

ABSTRACT
It is necessary to explore new molecules for the improvement of precise diagnosis and antitumor therapies in lung cancer. LncRNAs (long non-coding RNAs) play an important role in the regulation of cancer cell malignant behavior and tumor development. In this work, we found that a newly discovered lncRNA, lncRNA PGM5P4-AS1, was lower expressed in lung cancer tissues than adjacent tissues. Then, the lncRNA PGM5P4-AS1 was overexpressed or knocked-down in different lung cancer cells, and its effects on the malignant phenotypes were measured by 3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay, cell cycle assay, wound healing assay, and transwell assay. The results showed that the overexpression of PGM5P4-AS1 inhibited lung cancer cell proliferation, migration, and invasion activities, while these abilities were prominently promoted by the interference of PGM5P4-AS1. Further, the growth of lung cancer tumors in nude mice was also inhibited by PGM5P4-AS1 overexpression. In mechanism, PGM5P4-AS1 has the binding site of miR-1275 and could positively regulate the expression of LZTS3 via sponging miR-1275. In conclusion, PGM5P4-AS1 could be a potential precise diagnosis and therapeutic target biomarker of lung cancer.
GRAPHICAL ABSTRACT
Highlights:
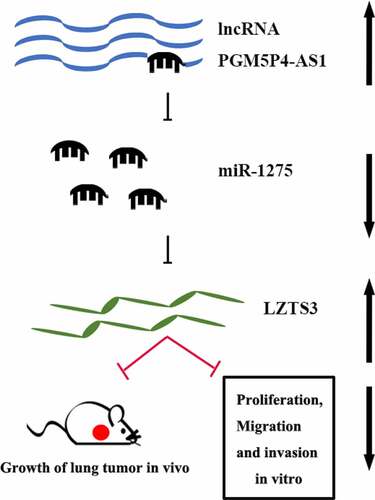
1. PGM5P4 AS1 was down-regulated in lung cancer tissues.
2. PGM5P4 AS1 inhibited lung cancer cell proliferation, migration, and invasion.
3. PGM5P4-AS1 suppressed the growth of lung cancer tumors in vivo.
4. PGM5P4-AS1 acted as a sponge of miR-1275 in lung cancer cells.
5. LZTS3 acted as a target of miR-1275 in lung cancer cells.
Acknowledgements
This research was supported by grants from the Key Project of Health Technology Research and Achievement Conversion in Hebei Province (Grant Number zh2018009) and the Technological Innovation Project of Cooperation Fund in Hebei Provincial Department of Science and Technology and Hebei Medical University (Grant Number 2020TXZH04).
Supplementary material
Supplemental data for this article can be accessed here.
Disclosure statement
No potential conflict of interest was reported by the authors.