

ABSTRACT
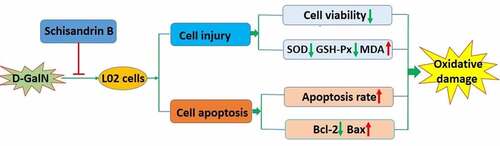
Schisandrin B is a dibenzocyclooctadiene derivative extracted fromSchisandra chinensis (Turcz.) Baill., that exhibits anti-oxidation, anti-inflammation, anti-tumor and hepatoprotective activities. To understand the hepatoprotective mechanism of schisandrin B, this study investigated the efficacy of schisandrin B on L02 cells after treatment with D-GalN. Following pretreatment with 40 μM schisandrin B, L02 cells were stimulated with 40 mM D-GalN. Cell viability, apoptosis, the expression levels of genes associated with apoptosis, and the intracellular oxidative stress indexes were measured. The viability of L02 cells was determined using MTT assay, and the Annexin V-FITC/PI assay kit was utilized for the assessment of apoptosis. The activities of GSH-Px and SOD, the level of MDA were assessed, separately, using relative detection kits. Moreover, RT-PCR as well as Western blot was applied to measure the mRNA and protein expression of Bax and Bcl-2. The results indicated that schisandrin B significantly prevented D-GalN‑induced oxidative damage in L02 cells (P<0.05), decreased GSH-Px and SOD activities (P<0.05), increased MDA content (P<0.05). Furthermore, schisandrin B inhibited D-GalN-induced apoptosis in L02 cells (P<0.05), regulated the expression of Bax and Bcl-2 (P<0.05). The results indicated that schisandrin B decreased the D-GalN-induced intracellular oxidative stress indexes generation, and inhibited the down-regulation of Bcl-2 and up-regulation of Bax induced by D-GalN. In conclusion, schisandrin B was shown to exert protective effect against oxidative damage of L02 cells, which, in part, was achieved by regulating the mRNA and protein levels of Bax and Bcl-2.
Graphical Abstract
Abbreviations
L02 cells normal human hepatocytes; D-GalN D-galactosamine; EDTA ethylenediamine tetraacetic acid; FBS fetal bovine serum; DMEM dulbecco’s modified eagle medium; P/S penicillin/streptomycin; DMSO dimethyl sulfoxide; MTT 3-(4, 5-dmethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide; MDA malondialdehyde; GSH-Px glutathione peroxidase; SOD superoxide dismutase; BCA bicinchoninic acid; SDS-PAGE sodium dodecyl sulfate-polyacrylamide gel electrophoresis; PVDF polyvinylidene fluoride; TBST tris-buffered saline containing 0.1% Tween-20; GAPDH glyceraldehyde-3-phosphate dehydrogenase; PCR polymerase chain reaction; mRNA messenger RNA; ANOVA one-way analysis of variance; Bcl-2 an anti-apoptotic protein; Bax an apoptotic protein.
Highlights
Schisandrin B inhibited D-GalN-induced apoptosis in L02 cells.
Schisandrin B prevent D-GalN-induced oxidative in L02 cells.
The regulatory effects of schisandrin B on Bax and Bcl-2 in D-GalN-induced L02 cells.
Disclosure statement
No potential conflict of interest was reported by the author(s).