

ABSTRACT
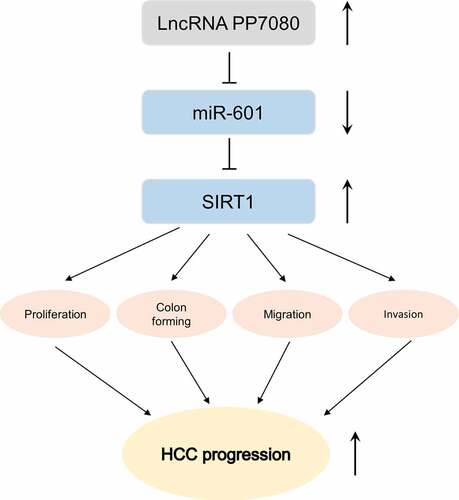
Hepatocellular carcinoma (HCC) is the most common primary liver malignancy in adults, ranking the second leading cause of cancer-related death. To date, the underlying mechanisms of HCC pathogenesis are still unclear. Recently, more and more studies have reported that long noncoding RNAs (lncRNAs) are involved in the occurrence and development of HCC. This study aims to investigate the expressions, clinical significance and roles of lncRNA PP7080 in HCC. We analyzed the transcriptome data of HCC cancer tissue (n = 369) and normal tissue (n = 50) in the TCGA database. We used the qRT-PCR method to detect the expression levels of lncRNA PP7080 in 40 pairs of HCC and adjacent tissues. The survival curve was drawn by KM-plotter. The changes of migration, invasion and proliferation of HCC cells were detected by transwall, CCK8 and colony forming assays, respectively. For the interaction between genes, we performed the luciferase activity assay to analyze. The expression of lncRNA PP7080 and miR-601 in cancer tissues of 40 cancer patients was analyzed by Pearson correlation. LncRNA PP7080 was highly expressed in HCC and predicted a poor prognosis. Luciferase activity assay identified lncRNA PP7080 as a molecular sponge for miR-601 in HCC cells. LncRNA PP7080 promoted HCC cells proliferation, migration and invasion by miR-601/SIRT1 signal axis. These results revealed lncRNA PP7080 effect in regulating miR-601/SIRT1 signal axis in the progression of HCC, indicating the important role of miR-601 in HCC pathogenesis.
Graphical abstract
KEYWORDS:
Acknowledges
The authors gratefully acknowledge the support of China Hepatitis Prevention and Control Foundation, Professor Wu Huiwen from Science and Technology Center of Fenyang College of Shanxi Medical University for his guidance.
Abbreviations
FBS: fatal bovine serun, BSA: Bovine Serum Albumin, TBST: TBST buffer, PBS: phosphate buffer saline, TNM: Tumor Node Metastasis, AFP: alpha fetoprotein, WB: western blot.
Supplementary material
Supplemental data for this article can be accessed here
Highlights
LncRNA PP7080 was highly expressed in HCC and predicted a poor prognosis.
LncRNA PP7080 directly target miR-601 as a molecular sponge.
LncRNA PP7080 promoted HCC cells proliferation, migration and invasion by downregulate miR601/SIRT1 signal axis.
LncRNA PP7080 knockdown led to inhibition of HCC progression.
Disclosure statement
The authors have no competing interests to declare.
Author contribution
Weifang Song, Ruiqiang Yao, Wenhui Zhang, Xinli Hu, Ting Shi, Meijiao Wang and Haijuan Zhang contributed to the conception of the study;
Weifang Song, Ruiqiang Yao, Wenhui Zhang, Xinli Hu and Meijiao Wang performed the experiment;
Weifang Song, Wenhui Zhang, Xinli Hu Meijiao Wang and Haijuan Zhang contributed significantly to analysis and manuscript preparation;
Weifang Song, Ruiqiang Yao, Wenhui Zhang, Xinli Hu, Ting Shi, Meijiao Wang and Haijuan Zhang performed the data analyses and wrote the manuscript.
Ethical statement
None.