

ABSTRACT
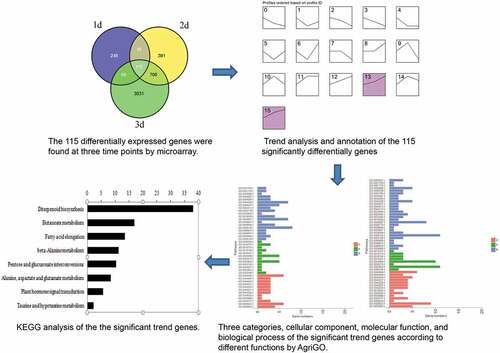
The main aim of the work is to study the regulation of gene expression in the interaction between rice and Magnaporthe oryzae by gene chip technology. In this study, we mainly focused on changes of gene expression at 24, 48, and 72 hours post-inoculation (hpi), through which we could conduct a more comprehensive analysis of rice blast-related genes in the process of infection. The results showed that the experimental groups contained 460, 1227, and 3937 significant differentially expressed genes at 24, 48, and 72 hpi, respectively. Furthermore, 115 significantly differentially expressed genes were identified in response to rice blast infection at all three time points. By annotating these 115 genes, they were divided into three categories: metabolic pathways, proteins or enzymes, and organelle components. As expected, many of these genes were known rice blast-related genes; however, we discovered new genes with high fold changes. Most of them encoded conserved hypothetical proteins, and some were hypothetically conserved genes. Our study may contribute to finding new resistance genes and understanding the mechanism of rice blast development.
GRAPHICAL ABSTRACT
Highlights
Some new signaling pathways involved in blast disease resistance are revealed.
Many new rice defense-related genes are found to involve in interaction between rice and M. oryzae.
Many conserved hypothetical proteins with unknown functions are found to involve in blast disease resistance.
Disclosure statement
No potential conflict of interest was reported by the authors.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (31370306), and the Special Fund for Basic Scientific Research of Central Colleges, South-Central University for Nationalities (CZY20035). We thank LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
Supplementary material
Supplemental data for this article can be accessed here.