagu D, Bisht S, Karande AA. Mechanistic Insights into the Neutralization of Cytotoxic Abrin by the Monoclonal Antibody D6F10. PLoS ONE 2013; 8:e70273; PMID:23922965; http://dx.doi.org/10.1371/journal.pone.0070273
- Rao L, Jones DP, Nguyen LH, McMahan SA, Burgess RR. Epitope mapping using histidine-tagged protein fragments: application to Escherichia coli RNA polymerase sigma 70. Anal Biochem 1996; 241:173-9; PMID:8921183; http://dx.doi.org/10.1006/abio.1996.0395
- Bagaria A, Surendranath K, Ramagopal UA, Ramakumar S, Karande AA. Structure-function analysis and insights into the reduced toxicity of Abrus precatorius agglutinin I in relation to abrin. J Biol Chem 2006; 281:34465-74; PMID:16772301; http://dx.doi.org/10.1074/jbc.M601777200
- Stirpe F, Barbieri L, Battelli MG, Soria M, Lappi DA. Ribosome-inactivating proteins from plants: present status and future prospects. Biotechnology (N Y) 1992; 10:405-12; PMID:1368484; http://dx.doi.org/10.1038/nbt0492-405
- Mishra R, Kumar MS, Karande AA. Inhibition of protein synthesis leading to unfolded protein response is the major event in abrin-mediated apoptosis. Mol Cell Biochem 2015; 403:255-65; PMID:25753921; http://dx.doi.org/10.1007/s11010-015-2355-9
- McGuinness CR, Mantis NJ. Characterization of a novel high-affinity monoclonal immunoglobulin G antibody against the ricin B subunit. Infect Immun 2006; 74:3463-70; PMID:16714577; http://dx.doi.org/10.1128/IAI.00324-06
- Neal LM, O'Hara J, Brey RN, 3rd, Mantis NJ. A monoclonal immunoglobulin G antibody directed against an immunodominant linear epitope on the ricin A chain confers systemic and mucosal immunity to ricin. Infect Immun 2010; 78:552-61; PMID:19858297; http://dx.doi.org/10.1128/IAI.00796-09
- O'Hara JM, Mantis NJ. Neutralizing monoclonal antibodies against ricin's enzymatic subunit interfere with protein disulfide isomerase-mediated reduction of ricin holotoxin in vitro. J Immunol Methods 2013; 395:71-8; PMID:23774033; http://dx.doi.org/10.1016/j.jim.2013.06.004
- Song K, Mize RR, Marrero L, Corti M, Kirk JM, Pincus SH. Antibody to Ricin A Chain Hinders Intracellular Routing of Toxin and Protects Cells Even after Toxin Has Been Internalized. PLoS ONE 2013; 8:e62417
- Yermakova A, Klokk TI, Cole R, Sandvig K, Mantis NJ. Antibody-mediated inhibition of ricin toxin retrograde transport. MBio 2014; 5:e00995; PMID:24713323; http://dx.doi.org/10.1128/mBio.00995-13
- Islam MS, Stimson WH. Production and characterization of monoclonal antibodies with therapeutic potential against Shiga toxin. J Clin Lab Immunol 1990; 33:11-6; PMID:1966939
- Lemley PV, Amanatides P, Wright DC. Identification and characterization of a monoclonal antibody that neutralizes ricin toxicity in vitro and in vivo. Hybridoma 1994; 13:417-21; PMID:7860097; http://dx.doi.org/10.1089/hyb.1994.13.417
- Behring, Kitasato. Ueber das Zustandekommen der Diphtherie-Immunität und der Tetanus-Immunität bei Thieren. Dtsch med Wochenschr 1890; 16:1113-4; http://dx.doi.org/10.1055/s-0029-1207589
- Gangatirkar P, Gangadharan S, Narendranath A, Nagpal S, Salunke DM, Karande AA. Monoclonal antibodies to gonadotropin-releasing hormone (GnRH) inhibit binding of the hormone to its receptor. Hybridoma and hybridomics 2002; 21:281-6; PMID:12193281; http://dx.doi.org/10.1089/153685902760213895
- Hegde R, Maiti TK, Podder SK. Purification and characterization of three toxins and two agglutinins from Abrus precatorius seed by using lactamyl-Sepharose affinity chromatography. Anal Biochem 1991; 194:101-9; PMID:1867374; http://dx.doi.org/10.1016/0003-2697(91)90156-N
- Grodzki AC, Berenstein E. Antibody purification: affinity chromatography - protein A and protein G Sepharose. Methods Mol Biol 2010; 588:33-41; http://dx.doi.org/10.1007/978-1-59745-324-0_5