

ABSTRACT
Cardiac hypertrophy can cause heart failure. However, the mechanisms underlying the progression of cardiac hypertrophy remain unclear. Emerging evidence suggests that circular RNAs (circRNAs) play a critical role in cardiac hypertrophy. However, the association between circ_nuclear factor I X (circNfix) and cardiac hypertrophy remain largely unknown. Therefore, the aim of the present study was to explore the role of circNfix in cardiac hypertrophy. In order to detect the function of circNfix in cardiac hypertrophy, cardiomyocytes were stimulated with angiotensin II (Ang II) to mimic the pathogenesis of the disease. In addition, pressure overload-induced cardiac hypertrophy in a mouse model was established using transverse aortic constriction (TAC) surgery. The mechanism via which circNfix regulated cardiac hypertrophy was investigated using RNA pull-down and luciferase reporter assays, and fluorescence in situ hybridization (FISH). circNfix was downregulated in Ang II-treated cardiomyocytes. Similarly, circNfix expression was markedly downregulated in mice following TAC surgery. In addition, circNfix overexpression significantly prevented the progression of cardiac hypertrophy in TAC-treated mice. Luciferase activity and RNA pull-down assays indicated that circNfix could indirectly target activating transcription factor 3 (ATF3) by binding with microRNA (miR)-145-5p in cardiomyocytes. miR-145-5p overexpression or ATF3 knockdown could reverse the effects of circNfix in Ang II-treated mouse cardiomyocytes. circNfix attenuated pressure overload-induced cardiac hypertrophy by regulating the miR-145-5p/ATF3 axis. Therefore, circNfix may serve as a molecular target for cardiac hypertrophy treatment.
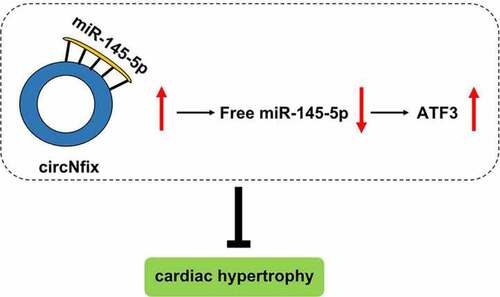
GRAPHICAL ABSTRACT
Highlights
CircNfix was downregulated in cardiac hypertrophy.
CircNfix could attenuate pressure overload-induced cardiac hypertrophy.
CircNfix/miR-145-5p/ATF3 axis regulates cardiac hypertrophy progression.
CircNfix might serve as a potential target for prevention of cardiac hypertrophy.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability
All the data are available from the corresponding author due to reasonable request.