

ABSTRACT
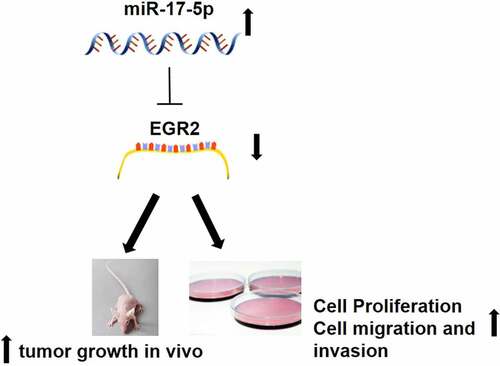
miR-17-5p has been proved that play important roles in many kinds of tumors progression. This study aimed at explore the function and mechanism of miR-17-5p in thyroid cancer (TC). RT-qPCR was used to detect miR-17-5p and Early growth response 2 (EGR2) expression in TC tissues and cells. CCK8 and colony formation assay were used to analyze cell proliferation. Cell migration and cell invasion was detected by Wound-healing assay and Transwell assay. Detection of protein expression using Western blot analysis. Dual-Luciferase assay was used to analyze the relationship between miR-17-5p and EGR2. In vivo experiment was performed by establishing Xenograft animal model to observe the function of miR-17-5p. We found that miR-17-5p is significantly increased in thyroid cancer tissues and cells. miR-17-5p inhibition repressed cell proliferation, clonal formation, cell migration, and cell invasion in thyroid carcinoma. Moreover, miR-17-5p inhibition suppressed tumorigenesis in vivo. Dual-Luciferase assay and Western blotting assay further proved that miR-17-5p has a negative regulation to EGR2. EGR2 was decreased in TC tissues and cells. Overexpressed EGR2 inhibited the development of thyroid carcinoma both vivo and in vivo. EGR2 knockdown remarkably decreased the anti-cancer effect of miR-17-5p inhibition. miR-17-5p is a thyroid cancer oncomir by modulation of the EGR2.
Graphical abstract
Highlights
1. miR-17-5p is significantly increased in thyroid cancer tissues and cells.
2. mR-17-5p promotes thyroid cancer tumorigenesis both in vitro and in vivo.
3. miR-17-5p is a thyroid cancer oncomir by modulation of the EGR2.
Disclosure statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical statement
This study was approved by The Affiliated Changzhou No.2 People’s Hospital of Nanjing Medical University ethics committee. All participants in this study signs a written informed consent.