

ABSTRACT
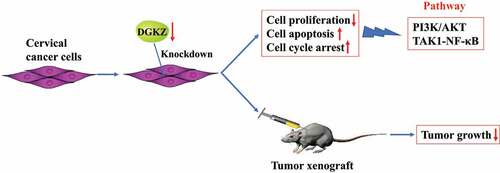
Diacylglycerol kinase zeta (DGKZ) participates in cancer progression. Here, the current work aims to identify the functional role of DGKZ in cervical cancer (CC). DGKZ expression in cervical cancer tissues and paired adjacent normal cervical tissues was assessed using Immunohistochemistry assay. SiHa and HeLa cells were transfected with lentivirus plasmids (sh-DGKZ or sh-NC) to evaluate the effects of DGKZ knockdown on cell proliferation, apoptosis and cell cycle distribution in vitro. Furthermore, BALB/c nude mice were injected subcutaneously with Lentivirus-sh-DGKZ-SiHa cells or Lentivirus-sh-NC-SiHa cells to analyze the influence of DGKZ silencing on tumor growth of CC in vivo. Moreover, the potential molecular mechanisms were predicted by GO and KEGG analysis and preliminarily explored through PathScan Analysis. Elevated DGKZ expression in cervical tumor was observed. Downregulation of DGKZ repressed proliferation and boosted apoptosis of SiHa and HeLa cells and induced cell cycle arrest at G0/G1 phase. In addition, Knockdown of DGKZ restrained tumor growth in tumor xenograft mice. Importantly, GO and KEGG analysis displayed that differentially expressed proteins induced by silence of DGKZ were mostly enriched in autophagy or mitophagy, indicating that the functions of DGKZ on cell proliferation and tumor growth may be associated with autophagy or mitophagy. PathScan analysis presented that PI3K-AKT and TAK1-NF-κB signaling pathways were prominently inhibited in SiHa cells transfected with sh-DGKZ. In summary, downregulation of DGKZ impeded cell proliferation, boosted cell apoptosis and induced cell cycle arrest to suppress tumorigenesis and progression of cervical cancer.
Graphical Abstract
Availability of data and material
All datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval
All procedures performed in this study involving human participants were in accordance with the ethical standards of institutional research ethics committee of Taizhou Hospital of Zhejiang Province and Helsinki Declaration (1964). All animal study was approved by the committee of Shanghai GeneChem Co. Ltd. according to the guidelines of the Institutional Animal Care and Welfare Committee of Xi’an Jiaotong University of Medicine.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Disclosure statement
No potential conflict of interest was reported by the author(s).