
ABSTRACT
Warburg effect is considered to be related to the malignancy of tumor cells under hypoxic conditions, but the underlying mechanism remains unknown. In this article, it has been reported that lncRNA LINC00525 is a hypoxia-responsive lncRNA and is essential for hypoxia-enhanced glycolysis. It was found that LINC00525 was up-regulated, and promoted cell proliferation in colorectal cancer in vitro and in vivo. In colorectal cancer cells, hypoxia increasedLINC00525 expression, whereas knocking down LINC00525 reduced hypoxia-enhanced glycolysis. For specific molecular mechanisms, it was found that LINC00525 promoted UBE2Q1 expression by binding miR-338-3p, and UBE2Q1-stabilized β-catenin enhances hypoxia-enhanced glycolysis by activating HIF-1α. In conclusion, these findings showed that LINC00525 was essential for hypoxia-enhanced glycolysis; its mechanism was related to activating HIF-1α through miR-338-3p/UBE2Q1/β-catenin axis in colorectal cancer cells.
1 Introduction
Cancer cells can escape the normal apoptosis pathway by exhibiting a unique metabolic phenotype that enhances the proliferation and migration ability. This unique metabolic phenotype refers to the Warburg effect, and it plays a significant role in the pathogenesis of tumors [Citation1]. The Warburg effect occurs when cancer cells weaken aerobic respiration through a series of molecular mechanisms under aerobic conditions, carry out efficient glycolysis, and then obtain a large amount of adenosine triphosphate, and create a microenvironment suitable for tumor cells to survive, proliferation and movement [Citation2,Citation3]. In addition, the Warburg effect inhibits the monitoring and lethality of T lymphocytes through local hypoxia, creating tumor immune escape [Citation4]. In conclusion, the Warburg effect is a critical factor in tumor cell malignancy, but its exact molecular mechanism is unclear, and adaptive responses to the unique tumor hypoxic microenvironment may represent a well-known mechanism [Citation5].
Long non-coding RNA (lncRNA) is a non-coding RNA with a length of more than 200nt, and was regarded as a ‘transcription byproduct’ or ‘junk gene’ in the long past years because it did not encode a protein [Citation6]. With the advancement of gene sequencing technology in recent years, an increasing number of lncRNAs have been identified and identified as being related to the occurrence and progression of malignant tumors [Citation6,Citation7]. LncRNA LINC0052 is a newly discovered lncRNA that has been found to function as oncogenes in a variety of malignant tumors, including glioma [Citation8], chordoma [Citation9], non-small cell lung cancer [Citation10], and colorectal cancer [Citation11]. Importantly, previous studies reported that LINC00525 is highly expressed in colorectal cancer tissues, and is related to the poor prognosis of colorectal cancer patients [Citation12], and promotes the proliferation of colorectal cancer cells, and inhibits apoptosis [Citation11,Citation13].
miR-338-3p has been found to function as a tumor suppressor gene in colorectal cancer [Citation14,Citation15], and can be used as a downstream target gene for a variety of lncRNAs to function as oncogenes to regulate biological characteristics of colorectal cancer cells [Citation16,Citation17], including LINC0052510. Moreover, miR-338-3p has also been found to be involved in the regulation of the Warburg effect of tumor cells [Citation18], and as a downstream gene of lncRNA to promote glycolysis in tumor cells [Citation19]. Ubiquitin-conjugating enzyme E2Q family member 1 (UBE2Q1), a target gene of miR-338-3p [Citation20], has been found to stabilize β-catenin in hepatocellular carcinoma [Citation21]. Due to β-catenin stabilized HIF-1α by interacting with HIF-1α under hypoxic condition [Citation22] and HIF-1α was a key mediator of hypoxic response, UBE2Q1 might stabilize HIF-1α by stabilizing β-catenin. In this study, it was hypothesized that LINC00525 regulated the expression of UBE2Q1 is activated by miR-338-3p via the bridging effect, whereas UBE2Q1 activated HIF-1 via stabilizing-catenin, ultimately promoting hypoxia-enhanced glycolysis in colorectal cancer. The aim of the present study was to discover a new mechanism for LINC00525 to regulate hypoxic glycolysis in colorectal cancer cells. The goal of this study was to demonstrate that LINC00525 activated HIF-1α through miR-338-3p/UBE2Q1/β-catenin axis to regulate the Warburg effect in colorectal cancer.
2 Materials and methods
2.1 Cell culture and cell
Human normal colonic epithelial cells (NCM-460) and human colorectal cancer cells (SW480, HCT116, HT29, SW620, SW1116, and LoVo) were all cultured in McCoy’s 5A medium (12,330,031, Gbico) supplemented with 10% fetal bovine serum (16,140,071, Gbico) in 37°C with 5% CO2.
2.2 qPCR analysis
Cells were collected to extract total RNA using an RNA extraction Kit (RC101-01, Vazyme, China), and cDNA was produced as directed using a PrimeScript RT reagent (RR047A, Taraka, Japan) [Citation23,Citation24]. Then, 20 μL of the qPCR system was prepared and analyzed as described in the instructions of GoTaq qPCR Master Mix (A6001, Promega, USA) in a real-time quantitative PCR instrument(Accurate96-x4, Dlab). The relative expression of the gene was calculated by the 2−ΔΔCt method. β-actin was loaded as a control for lncRNA and mRNA, and U6 was loaded as a control for miRNA. Primers for qPCR are shown in .
Table 1. Sequences of primers of qPCR, si-RNA and miRNA
2.3 Cell transfection and adenovirus infection
To knock down the expression of LINC00525 and β-catenin, we transfected small interfering RNA into colorectal cancer cells as previously described [Citation25,Citation26]. In brief, 50 nmol/l of special small interfering RNA for LINC00525 (si-525) or β-catenin (si-β-catenin) were directly transfected into 2.5 × 106 SW620, SW1116 and LoVo cells using Lipofectamine 2000 according to the manufacturer’s protocols. After 24 hours of transfection, experiments were performed. Similarly, 50 nmol/l of miR-338-NC (NC), miR-338-mimic (mimic), and miR-338-inhibitor (inhibitor) were directly transfected into 2.5 × 106 SW620, SW1116 and LoVo cells using Lipofectamine 2000 according to the manufacturer’s protocols. To test the target binding between miR-338-3p and LINC00525/UBE2Q1, we first cloned wild-type (WT) or mutated (MUT) versions of the 3ʹ-UTR of LINC00525 or UBE2Q1 into pisCHECK2 (Addgene, 97,157) to build a recombinant plasmid. Next, LoVo cells expressing NC, mimic, or inhibitor was transfected with recombinant plasmid. To overexpress UBE2Q1 expression, 30 μl adenovirus for overexpressing UBE2Q1 (AAV-UBE2Q1) (Vector Biolabs, AAV-MI0012528) were directly added into the cell culture medium, and empty adenovirus (AAV-NC) was used as control.
2.4 FISH staining analysis
1 x 105 cells were seeded into Lab-Tek cell culture well (155,411, Thermo Scientific). The cells were fixed with cold acetone 24 hours later and then incubated with a fluorescent probe for binding to the human version of the LINC00525 gene, which was synthesized by Genomeditech Co., Ltd. For cells, it should be counterstained the nucleus with 5 μg/mL DAPI for 5 minutes at room temperature. At last, all samples were analyzed by confocal microscopy (TCS SP5, Leica).
2.5 Cell proliferation assay
A total of 1 × 104 cells were seeded into 96 cell culture plates and cultured for 72 hours at 37°C with 5% CO2. The cell viability was measured with CCK8 reagent (C0038, Beyotime) at 0, 24, 48, and 72 hours respectively.
For cell clone test: 200 cells with 2 mL medium were seeded into a 6-well cell culture plate and culture for 2 weeks at 37°C with 5% CO2. At last, we removed the cell culture medium and washed them twice with PBS buffer. Added formaldehyde to fix at room temperature for 30 minutes, and then add 0.1% crystal violet for staining at room temperature for 20 minutes and wash 3 with PBS. Finally, the cell clone number was counted.
In vivo cell proliferation experiment [Citation27]: we injected 5 × 106/0.2 ml cells into the nude mouse (6 weeks old, 20–23 g) in middle axillary lateral skin (n = 7 each group). We euthanized nude mice after 2 weeks, isolated xenograft tumors from colorectal cancer nude mice, and weighed them.
2.6 Cellular immunofluorescence
A total of 1 × 105 cells were seeded into Lab-Tek cell culture well ((155,411, Thermo Scientific). After different treatments, we removed the cell culture medium and washed cells twice with PBS buffer. After being fixed and blocked, cells were incubated with GLUT1 antibody (ab115730, Abcam) or UBE2Q1 antibody (bs-24,324 R, Bioss) overnight at 4°C. The next day, we washed the cells 3 times with PBS buffer before incubating them with Alexa Fluor488 goat-anti-rabbit IgG (H + L) (A11008, Invitrogen). For cells, it should be counterstained the nucleus with 5 μg/mL DAPI for 5 minutes at room temperature. At last, all samples were analyzed by confocal microscopy (TCS SP5, Leica).
2.7 Lactate production and glucose uptake assay
After 24 hours of culture under normoxic (21% O2) or hypoxic conditions (1% O2), a cell culture medium was collected to detect the levels of lactate using a PicoProbe™ Lactate Fluorometric Assay Kit (K638, BioVision), and to detect the intracellular glucose levels using a PicoProbe™ Glucose Fluorometric Assay Kit (K688, BioVision).
2.9 Western blot analysis
Cells were harvested to extract total protein a Tissue/cells Protein Extraction Reagent (78,510, ThermoFisher). To determine the protein concentration, a BCA kit(23,227, ThermoFisher, USA) is used. Next, 50 μg total protein is analyzed using a 10% SDS-PAGE. After transferring, PVDF membranes (LC2002, ThermoFisher) are first blocked with 5% skimmed milk powder and then are probed with primary antibodies against UBE2Q1 (bs-24,324 R, Bioss), HIF-1α (ab279654, Abcam), or β-catenin (ab32572, Abcam). Proteins were visualized with ECL solution (WBKLS0100, Beijing Xinjingke Biotechnologies Co., Ltd, China), followed by densitometry analysis using Imag J 3.0 (IBM, USA) and β-actin was loading as control.
2.10 Statistical analysis
Data in this study were analyzed by SPSS 20.0 software. Data shown are mean ±SD. The difference between the two groups was compared by Student's t-test. For multiple groups, one-way ANOVA was used to compare the differences between multiple groups and then use the Turkey test as a post-doc test to compare the differences between the two groups. P-value less than 0.05 indicated a significant difference.
3 Results
In the present study, the main work is about glycolysis in colorectal cancer cells under hypoxia. Our goal is to discover a new mechanism for LINC00525 to regulate the Warburg effect of colorectal cancer cells under hypoxia. Fortunately, we not only found that LINC00525, which was highly expressed in colorectal cancer cells, promoted the proliferation of colorectal cancer cells in vivo and in vitro but also found that LINC00525 activated HIF-1α through the miR-338-3p/UBE2Q1/β-catenin axis to enhabcehypoxia-enhanced glycolysis in colorectal cancer.
3.1 LINC00525 is up-regulated and promotes cell proliferation in colorectal cancer
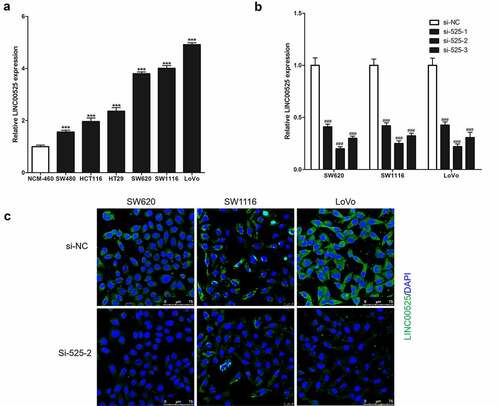
As a result of analysis in GSE110715, GSE32323, and GSE21510 datasets [Citation12], LINC00525 was highly expressed in colorectal cancer tissues. In this study, it was found that the expression of LINC00525 in human colorectal cancer cells (SW480, HCT116, HT29, SW620, SW1116, and LoVo) were all markedly higher than its expression in human normal colonic epithelial cells (NCM-460) ()). To investigate the function of LINC00525, the expression of LINC00525 was knocked down in human colorectal cancer cells (SW620, SW1116, and LoVo) by transferring into LINC00525-specific small interfering RNA (si-525-1, si-525-2, and si-525-3), and meaningless small interfering RNA (si-NC) was used as a negative control. As shown in , si-525-1, si-525-2, and si-525-3 all strongly reduced the expression of LINC00525 in colorectal cancer cells, with si-525-2 having the highest knockdown effect of the three siRNAs, decreasing LINC00525 expression to less than 25%. Similarly, the results of FISH staining also showed that si-525-1, si-525-2, and si-525-3 were all strongly knockdown the expression of LINC00525 in colorectal cancer cells, and si-525-2 has the best knockdown effect ()). Therefore, if there were no special instructions, LINC00525-specific small interfering RNA (si-525) was si-525-2 in the following research. In addition, FISH staining analysis also showed that LINC00525 was mainly cytoplasmic localization in colorectal cancer cells ()).
Figure 1. Expression of LINC00525 in colorectal cancer cells.
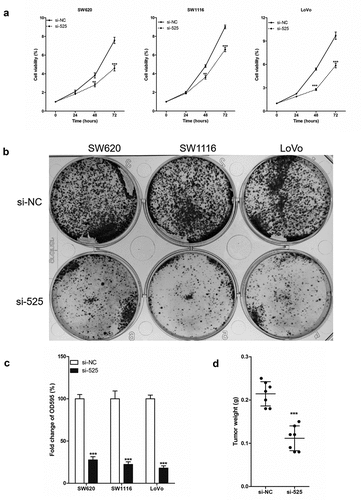
Simultaneously, the effect of LINC00525 expression on the proliferation of colorectal cancer cells was explored with revealed that as compared to the colorectal cancer cells of the si-NC group, the cell viability of the si-525 group was significantly reduced in the 3-day culture experiment ()). Similarly, the results of the cell clone formation experiment also showed that the number of colorectal cancer cell clones in the si-525 group was significantly were significantly less than that in the si-NC group (). In vivo, the weights of LoVo xenografts harvested 3 weeks after injection in the si-525 group were significantly lower than that in the si-NC group (). In conclusion, LINC00525 was highly expressed in colorectal cancer cells, and LINC00525 knockdown reduced the proliferation of colorectal cancer cells.
Figure 2. LINC00525 promoted the proliferation of colorectal cancer cells in vitro and in vivo.
3.2 LINC00525 is essential for hypoxia-enhanced glycolysis
One of the features of tumor cells is their dependence on glycolytic energy supply. To study whether LINC00525 regulates the Warburg effect under hypoxic conditions, we first assessed the amount of LINC00525 in colorectal cancer cells (SW620, SW1116, and LoVo) using RT-qPCR analysis. As shown in ), the expression of LINC00525 in colorectal cancer cells under hypoxic conditions (1% O2 for 24 hours) was significantly higher than that in colorectal cancer cells under normoxic conditions (21% O2 for 24 hours). Moreover, we also study the effect of different hypoxia conditions on the expression of LINC00525 in colorectal cancer cells (LoVo) and found that LINC00525 expression increased as the oxygen concentration decreased ()), and also increased as with the duration of hypoxia ()). At the same time, the expression of HIF-1α protein was also detected which showed that its expression was also related to hypoxia in a concentration-dependent and time-dependent manner.
Figure 3. LINC00525 promoted hypoxic glycolysis in colorectal cancer cells.
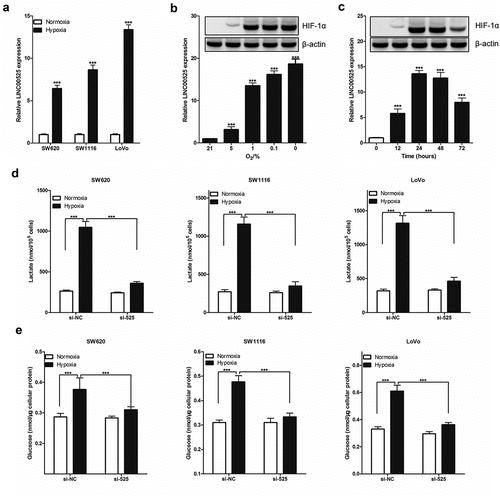
To investigate the functional role of hypoxia-induced LINC00525, we knocked down the expression of LINC00525 in colorectal cancer cells (SW620, SW1116, and LoVo) by transferring into si-525. It has been well recognized that hypoxia causes increased glycolysis. Treatment with hypoxia resulted in a significant increase in lactate production ()) and glucose uptake ()). All these hypoxia-induced effects, however, were greatly reversed by LINC00525 knockdown (), indicating that LINC00525 functions as a critical mediator of hypoxia-enhanced glycolysis.
3.3 HIF-1α is necessary for the effect of LINC00525 on hypoxia-enhanced glycolysis
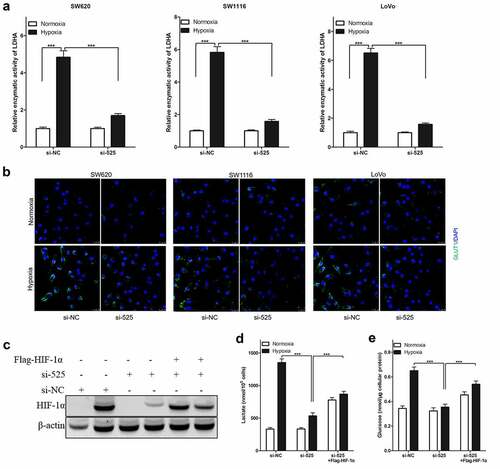
In colorectal cancer cells, LINC00525 knockdown significantly reduced hypoxia-enhanced lactate production ()) and glucose uptake ()) (SW620, SW1116, and LoVo), similar to the above findings. It was also found that LINC00525 knockdown significantly decreased the relative enzymatic activity of LDHA ()) and GLUT1 protein expression ()) in colorectal cancer cells (SW620, SW1116, and LoVo) after being cultured under hypoxia. Due to HIF-1α being necessary for hypoxia-enhanced LDHA activity and GLUT1 expression, whether LINC00525 promoted hypoxic-enhanced glycolysis by regulating HIF-1α was investigated. Firstly, it was found that LINC00525 knockdown decreased the expression of HIF-1α in LoVo cells after being cultured under hypoxia ()). Therefore, to study whether HIF-1α is necessary for the effect of LINC00525 on hypoxia-enhanced glycolysis, exogenous HIF-1α was introduced into LINC00525 knockdown LoVo cells, which upregulated the expression of HIF-1α protein. The results of immunoblotting showed that Flag-HIF-1α increased the expression of HIF-1α protein in LINC00525 knockdown LoVo cells ()). Next, the glycolysis in LoVo cells was assessed. Overexpression of HIF-1a indeed reversed the effect of LINC00525 on lactate production ()) and glucose uptake ()). Therefore, our data indicated that the effect of LINC00525 on hypoxic glycolysis was HIF-1α dependent.
Figure 4. Effect of LINC00525 on hypoxic glycolysis is HIF-1α dependent.
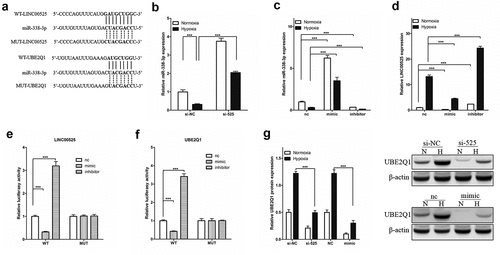
3.4 LINC00525 promotes UBE2Q1 expression by binding miR-338-3p
The public databases (UCSC, miRbase, and BiBiserv2) were used to gain insight into the specific molecular mechanism of LINC00525 in hypoxic glycolysis. Interestingly, it was found that miR-338-3p, a Warburg effect-related miRNA [Citation18] and a tumor suppressor gene in colorectal cancer cells [Citation14,Citation28]. Interestingly, we found that miR-338-3p, LINC00525, and UBE2Q1 all have mutually binding sequences ()). As shown in ), LINC00525 knockdown significantly increased the expression of miR-338-3p in LoVo cells under normoxic and hypoxic conditions (). Next, miR-338-3p expression was enhanced by transferring into miR-338-3p-mimic (mimic) and knockdown miR-338-3p expression by transferring into miR-338-3p-inhibitor (inhibitor) in LoVo cells and confirmed by qPCR ()). Importantly, it was found that under normoxic and hypoxic conditions, mimic significantly decreased LINC00525 expression whereas inhibitor significantly enhanced LINC00525 expression in LoVo cells ()), which suggests LINC00525 and miR-338-3p reduce the expression of each other by binding. Consistent with the above findings that miR-338-3p inhibited LINC00525 expression in LoVo cells, the dual fluorescent gene reporting system showed the luciferase activity of LINC00525-wt was significantly decreased by mimic, while miR-338-3p had no influence on LINC00525-Mut in LoVo cells, suggesting miR-338-3p directly inhibited LINC00525 expression ()). At the same time, the results of the dual fluorescent gene reporting system also showed that miR-338-3p ()). Following immunoblotting results, LINC00525 knockdown significantly decreased UBE2Q1 protein expression, while miR-338-3p overexpression also significantly decreased UBE2Q1 protein expression in LoVo cells under normoxic and hypoxic conditions ()).
Figure 5. LINC00525 promoted UBE2Q1 expression by sponging to miR-338-3p.
3.5 LINC00525 promotes hypoxia-enhanced glycolysis is UBE2Q1 dependent
UBE2Q1, whose full name is ubiquitin-conjugating enzyme E2Q family member 1, was found to be highly expressed in colorectal cancer tumor tissues and cells [Citation29]. To investigate the effect of UBE2Q1 on hypoxia-enhanced glycolysis, we infected LoVo cells with AAV-UBE2Q1 adenovirus and found that immunoblotting and immunofluorescence significantly increased the expression of UBE2Q1 in LoVo cells ()). LINC00525 knockdown LoVo cells were infected with AAV-UBE2Q1 to upregulated UBE2Q1 protein expression.
Figure 6. Effect of LINC00525 on hypoxic glycolysis is UBE2Q1 dependent.
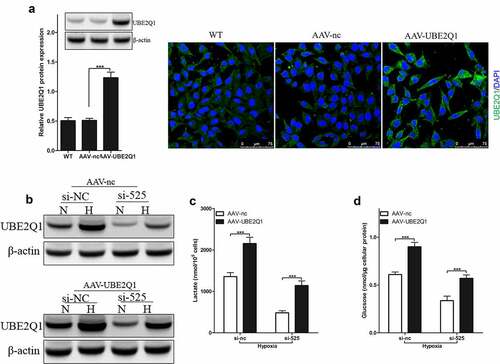
Results of immunoblotting showed that AAV-UBE2Q1 increased the expression of UBE2Q1 protein in LINC00525 knockdown LoVo cells ()). Then, glycolysis in LoVo cells was checked. Overexpression of UBE2Q1 indeed reversed the effect of LINC00525 on lactate production ()) and glucose uptake ()). Therefore, these data indicated that the effect of LINC00525 on hypoxic glycolysis was UBE2Q1 dependent.
3.6 UBE2Q1enhanced hypoxia-enhanced glycolysis by stabilizing β-catenin
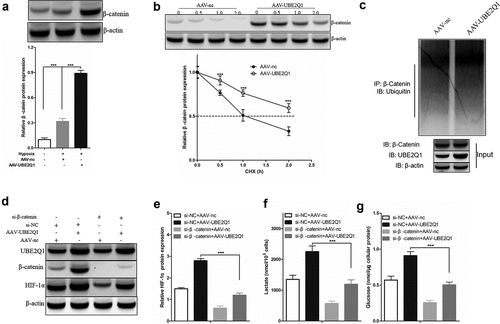
Overexpression of UBE2Q1 reversed the effect of LINC00525 on hypoxia-enhanced glycolysis, as seen above. Overexpression of UBE2Q1 specifically enhanced hypoxia-enhanced glycolysis. However, so far, its molecular mechanism is still unknown. In a previous study, UBE2Q1 has found to stabilize β-catenin in hepatocellular carcinoma [Citation21], and β-catenin interacts with HIF-1α under hypoxic conditions [Citation22]. These clues hint that UBE2Q1-stabilized β-catenin enhances hypoxia-enhanced glycolysis by activating HIF-1α. To test this hypothesis, we first found that overexpression of UBE2Q1 significantly β-catenin protein expression in LoVo cells under hypoxia ()). In addition, cycloheximide (CHX) tracking experiments showed that the half-life of β-Catenin was prolonged in LoVo cells overexpressing UBE2Q1 (Figure 7BAt the same time, we observed that overexpression of UBE2Q1 significantly enhanced-catenin ubiquitination in LoVo cells ()). To study the effect of β-catenin on the effect of UBE2Q1 on hypoxia-enhanced glycolysis, we used si-β-catenin to knock down the expression of β-catenin in UBE2Q1 overexpression LoVo cells ()). Importantly, it was found that β-catenin knockdown reversed the effect of UBE2Q1 on HIF-1α activation ()), lactate production ()), and glucose uptake ()). Therefore, these results indicated that UBE2Q1-stabilized β-catenin enhances hypoxia-enhanced glycolysis by activating HIF-1α.
Figure 7. UBE2Q1 stabilized β-catenin, and β-catenin knockdown hypoxia-induced abrogated LINC00525-augmented HIF-1α activation and glycolysis.
4. Discussion
In this study, it was found that LINC00525 was highly expressed and promoted cell proliferation in colorectal cancer, which has been found in previous studies [Citation11,Citation13]. This study focused on the Warburg effect as it is a key factor in tumor pathogenesis 1 and provides a microenvironment suitable for tumor cells to survive, proliferate, and move [Citation2,Citation3]. More importantly is that there is no research between LINC00525 and the Warburg effect of colorectal cancer cells, which guarantees the innovation of research.
In the present study, it was found for the first time that LINC00525 knockdown decreased hypoxia-enhanced glycolysis, and overexpression of HIF-1α reversed the effect of LINC00525 knockdown on hypoxia-enhanced glycolysis in colorectal cancer, which suggests that LINC00525 promoted hypoxia-enhanced glycolysis, and this effect was HIF-1α dependent. Previous studies have shown that under hypoxic conditions, the activation of HIF-1α contributes to the Warburg effect by synergistically up-regulating glycolysis and down-regulating oxidative phosphorylation. It followed this mechanism: increase the uptake of glucose by cancer cells by up-regulating the expression of GLUT1 [Citation30,Citation31], and speed up the metabolism from glucose to pyruvate, and promote the conversion of pyruvate to lactic acid by up-regulating the expression of LDHA [Citation32,Citation33].
Furthermore, we found that LINC00525 is mostly present in the cytoplasm of colorectal cancer cells. The localization of lncRNA in cells is well known to be closely related to its mechanism of action. The ceRNA regulation mechanism is one of the important ways for lncRNA to regulate post-transcription in the cytoplasm. Previous studies have found that lncRNA can be used as the ‘sponge’ of miRNA, competitively binding to miRNA, thereby affecting the regulation of miRNA on downstream target genes [Citation6,Citation32]. Therefore, the core of the cytoplasm transcriptional regulation mechanism of lncRNA is that lncRNA and a specific mRNA share a miRNA. miR-338-3p was identified as a downstream gene of LINC00525 through literature search and public databases to study the molecular mechanism of LINC00525 regulating hypoxic glycolysis in colorectal cancer cells. miR-338-3p was identified as a downstream gene of LINC00525 to study hypoxic glycolysis because miR-338-3p was found to inhibit the proliferation and migration of colorectal cancer cells [Citation14,Citation15], as well as it was observed to be involved in the regulation of tumor cell glycolysis 1819. Herein, it was found that LINC00525 and miR-338-3p reduced the expression of each other by binding, and miR-338-3p overexpression and LINC00525 knockdown inhibited UBE2Q1 protein expression in colorectal cancer cells.
UBE2Q1 is found on the long arm of chromosome 1 and encodes the ubiquitinated ligase E2 protein. UBE2Q1 moves activated ubiquitin from the ubiquitin-activating enzyme (E1) to the ubiquitin ligase (E3) and the substrate, activating the substrate’s role to degrade ubiquitin 33. The previous study has found that UBE2Q1 was highly expressed in colorectal cancer [Citation29], and increased the levels of β-catenin protein by stabilizing β-catenin in cancer cells [Citation21], and β-catenin interacts with HIF-1α under hypoxic condition [Citation22]. Consistently, it was found that UBE2Q1 overexpression increased the expression of β-catenin and also increased the ubiquitination level of β-catenin and elongated its half-life in colorectal cancer cells. Moreover, it was found that β-Catenin knockdown removed the enhanced expression of HIF-1α by UBE2Q1 overexpression and reversed the effect of UBE2Q1 on lactate production and glucose uptake in colorectal cancer under hypoxia, suggesting that UBE2Q1-stabilized β-catenin enhances hypoxia-enhanced glycolysis by activating HIF-1α.
Conclusion
It can be concluded that LINC00525 promoted UBE2Q1 expression by binding miR-338-3p, and UBE2Q1-stabilized β-catenin enhances hypoxia-enhanced glycolysis by activating HIF-1α. Understanding how LINC00525 participates in the regulation of the Warburg effect in colorectal cancer provides novel insights on the issue.
Data Availability
The data used to support the findings of this study are available from the corresponding author upon request.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Heiden M, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009;324:1029–1033.
- Liberti MV, Locasale JW. The Warburg effect: how does it benefit cancer cells? Trends Biochem Sci. 2016;41:211–218.
- Icard P, Shulman S, Farhat D, et al. How the Warburg effect supports aggressiveness and drug resistance of cancer cells? Drug Resist Updat. 2018;38:1–11.
- Kornberg MD. The immunologic Warburg effect: evidence and therapeutic opportunities in autoimmunity. Wiley Interdiscip Rev Syst Biol Med. 2020;12:e1486.
- Hayashi Y, Yokota A, Harada H, et al. Hypoxia/pseudohypoxia‐mediated activation of hypoxia‐inducible factor‐1α in cancer. Cancer Sci. 2019;110:1510.
- Gibb EA, Brown CJ, Lam WL. The functional role of long non-coding RNA in human carcinomas. Mol Cancer. 2011;10:38–54.
- Geisler S, Coller J. RNA in unexpected places: long non-coding RNA functions in diverse cellular contexts. Nat Rev Mol Cell Biol. 2013;14:699–712.
- Wan Y, Liang F, Wei M, et al. Long non-coding RNA LINC00525 regulates the proliferation and epithelial to mesenchymal transition of human glioma cells by sponging miR-338-3p. AMB Express. 2020;10:1–9.
- Gong F, Wang X, Sun Q, et al. Long non-coding RNA LINC00525 interacts with miR-31–5p and miR-125a-5p to act as an oncogenic molecule in spinal chordoma. Biochem Biophys Res Commun. 2021;536:80–87.
- Yang Z, Lin X, Zhang P, et al. Long non-coding RNA LINC00525 promotes the non-small cell lung cancer progression by targeting miR-338-3p/IRS2 axis. Biomed Pharmacother. 2020;124:109858.
- Zheng Q, Peng P. Long non-coding RNA Linc00525 is an unfavorable prognostic factor and regulates proliferation and apoptosis in colorectal cancer. Ann Oncol. 2019;30:iv8.
- Ghasemi T, Khalaj-Kondori M, Asadi P. lncRNA-miRNA-mRNA interaction network for colorectal cancer; An in silico analysis. Comput Biol Chem. 2020;89:107370.
- Wang S, Li J, Yang X. Long non-coding RNA LINC00525 promotes the stemness and chemoresistance of colorectal cancer by targeting miR-507/ELK3 axis. Int J Stem Cells. 2019;12:347.
- Teng Z, Jing D, Liang J, et al. miR-338-3p suppresses colorectal cancer proliferation and progression by inhibiting MACC1. Int J Clin Exp Pathol. 2018;11:2256–2267.
- Yong FL, Law CW, Wang CW. Potentiality of a triple microRNA classifier: miR-193a-3p, miR-23a and miR-338-5p for early detection of colorectal cancer. BMC Cancer. 2013;13:1–10.
- Zhang W, Wang B, Wang Q, et al. Lnc-HSD17B11-1:1 functions as a competing endogenous RNA to promote colorectal cancer progression by sponging miR-338-3p to upregulate MACC1. Front Genet. 2020;11:628.
- Li W, He Y, Cheng Z. Long noncoding RNA XIST knockdown suppresses the growth of colorectal cancer cells via regulating microRNA-338-3p/PAX5 axis. Eur J Cancer Prev. 2021;30:132–142.
- Huizhen N, Jun L, Xiao-Mei Y, et al. Mineralocorticoid receptor suppresses cancer progression and the Warburg effect by modulating the miR-338-3p-PKLR axis in hepatocellular carcinoma. Hepatology. 2015;62:1145–1159.
- Liu X, Zhu Q, Guo Y, et al. LncRNA LINC00689 promotes the growth, metastasis and glycolysis of glioma cells by targeting miR-338-3p/PKM2 axis. Biomed Pharmacothe. 2019;117:109069.
- Wang JC, Huang Y, Zhang RX, et al. miR-338–3p inhibits autophagy in a rat model of allergic rhinitis after PM2.5 exposure through AKT/mTOR signaling by targeting UBE2Q1. Biochem Biophys Res Commun. 2021;554:1–6.
- Zhang B, Deng C, Wang L, et al. Upregulation of UBE2Q1 via gene copy number gain in hepatocellular carcinoma promotes cancer progression through β‐catenin‐EGFR‐PI3K‐Akt‐mTOR signaling pathway. Mol Carcinog. 2018;57:201–215.
- Kaidi A, Williams AC, Paraskeva C. Interaction between β-catenin and HIF-1 promotes cellular adaptation to hypoxia. Nat Cell Biol. 2007;9:210–217.
- Zhou X, Wang Z, Xu B, Ji N, Meng P, and Gu Let al, Long non-coding RNA NORAD protects against cerebral ischemia/reperfusion injury induced brain damage, cell apoptosis, oxidative stress and inflammation by regulating miR-30a-5p/YWHAG. Bioengineered 2021; 12:9174–88.
- Chen X, Wang C, Yang P, et al. Ube2s-stabilized β-catenin protects against myocardial ischemia/reperfusion injury by activating HIF-1α signaling. Aging (Albany NY). 2020;12:5716.
- Zhong X, and Cai Y. Long non-coding RNA (lncRNA) HOXD-AS2 promotes glioblastoma cell proliferation, migration and invasion by regulating the miR-3681-5p/MALT1 signaling pathway. Bioengineered 2021; 12:9113–27.
- Xu K, Zhan Y, Yuan Z, et al. Hypoxia induces drug resistance in colorectal cancer through HIF-1α/miR-338-5p/IL6 feedback loop. Mol Ther. 2019;272:1810–1824.
- Shafiee SM, Seghatoleslam A, Nikseresht M, et al. UBE2Q1 expression in human colorectal tumors and cell lines. Mol Biol Rep. 2013;40:7045–7051.
- de Andrade Barreto E, de Souza Santos PT, Bergmann A, et al. Alterations in glucose metabolism proteins responsible for the Warburg effect in esophageal squamous cell carcinoma. Exp Mol Pathol. 2016;101:66–73.
- Ryan HE. HIF-1alpha is required for solid tumor formation and embryonic vascularization. EMBO (European Molecular Biology Organization) Journal. 1998;17:3005–3015.
- Kolev Y, Uetake H, Takagi Y, et al. Lactate Dehydrogenase-5 (LDH-5) expression in human gastric cancer: association with Hypoxia-Inducible Factor (HIF-1α) pathway, angiogenic factors production and poor prognosis. Ann Surg Oncol. 2008;15:2336.
- Palsson-mcdermott EM, Curtis AM, Goel G, et al. Pyruvate kinase M2 regulates Hif-1α activity and IL-1β induction and is a critical determinant of the warburg effect in LPS-activated macrophages. Cell Metab. 2015;21:65–80.
- Marchese FP, Raimondi I, Huarte M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017;18:206.
- Wei J, Yuan Y, Jin C, et al. The ubiquitin ligase TRAF6 negatively regulates the JAK-STAT signaling pathway by binding to STAT3 and mediating its ubiquitination. Plos One. 2012;7:e49567.