

ABSTRACT
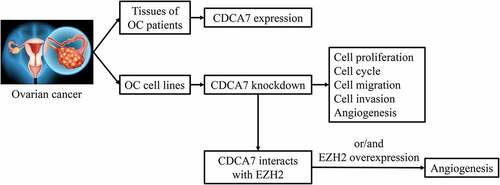
The purpose of the current study was to investigate the biological function of cell division cycle-associated protein 7 (CDCA7) on ovarian cancer (OC) progression and analyze the molecular mechanism of CDCA7 on OC cellular processes and angiogenesis. CDCA7 expression in OC tissues and adjacent normal tissues was obtained from Gene Expression Profiling Interactive Analysis (GEPIA) and in various cancer cell lines was obtained from Cancer Cell Line Encyclopedia (CCLE). Moreover, CDCA7 expression in adjacent normal tissues and tumor tissues of OC patients as well as in normal ovarian epithelial cells (NOEC) and ovarian cancer cells (OVCAR3, SKOV3, CAOV-3, A2780) was further confirmed via Western blot assay and Reverse transcription-quantitative polymerase chain reaction (RT-qPCR). In addition, Immunohistochemistry (IHC) was also applied for determination of CDCA7 expression in tissues of OC patients. Then, SKOV3 cells were introduced with shRNA-CDCA7 for functional experiments. GeneMANIA database analysis and coimmunoprecipitation (Co-IP) assay verified the interaction between CDCA7 and enhancer of zeste homolog 2 (EZH2) to probe the potential mechanism. CDCA7 expression was elevated in tumor tissues of OC patients and OC cell lines. CDCA7 silencing restrained the proliferative, migrative and invasive capacities and arrested cell cycle of OC cells. In addition, CDCA7 knockdown induced a weaker in vitro angiogenesis of HUVECs. Mechanistically, CDCA7 interacted with EZH2. Downregulation of CDCA7 arrested angiogenesis by suppressing EZH2 expression. To sum up, the current study revealed the impact and potential mechanism of CDCA7 on OC cellular processes, developing a promising molecular target for OC therapies.
Graphical Abstract
Disclosure statement
No potential conflict of interest was reported by the author(s).
Availability of data and materials
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
Ethic approval
This study was approved by the Ethics Committee of the Affiliated Huaian No.1 People’s Hospital of Nanjing Medical University.