

ABSTRACT
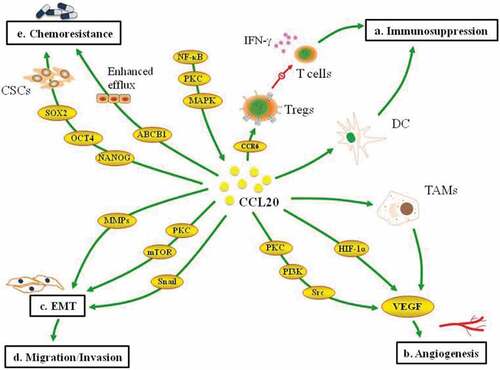
Emerging studies have demonstrated notable roles of CCL20 in breast cancer progression. Based on these findings, CCL20 has become a potential therapeutic target for cancer immunotherapy. Accordingly, studies utilizing monoclonal antibodies to target CCL20 are currently being experimented. However, the existence of cytokine network in the tumor microenvironment collectively regulates tumor progression. Hence, a deeper understanding of the role of CCL20 and the underlying signaling pathways regulating the functions of CCL20 may provide a novel strategy for therapeutic interventions. This review provides the current knowledge on how CCL20 interacts with breast cancer cells to influence tumor progression via immunosuppression, angiogenesis, epithelial to mesenchymal transition, migration/invasion and chemoresistance. As a possible candidate biomarker, we also reviewed signal pathways and other factors in the tumor microenvironment regulating the tumor-promoting functions of CCL20.
These new insights may be useful to design new potent and selective CCL20 inhibitors against breast cancer in the future.
Graphical Abstract
Acknowledgements
We acknowledge the funding support from The Academic and Technological Leaders and Reserves Foundation of Anhui Province (2019H230) and China Scholarship Council (2020-1417).
Article Highlights
The roles of CCL20 in breast cancer progression have been summarized.
Communication between CCL20 and other factors in breast cancer tumor microenvironment have been reported.
Targeting CCL20 can be a useful strategy for breast cancer treatment.
Author Contributions
Louis BK drafted and revised the manuscript; SJ Wang and Youjing Sheng revised the manuscript; Q Wu provided overall research supervision, revised the manuscript and funded the work.
Disclosure statement
Authors declare there is no competing interest.