

ABSTRACT
Metabolism affects the development, progression, and prognosis of various cancers, including breast cancer (BC). Our aim was to develop a metabolism-related long non-coding RNA (lncRNA) signature to assess the prognosis of BC patients in order to optimize treatment. Metabolism-related genes between breast tumors and normal tissues were screened out, and Pearson correlation analysis was used to investigate metabolism-related lncRNAs. In total, five metabolism-related lncRNAs were enrolled to establish prognostic signatures. Kaplan-Meier plots and the receiver operating characteristic (ROC) curves demonstrated good performance in both training and validation groups. Further analysis demonstrated that the signature was an independent prognostic factor for BC. A nomogram incorporating risk score and tumor stage was then constructed to evaluate the 3 – and 5-year recurrence-free survival (RFS) in patients with BC. In conclusion, this study identified a metabolism-related lncRNA signature that can predict RFS of BC patients and established a prognostic nomogram that helps guide the individualized treatment of patients at different risks.
GRAPHICAL ABSTRACT
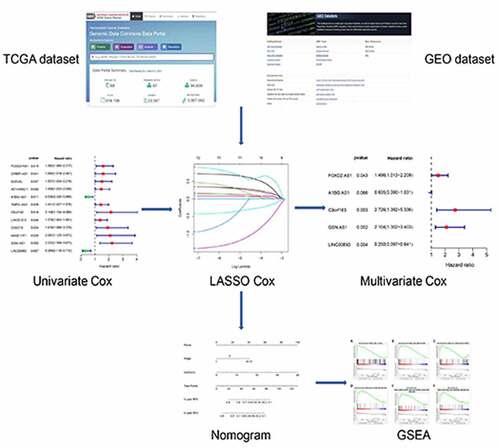
Acknowledgements
We thank all the participants who were involved in this study and gratefully acknowledge contributions from TCGA and GEO Network.
Abbreviations
BC, breast cancer; lncRNA, long non-coding RNA; RFS, recurrence-free survival; TCGA, The Cancer Genome Atlas; GEO, Gene Expression Omnibus; KEGG, Kyoto Encyclopedia of Genes and Genomes; GSEA, gene set enrichment analysis; ROC, receiver operating characteristic; PCA, principal component analysis; MSigDB, Molecular Signatures Database; LASSO, least absolute shrinkage and selection operator; TNM, tumor-node-metastasis; HCC, hepatocellular carcinoma.
Availability of data and material
The datasets analyzed in this study were obtained from the TCGA database (http://www.cancer.gov/tcga) and GEO (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE21653).
Disclosure statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Author contributions
JYM and QL designed the study. JYM, SHL, and JC performed the experiments. JYM, SHL, JC, and QL analyzed the data. All authors contributed to writing the manuscript. All authors contributed to editorial changes involving the manuscript. All authors have read and approved the final manuscript.
Highlights
Crosstalk between lncRNAs and metabolism has been confirmed in several types of cancer.
A novel, robust and reliable metabolism-related lncRNA signature was established for recurrence of breast cancer.
We also developed a prognostic nomogram using a combination of risk scores and clinicopathological variables.
Our nomogram describes a novel quantitative signature for the risk of breast cancer recurrence.
Supplementary material
Supplemental data for this article can be accessed here