

ABSTRACT
Osteosarcoma (OS) is the most common primary malignant tumor in children and adolescents. Numerous studies have reported the importance of miRNA in OS. The purpose of this study is to predict potential biomarkers and new therapeutic targets for OS diagnosis and prognosis by analyzing miRNAs of OS plasma samples from the Gene Expression Omnibus (GEO) database.
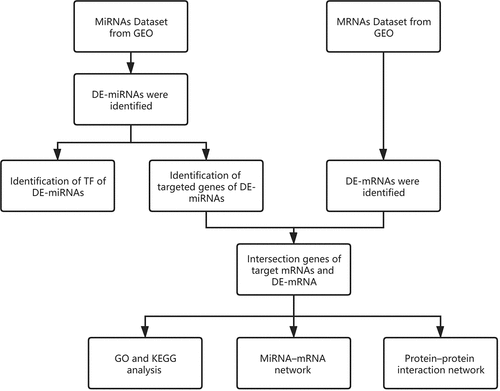
Data-sets were downloaded from the GEO and analyzed using R software. Different expressions of miRNAs (DE-miRNAs) in plasma and mRNAs (DE-mRNAs) in OS patients were identified. Funrich was used to predict the transcription factors and target genes of miRNAs. By comparing the target mRNAs and DE-mRNAs, the intersection mRNAs were identified. The intersection mRNAs were imported to perform Gene Ontology (GO) functional annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. MiRNA-mRNA regulatory network and a protein-protein interaction (PPI) network were constructed by using Cytoscape. Finally, a total of 164 DE-miRNAs, 256 DE-mRNAs, and 76 intersection mRNAs were identified. The top 10 TF of up- and down-regulated DE-miRNAs were also predicted. In addition, GO and KEGG analyses further revealed the intersection mRNAs. By constructing the miRNA–mRNA networks, we found miR-30d-5p, miR-17-5p, miR-98-5p, miR-301a-3p, and miR-30e-5p were the central hubs. COL1A1, COL1A2, MMP2, CDH11, COL4A1 etc. were predicted to be the key mRNA by constructing the PPI networks. Through a comprehensive bioinformatics analysis of miRNAs and mRNAs in OS, we explored the potential effective biomarkers and novel therapeutic targets for the diagnosis and prognosis of OS.
Highlights
We predicted underlying TF and target genes of DE-miRNAs from OS plasma samples.
Based on the blood samples of OS patients, GO functional annotation and KEGG pathway enrichment analysis were performed.
Based on the blood samples of OS patients, MiRNA-mRNA network, and PPI network were constructed.
Availability of data and material
All data are fully available without restriction.
Disclosure statement
The authors declare that they have no competing interests.
Authors’ contributions
WJ and XK conceived of the design of the study. LY, ZP, and ZJ participated in the design of the study and performed the statistical analysis. XK finished the manuscript. All authors read and approved the final manuscript.