

ABSTRACT
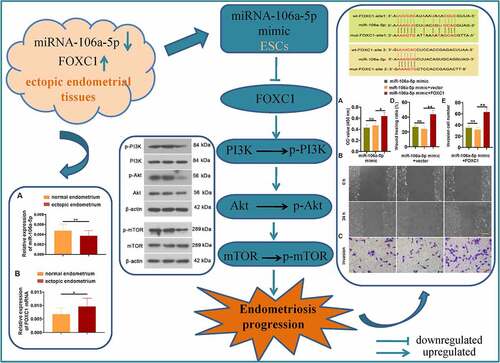
Emerging evidence has exhibited an obvious decreased expression of miR-106a-5p in the ectopic endometrial tissue of endometriosis (EMS) patients. Thus far, the pathophysiological function of miR-106a-5p in EMS is unknown. A previous study showed an increased FOXC1 expression in the ectopic endometrial tissue of patients with EMS. Moreover, we found that there was a binding site of miR-106a-5p on the 3ʹUTR of FOXC1 through bioinformatics predictions. Hence, we speculated that miR-106a-5p might affect the development of EMS via targeting FOXC1. We first showed a decreased level of miR-106a-5p and an increased level of FOXC1 mRNA in ectopic endometrial tissues compared with normal tissues. Functionally, we transfected ectopic endometrial stromal cells (ESCs) with miR-106a-5p mimics or NC mimics and indicated an inhibitory role of miR-106a-5p on ESC proliferation, invasion, and migration. Mechanistically, FOXC1 was found to be a target gene of miR-106a-5p. To confirm whether miR-106a-5p exerted an inhibitory activity in ESCs via targeting FOXC1, miR-106a-5p mimic was co-transfected into ESCs with the FOXC1-plasmid or vector. We found that FOXC1 overexpression evidently reversed the results caused by a miR-106a-5p mimic in ESCs. Additionally, our results demonstrated that miR-106a-5p mimic inhibited the expression of p-Akt and p-PI3K. Collectively, these results revealed that miR-106a-5p inhibited the proliferative, migratory, and invasive ability of ESCs via directly binding to FOXC1, likely through the suppression of the PI3K and its downstream signaling pathway, which offered a potential and novel therapeutic strategy for EMS treatment.
Graphical abstract
KEYWORDS:
Research highlights
miR-106a-5p mimic repressed ESC proliferation, invasion, and migration
miR-106a-5p bound to FOXC1
miR-106a-5p suppressed the PI3K and its downstream signaling pathway
miR-106a-5p inhibited EMS progression via targeting FOXC1
Data availability statement
The data used to support the findings in this study are available from the corresponding author upon request
Disclosure statement
The authors have no conflicts of interest to declare.
Supplementary material
Supplemental data for this article can be accessed here