

ABSTRACT
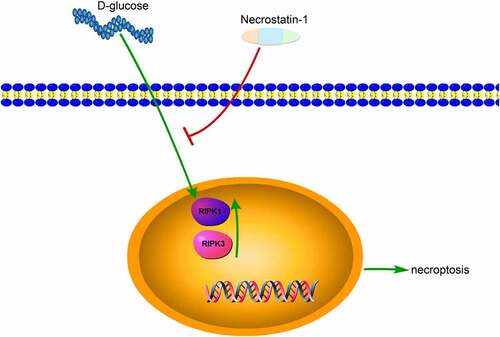
Diabetic retinopathy (DR) represents the most typical complication of type 2 diabetes mellitus and one of the most primary oculopathy causing blindness. However, the mechanism of DR remains unknown. RIPK1/RIPK3, as homologous serine/threonine kinases, are key elements in mediating necroptosis and may have functions in DR development. To clarify the relationship between DR and RIPK1/RIPK3, this study established a model of apoptosis using high-glucose induced RGCs, which were treated with 7.5, 19.5, and 35 mM D-glucose for 12, 24, and 48 h, respectively. Subsequently, the expression of RIPK1/RIPK3 was determined and the protective effect of necrostatin-1 on RGCs injury induced by high glucose was explored. The results demonstrated that the expression of RIPK1 and RIPK3 in the cells was increased markedly following 12 h treatment with 19.5 mM D-glucose. Additionally, following an addition of 100 μM necrostatin-1 in 19.5 mM D-glucose medium for RGCs treatment 12 h, the protein expression of RIPK1 and RIPK3 was decreased markedly, and the number of Nissl bodies in cells was increased substantially. The findings of the present study indicated that high glucose could induce the expression of RIPK1/RIPK3, and necrostatin-1 could effectively protect RGCs from D-glucose-induced cell necrosis.
Graphical Abstract
Highlights
This study established a model of apoptosis using high-glucose induced RGCs.
The expression of RIPK1/RIPK3 in RGCs increased after D-glucose treatment.
Necrostatin-1 suppressed RIPK1/RIPK3 overexpression in RGCs under high glucose induction.
Acknowledgements
This study was supported by Sichuan Provincial Science, Technology Support Project (No. 2018FZ0031).
Disclosure statement
No potential conflict of interest was reported by the author(s).