

ABSTRACT
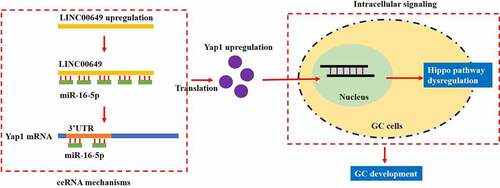
Although long non-coding RNA (LncRNA) LINC00649 is reported to be closely associated with acute myeloid leukemia (AML), prostate cancer and colorectal cancer, its role in regulating other types of cancer, such as gastric cancer (GC), has not been studied. This study analyzed the expression status of LINC00649 in GC tissues and cells by performing Real-Time qPCR analysis, and we found that LINC00649 tended to be enriched in cancerous tissues and cells but not in their normal counterparts, which were supported by the data from TCGA dataset. Next, by performing the gain- and loss-of-function experiments, we expectedly found that LINC00649 acted as an oncogene to accelerate GC cell proliferation, migration and epithelial-mesenchymal transition (EMT) in vitro and promote its tumorigenesis in vivo. Moreover, the online miRDB software predicted that miR-16-5p bound to both LINC00649 and 3ʹ untranslated region (3ʹUTR) of YAP1 mRNA, which were validated by the following dual-luciferase reporter gene system assay and RNA pull-down assay. Finally, we proved that LINC00649 exerted its tumor-promoting effects in GC by regulating the miR-16-5p/YES-associated protein 1 (YAP1)/Hippo pathway. Mechanistically, knock-down of LINC00649 suppressed YAP1 expressions by releasing miR-16-5p, resulting in the recovery of the Hippo pathway, which suppressed the expression levels of the downstream oncogenes, including EGFR, SOX2 and OCT4, leading to the inhibition of the malignant phenotypes in GC cells. In conclusion, this study, for the first time, evidenced that LINC00649 promoted GC progression by targeting the miR-16-5p/YAP1/Hippo signaling pathway, which provided potential diagnostic and therapeutic indicators for GC treatment for clinical utilization.
GRAPHICAL ABSTRACT
Highlights
LINC00649 promotes cell proliferation, migration and EMT in GC.
LINC00649 positively regulates YAP1 expressions in GC cells by sponging miR-16-5p.
Silencing of LINC00649 suppresses GC malignancy via modulating the miR-16-5p/YAP1 axis.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed here.