
ABSTRACT
Maize has sustained the Zuni and other peoples of the arid American Southwest for more than four millennia. In Zuni dryland agriculture, fields on alluvial fans and other valley-edge landforms are managed to receive supplemental water and nutrients by retaining storm runoff and associated sediment and organic material transported from adjoining uplands. An experiment with Zuni blue maize (Zea mays L.) and commercial hybrid maize was conducted in two Zuni fields to study traditional maize and its productivity as part of a larger study of Zuni agroecosystems. Following a closely related paper on maize growth and productivity, studies of maize nutrient composition and quality are presented here. More nutrient differences occurred between Zuni and hybrid maize than between runoff and other treatments. Zuni grain had higher concentrations of more nutrients than hybrid maize, and for several elements, there was an inverse pattern in grain vs. leaf nutrient concentration between cultivars. Zuni grain had a higher N (protein) concentration than hybrid maize, and a higher proportion of N in grain vs. leaf, indicating an ability to partition more N and other nutrients into grain. Parallel differences in maize and soil N and P were found between the two fields, though the cultivars had different P content patterns. Overall, agronomically sufficient concentrations of most nutrients were present across cultivars and treatments. Long-term production and nutrient management of maize grown without conventional irrigation or fertilization reflect a traditional agroecosystem that incorporates sustainable practices intended to maintain productivity with conservation of soil and water resources.
Introduction
Long-term knowledge of crops and agroecosystems is inherent to many traditional cultures and also relevant to current and future challenges in agriculture. This deep-time experience in farming a wide range of environments and conditions is relevant to current critical challenges such as response to climate change through development of more resilient crop varieties and sustainable agricultural systems that maintain productivity while protecting land and water resources (Altieri Citation2004; Fisher Citation2020; Gliessman Citation2014; Nabhan Citation2013; Reed and Ryan Citation2019).
Traditional crops and wild food plants are important in maintaining and redeveloping diets that promote good health. Historical changes in diets following cultural suppression and drastic social-economic change are a factor in health problems in many indigenous communities (Mihesuah and Hoover Citation2019). More data and understanding of the nutrient composition of traditional food crops such as maize, the subject of this paper, are needed to support efforts toward healthy foodways (Oas and Adams Citation2021). Landrace maize cultivars have enhanced nutritional value and health benefits relative to modern commercial hybrid maize (Colín-Chávez et al. Citation2020; Nankar et al. Citation2016; Ryu et al. Citation2013).
Domesticated in Mexico about 9000 years ago, maize has become a major crop globally, with the highest production of all cereal grains (FAOSTAT Citation2021; Kistler et al. Citation2020; Nuss and Tanumihardjo Citation2010; Palacios-Rojas et al. Citation2020). Maize continues to be essential to many American Indian agricultural peoples, including in the American Southwest (Adams Citation2015; Barreiro Citation1989; Bocinsky and Varien Citation2017; Fish Citation2004; Johannessen and Hastorf Citation1994; Mt.Pleasant and Burt Citation2010; Muenchrath and Salvador Citation1995; Oas and Adams Citation2021). Over millennia, a remarkable array of maize varieties and agricultural management strategies have been developed and adapted to a diverse range of environments and climates by indigenous farmers. Many traditional maize varieties continue to be grown, and are critical to world crop diversity (Jaradat Citation2013; Khoury et al. Citation2021).
In partnership with the Pueblo of Zuni, an experiment in growing traditional maize with water and nutrient treatments was conducted in traditional fields as part of a larger study of dryland (non-irrigated) Zuni agroecosystems and soils. The experiment provided data from replicated plots within two traditional Zuni fields, and a comparison of traditional and hybrid maize grown side-by-side under the same conditions. The purpose of this paper is to report on the composition and distribution of nutrients in the maize. A larger objective is to better understand the nutritional quality and related characteristics of traditional maize.
This paper is a direct continuation of a previous paper about the Zuni maize experiment that reported on maize growth and production (Muenchrath et al. Citation2017). Information essential to the current paper is provided herein. Readers interested in more information on the maize experiment can link directly to the 2017 paper (open access) through the DOI (https://doi.org/10.2993/0278-0771-37.2.172).
The Zuni live in the mesa landscape of the southeastern Colorado Plateau in present-day western New Mexico (). They and other indigenouspeoples are renowned for their skill in farming the arid to semiarid American Southwest over many generations. Archaeological and historical evidence documents Zuni dryland and irrigated fields that are at least 1000–3000 years old (Damp Citation2007; Homburg, Sandor, and Norton Citation2005), and maize has been grown in the region for more than 4000 years (Adams Citation2015; Merrill et al. Citation2009). More information about the environmental and agricultural setting of the Zuni study area is provided in the first paper (Muenchrath et al. Citation2017) and publications on other aspects of the agroecology project (Homburg, Sandor, and Norton Citation2005; Muenchrath et al. Citation2002; Norton et al. Citation2002, Citation2007b; Norton, Sandor, and White Citation2003, Citation2007a; Sandor et al. Citation2007).
Figure 1. Map of Zuni. Maize experiment fields (Laate Field and Weekoty Field, shown by black circles), located near the traditional farming villages of Nutria and Pescado (black squares). Muenchrath et al. (Citation2017). © (2017) Society of Ethnobiology.
Methods
The maize experiment was conducted in two traditional Zuni fields (Laate and Weekoty Fields) on alluvial fans in the eastern part of the Zuni Indian Reservation (). The fields, 11 km apart and at similar elevations (2072–2088 m), have a long record of historic and prehistoric farming. The experiment was done at both fields in 1997 and repeated in the Weekoty field in 1998, for a total of three field-years. The Laate field was eliminated from the study in 1998 because sheep corralled there during the intervening winter disrupted experiment treatments.
The experiment was conducted as a generalized randomized complete block design for each field-year. Two types of maize were tested in the cropping experiment: Zuni blue maize and an equal mix of two modern hybrids (referred to as “Hybrid” in this paper) recommended by New Mexico State University. The experiment included five treatments: precipitation-only, runoff water, runoff water and associated sediments, irrigation water to match amount and timing of applied runoff, and a treatment with more irrigation water (applied when needed to avoid crop water-deficit stress) plus N and P fertilizer. Treatments consisted of factorial combinations of the two cultivars and five water/nutrient treatments. Within each field, treatment combinations were randomly assigned to individual plots and were replicated three times for Zuni maize and twice with Hybrid maize. This design resulted in 25 plots per field-year, with each plot divided into 25 subplots. Maize data were collected only from the interior nine subplots of each plot to minimize border effects between plots. More details on experimental design for fields and plots (including plot maps), treatments, cultivars, and maize growth and production measurements are available in Muenchrath et al. (Citation2017).
Maize nutrient concentrations were measured in leaves and grain by plot for each field-year. At 50% flowering, the leaf opposite and below the apical ear of one random plant in each of the nine interior subplots of each plot was harvested and dried (Cerrato and Blackmer Citation1991). Grain samples consisted of 60 kernels combined from 15-kernel subsamples from four random ears from the dried median fresh weight subplot of each plot (Muenchrath et al. Citation2017). Sample preparation was completed and nutrients were analyzed at the Iowa State University Horticulture Plant Nutrition Laboratory. The following method description was provided by the lab. Leaves and grain were further dried in a forced-air oven at 67°C for 72 hours and ground to pass a 40-mesh screen in a Wiley mill. Total N was determined using a modified micro-Kjeldahl digestion procedure (Jones Citation1991; Nelson and Sommers Citation1980), in conjunction with a nitroprusside-salicylate assay (Wall et al. Citation1975) using flow injection analysis (Smith and Scott Citation1990). Each set of tissue samples was analyzed twice and their results averaged. Tissue elemental concentrations of P, K, S, Ca, Mg, Fe, Mn, Zn, Cu, B, Mo, and Na (Ni for 1997 leaf samples only) were measured using calibrated inductive coupled argon plasma emission techniques (ICP) (Jones Citation1977; Munter and Grande Citation1981) following dry ashing at 490°C and digesting in aqua-regia. The instrument was a Thermo Jarrell-Ash ICP/IRIS model with a charged injection device (Epperson et al. Citation1988). Plant standards and quality control checks for the N and ICP analyses were within ±5.0% range of true value, except for S and Fe, which require a special pre-analysis dry ashing treatment that was not available in the lab (Diane Shogren, written communication). Sulfur and Fe data were adjusted (S/0.84, Fe/0.485), based on average S and Fe recovery compared with the national standard, determined over many analyses. Nutrient data are expressed on a dry weight basis.
Methods for soil total N, total P, and available P (Olsen method) analyses, as well as organic C, pH (1:1 soil:water), bulk density, and particle-size distribution (texture) are given in Homburg, Sandor, and Norton (Citation2005). For these properties, soils were sampled by plot at 0–15 cm depth before planting and treatment application, and at two later dates for the Weekoty Field. These properties were also determined for one complete soil profile described for each field. Soil potentially mineralizable nitrogen (PMN, including nitrate-N and ammonium-N), sampled by plot at 0–20 cm depth before planting and treatment application and at the end of the first growing season, was determined by methods in White, Dreesen, and Loftin (Citation1997).
Analysis of variance (ANOVA) and correlation (Pearson) statistics for the maize nutrient data were performed using JMP Pro 14 to 16 (SAS Institute, Inc. 2021). Least square means (LS Means) of maize variables were used to adjust for the unbalanced design. Means were separated using t-tests. In those analyses in which a cultivar x treatment or field x cultivar interaction was evident in 2-way ANOVA, 1-way ANOVA was done. It was not possible to run an overall ANOVA model using all factors (treatment, cultivar, field, and year) because the experiment could only be conducted on the Laate Field for the first year due to later field disruption, and the treatments that could be applied varied in each field-year because of differences in runoff events. For the Laate Field 1997, all five treatments were run. For the Weekoty Field 1997, no runoff was available to apply so all plots other than the irrigation-fertilizer treatment were precipitation-only. For the Weekoty Field 1998, one limited runoff event occurred, allowing all treatments except runoff water, which reverted to precipitation-only.
All basic data on the plot basis for the three field-years are available in Supplementary Table S1: maize nutrient concentrations for grain and leaves, maize yields for grain and total aboveground biomass (kg ha−1 and kg plant−1), and soil properties. Data on nutrient and water inputs for each treatment are in Muenchrath et al. (Citation2017) Table 5 and Figure 5.
Results
Effects of the five water and nutrient treatments on maize nutrient composition, and a comparison of nutrient composition between Zuni blue maize and commercial hybrid maize, are presented. Data on relationships between nutrient composition (mainly N and P), soil nutrients and other properties, water inputs, and maize yields are also summarized.
Significant treatment and cultivar effects on maize nutrient composition were observed in all three field-years, but cultivar effects were more prevalent (). Significant cultivar differences (p ≤ .05) in grain nutrient concentrations occurred in over half of the 12 elements measured in three field-years, versus one-third significant treatment differences. Cultivar differences in leaf nutrient concentrations occurred in two-thirds of 13 elements measured, versus about half significant differences for treatments. Most treatment differences were between those in which fertilizer was applied and the other four other treatments.
Table 1. Nutrient concentrations in Zuni and Hybrid maize grain and leaves by field-year.
In the 2-way ANOVA, there was no significant (p ≤ .05) cultivar x treatment interaction for grain for any nutrient. For leaves, there was no significant cultivar x treatment interaction for any nutrient in the Laate Field, one case out of 13 elements in the Weekoty Field 1998, and five cases out of 13 elements for Weekoty in 1997. There was only one case, leaf Mn in the Weekoty Field 1997, where there was both a significant treatment and cultivar effect and a significant treatment x cultivar interaction.
Water-nutrient treatment effects on maize nutrient composition
A main question is whether additions of runoff water and sediment affected nutrient uptake or composition of the maize. Almost no significant differences in grain or leaf nutrient concentration were found between the precipitation-only and runoff treatments for the two field-years in which runoff treatments were applied (). In the experiment overall, no nutrient deficiency symptoms were observed, and leaf and grain N fall within the range reported in the literature for sufficient N concentration (see discussion). At the Laate Field, where runoff with sediment was applied four times, there was a trend toward increased maize N concentration in runoff treatments compared with the precipitation treatment for the Zuni blue maize, but differences were not statistically significant ().
Most of the few treatment differences were between those in which fertilizer was applied and the other four treatments. The applied fertilizer was primarily N and P, but analysis of the fertilizers indicated other nutrient elements were also present (Supplementary Table 2). Higher concentrations of the following nutrients were found in the irrigation-fertilizer treatment than in other treatments: Laate Field 1997 grain: N, leaf: N, S, Mg, Mn, Cu; Weekoty Field 1997 grain: no differences, leaf: N, P, S, Mn, Cu; Weekoty Field 1998 grain: no differences, leaf: P, S, Mn. Leaf Mo is the only element that trended lower in the irrigation-fertilizer treatment compared with the others for the three field-years.
Nutrient composition comparison between Zuni and hybrid maize
Another important question is whether the Zuni maize, as an open-pollinated landrace cultivar, differs in nutrient composition from modern hybrid maize. A number of cultivar differences in grain and leaf nutrient concentrations were found. For several nutrients, grain and leaf concentration differences among cultivars were inverse, and other patterns were also observed.
Across treatments, Zuni maize grain had higher concentrations of more kinds of nutrients than Hybrid maize grain, but Hybrid leaves were higher in more kinds of nutrients than Zuni leaves (). At the Laate Field in 1997, Zuni grain was higher than Hybrid in five nutrients (N, S, Mg, Zn, and Cu) and Hybrid was higher in three nutrients (P, K, B). Hybrid leaves were higher in five nutrients (N, P, Mn, Zn, and Cu) and Zuni was higher in four nutrients (K, Ca, Mg, and Fe). At the Weekoty Field in 1997, Zuni grain was higher than Hybrid in four nutrients (N, P, Mg, and Zn) and Hybrid was higher in one nutrient (B). Hybrid leaves were higher in five nutrients (N, P, Mn, Zn, and Cu) and Zuni was higher in two nutrients (K and Ca). At the Weekoty Field in 1998, Zuni grain was higher than Hybrid in six nutrients (N, P, K, Mg, Zn, and Cu) and Hybrid was higher in one nutrient (B). Hybrid leaves were higher in six nutrients (N, P, Mn, Zn, Cu, and B) and Zuni was higher in four nutrients (S, K, Ca, and Mg). The original seed used in the maize experiment (one sample analyzed) does not show the same differences as the harvested grain (Supplementary Table 3).
The data also show differences in maize nutrient concentration partitioning between grain and leaves, and in some nutrients in opposite directions between cultivars (, for N). Nitrogen and Zn were consistently higher in grain and lower in leaf for Zuni maize than Hybrid for all three field-years, and Cu in two of three field-years. Phosphorus shows this same pattern, but only at the Weekoty Field for both years. At the Laate Field, P was higher in both Hybrid grain and leaves (see field comparison section).
Figure 2. Nitrogen proportion in maize grain, expressed as N grain concentration as a percentage of N concentration in grain plus leaves (concentration units g kg−1). Bars are means with standard deviations. *, **, *** indicate differences in means between maize cultivars within treatments for each field-year at p ≤ .05, 0.01, and 0.001, respectively, using t-tests following 2-way ANOVA.
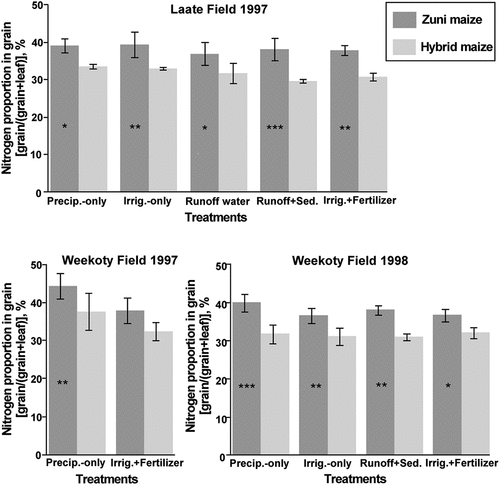
Different patterns were measured for other nutrients. For K, Hybrid had higher grain concentrations and lower leaf concentrations at the Laate Field, but Zuni maize had mostly higher concentrations in both grain and leaves both years at the Weekoty Field. Calcium was consistently higher in Zuni leaves, and Mg was consistently higher in Zuni grain. Manganese was consistently higher in Hybrid leaves. Boron was higher in Hybrid grain in all three field-years.
Effects of field location (soil differences) on maize nutrient composition
Differences in maize and soil nutrient composition were found between fields (, Supplementary Tables S4 and S5). These differences were measured in maize during the first year of the experiment for the precipitation-only treatment (no extra water or nutrient inputs) and for the non-fertilized treatments combined, and in maize plot soils before planting and treatment application. Direct nutrient comparisons of maize and soil were only possible with N and P because these were the only nutrients measured in both maize and soils.
Table 2. Comparison of nitrogen and phosphorus in maize and soil between the two traditional fields in the first year of the maize experiment.
Nitrogen was higher in both maize grain (but not leaves) and soils in the Weekoty Field than in the Laate Field. Grain N concentration (g kg−1) was higher by about 25%, and surface soil total N was higher by about 15% both on a mass concentration (g kg−1) and volume (kg ha−1) basis. The grain concentration difference was not present in the treatment with N fertilizer (Supplementary Table S4). Surface soil potentially mineralizable N (PNM) was also higher at the Weekoty Field, by 29% at the beginning of the growing season. As with grain, N fertilization mostly eliminated soil PNM differences (Supplementary Table S1).
Phosphorus also showed maize nutrient differences in both grain and leaves between fields that paralleled soil differences. Laate Field maize grain had 10–15% more and leaves about 40% more P on the mass concentration basis. For grain, the difference was due to Hybrid but not Zuni maize. Surface soil in the Laate Field was substantially higher in both total and available P than the Weekoty Field, and total P was higher in the Laate soil profile down to about 1 m depth. With P fertilizer inputs, maize grain P concentration differences between fields were eliminated, though leaf P concentration was still marginally higher in the Laate Field (p = .06).
Unlike the higher N grain concentration in the Weekoty Field, there were no differences between fields in grain N content on an areal (kg ha−1) or per plant basis (g plant−1) in the precipitation-only and non-fertilized treatments (Supplementary Tables S1 and S4). This is mostly because N content is calculated by N concentration x grain yield, and grain yields trended higher in the Laate Field (Supplementary Table S1; Muenchrath et al. Citation2017 Figure 6). In contrast, grain P content was significantly higher in the Laate Field in the non-fertilized treatments because both grain P concentration and yield trended higher in the Laate Field. With fertilization, P grain content was similar between fields.
For other nutrients without soil measurements, both grain and leaf Zn concentrations were higher at the Weekoty Field (). Cases of grain or leaf nutrient differences were Cu (higher in Weekoty Field grain), B (higher in Laate Field grain), Mg (higher in Weekoty Field leaves), and Mo (higher in Laate Field leaves). Some nutrient concentration differences between fields vary by cultivar. Besides grain P, K is also higher in the Laate Field in Hybrid but not Zuni grain. Magnesium is higher in the Weekoty Field in Zuni but not Hybrid grain.
Table 3. Comparison of nutrients/elements other than N and P in maize grain and leaves between the two traditional fields in the first year of the maize experiment.
Discussion
Main findings from the experiment on maize nutrient composition and quality are organized in this discussion by treatment effects, comparison of cultivars, and maize-soil relationships.
Treatments: runoff application and maize nutrients
Although nutrient differences among treatments were evident, they were mostly between the irrigation-fertilizer treatment and all others. The lack of response of maize to runoff applications in terms of both increased yield and nutrient concentrations compared with the precipitation-only treatment was unexpected because the runoff applied added somewhat more water and nutrients for the maize (mainly for Laate Field), and data indicate that the N and P in runoff sediment are higher than in the soils (Muenchrath et al. Citation2017: Figures 5–7, Table 5; Sandor et al. Citation2007: Table I; Sandor and Homburg Citation2015: Figure 2.2). Zuni farmers and researchers have generally observed reduced maize growth and nutrient deficiency after a few years of continuous production (Muenchrath et al. Citation2002; Sandor et al. Citation2007). For example, at one traditional Zuni field exhibiting nutrient deficiency symptoms (leaf chlorosis) after four straight years of maize production, maize growth was vigorous in an adjacent field in its second year of production after being fallowed for several years, showing the importance of fallow for nutrient replenishment. A key component of Zuni traditional agriculture is the contribution of watershed runoff and accompanying sediment and organic matter, plus eolian inputs, to building and maintaining soils and soil fertility (Cushing Citation1920; Homburg, Sandor, and Norton Citation2005; Muenchrath et al. Citation2002; Norton, Sandor, and White Citation2003, Citation2007a; Norton et al. Citation2007b; Sandor et al. Citation2007; Sandor and Homburg Citation2015).
One factor that may explain the lack of nutrient and yield differences between precipitation-only and runoff treatments is that soil nutrient levels at the time of the experiment may have already been sufficient due to the extended fallow period of at least seven years that preceded the experiment. Maize nutrient concentrations in all treatments mostly fall within the sufficiency ranges reported in other maize nutrient studies (see cultivar discussion), and no visual symptoms of nutrient deficiency were observed in any field-year.
Another possible factor is that runoff for treatment applications was limited to one year at each site. At the Laate Field, runoff was applied but only for one season because contamination from sheep kept in the fenced field after the first growing season ended the experiment there, and there may have been some suppression of nutrient availability during that initial season. Also, the method of applying runoff sediment separately from runoff water at the Laate Field (done for the largest runoff application to better quantify water and sediment inputs) may have suppressed nutrient availability to the maize. No runoff was available for application at the Weekoty Field in 1997, and in 1998 runoff was limited to one event with relatively small amounts of water and nutrients (little sediment) available for the runoff plots. A traditional Zuni practice was to wait to farm newly prepared fields for a year or more (Cushing Citation1920, 153, 165; Green Citation1979, 255), suggesting that more time would be required for runoff materials to decompose and contribute significant amounts of plant-available nutrients. More information on traditional field management is also presented in the observational study of Zuni farmer fields that was complementary to this maize experiment (Muenchrath et al. Citation2002).
Another probable factor is that water limited maize production more than nutrients. The amount of supplemental water applied to runoff plots represented a relatively small increase over that supplied by precipitation (Muenchrath et al. Citation2017 Figure 5), especially at the Weekoty Field in 1998. There may not have been enough of an increase in runoff application over precipitation to significantly increase yield. In modern commercial maize production, 500–600 mm is considered sufficient (Olson and Sander Citation1988), whereas total water inputs for precipitation-only and runoff treatments in this experiment ranged from 170–284 mm, and were 239–370 mm in the irrigation-fertilizer treatment. Perhaps nutrient inputs and levels in this experiment were mostly sufficient given the more limited water supply and considering that production is lower in traditional agriculture relative to intensive commercial maize production. Minimum water requirements for dryland maize production are estimated at 150 mm for the growing season and 250 mm annually (Shaw Citation1988; also see Benson Citation2010; Muenchrath and Salvador Citation1995; Strawhacker et al. Citation2020).
In this experiment, water inputs were positively correlated with maize grain yields (kg ha−1, r = 0.46, p < .001, plot basis) for all field-years combined, and higher for the 1997 fields (Supplementary Tables S1 and S6). Correlations of water input and total aboveground biomass are slightly higher than for grain. The higher water input-grain yield correlations for the 1997 Laate and Weekoty Fields (r = 0.74, p < .001) than the 1998 Weekoty Field (r = 0.43, p = .03) may partly be due to the irrigation water in the irrigation-fertilizer treatment being higher by 105–131 mm in 1997 than 1998. The positive water-yield correlations are primarily due to the effect of the irrigation-fertilizer treatment. It appears that the amount of added water input in the runoff treatments was not enough to increase yields over the precipitation-only treatment.
Cultivars
Comparing cultivars, Zuni maize grain had higher concentrations of more essential elements than Hybrid maize, and the opposite was true for nutrient concentrations in leaves (). For several elements, there was an inverse pattern in grain vs. leaf nutrient concentration between cultivars.
The higher N concentration in Zuni maize grain compared with the Hybrid grain indicates that the Zuni blue maize has a higher protein content. This agrees with a comparison of blue and commercial maize in New Mexico by Dickerson (Citation2003), who found that blue corn was a more complete protein source than white or yellow commercial dent corn. Other studies have found that landrace and older hybrid maize cultivars are generally higher in protein, and therefore more nutritious, than modern hybrid maize (Ciampitti and Vyn Citation2013; Jaradat Citation2013; Nankar et al. Citation2016; Ryu et al. Citation2013; Woli et al. Citation2018). Colored maize has additional nutritional benefits such as from anthocyanins in blue maize (Nankar et al. Citation2016; Ryu et al. Citation2013).
Another aspect of the N difference among cultivars in this study was that the Zuni maize had a higher percentage of grain N relative to leaves than the Hybrid maize, indicating that Zuni maize was able to partition or mobilize more of its protein into grain, adding to its higher nutritional quality. This pattern held for nearly all treatments in all field-years (). In comparing older and modern hybrid maize, Ciampitti and Vyn (Citation2013) found that while maize grain N concentration decreased over time, grain and non-grain (stover) N partitioning did not change. Differences in root architecture may play a role in nutrient acquisition and composition differences between landrace and hybrid maize, which should be explored in future research. Bousselot et al. (Citation2017) measured higher root-to-shoot ratios in several Southwest landraces, including the Zuni maize grown in this experiment, compared with a hybrid dent maize.
Results also indicate consistently higher nutritional quality of the Zuni maize grain in Mg and Zn in all three field-years (). Phosphorus and Cu were higher in the Zuni maize grain in two field-years. Zuni grain at the Weekoty Field was higher in P, and P as a percentage of grain plus leaf biomass for both field-years, but P was higher in Hybrid grain and leaves at the Laate Field (see field comparison section). Evidence of greater arbuscular mycorrhizal fungi activity in the Zuni maize possibly influenced P differences between the cultivars (Havener Citation1999; Sandor et al. Citation2007). Potassium trended higher in Zuni grain and leaves at the Weekoty Field. Boron was the one nutrient that was consistently higher in Hybrid grain. Dickerson (Citation2003) also found that blue maize was higher in Zn, and mostly higher in Fe, while dent maize was higher in B, as found in this study.
Although data from this and other research indicate that differences in grain nutrient quality between the Zuni and Hybrid maize are genetically based, other possible factors should be considered relating to yield differences between the cultivars. Hybrid maize grain yield trended higher on an areal basis (kg ha−1), but Zuni vegetative and total aboveground biomass trended higher on an areal basis. Zuni grain, and vegetative and total aboveground biomass, trended higher on a per plant basis (kg plant−1) (Muenchrath et al. Citation2017). These grain and biomass yield differences could lead to a dilution effect, in which nutrient concentrations are lower when yields are higher (Jarrell and Beverly Citation1981). This effect has been measured in maize growth studies, and “ … occurs when dry-weight accumulation increases at a faster rate than mineral-nutrient accumulation” (Riedell Citation2010, 869). In this case, it could be that N supply was limiting to the point that N concentration was lower at higher yields. This could also be true for P and other nutrients if they were limiting. The dilution effect, observed in maize and other crops since the 1940ʹs, is now primarily attributed to plant breeders selecting for higher crop yields (mainly increased carbohydrate content) but not nutritional quality (Davis Citation2009).
One way to test for a dilution effect is with correlation analysis: if there is a dilution effect, nutrient concentrations and yields should be negatively correlated (Davis Citation2009). This should be most pronounced in those treatments in which nutrients are most likely to be limiting (precipitation-only and non-fertilized treatments), and not present where sufficient levels of nutrients are available (especially the N and P fertilizer treatment). Ratios of N and P inputs in the fertilizer/non-fertilized treatments far exceed ratios of higher/lower yields between cultivars (Muenchrath et al. Citation2017 Table 5, Figures 6–7).
Although not all data needed to fully address the dilution effect question are available in this study, some relevant data are available. The results of correlation analyses are mixed and inconsistent by field-year, treatment, and cultivar (Supplementary Tables S1 and S7). Correlation trends also vary depending on how yield is expressed: areal or per plant basis, and for grain or aboveground total or vegetative biomass. For example, in the non-fertilizer treatments, some significant (p ≤ .10) negative correlations of grain N and P concentrations and grain and biomass yield occurred for Zuni maize in one field-year (Weekoty Field 1997). However, significant negative correlations mostly did not occur in the other two field-years for grain or biomass. Also, leaf N and P concentration correlations with grain and biomass were either not significant or were actually positive, suggesting a synergistic rather than dilution effect. Correlations between N and P concentration and yields in the fertilizer treatment were more difficult to evaluate because of low sample size (n = 5 across cultivar), but also were inconsistent by field-year, plant part (grain or leaf), nutrient (N or P), and areal or per plant yield basis.
These results support the hypothesis that nutrient concentration differences between cultivars primarily reflect genetic differences between Zuni and Hybrid maize other than those that produce a dilution effect. Another finding pointing to genetic cultivar differences is that nutrient difference patterns between cultivars tend to occur across all treatments, including the fertilizer treatment. Also, nutrient concentrations in all the maize mostly fall within the sufficiency ranges reported in other maize nutrient studies (see paragraph below), and no visual symptoms of nutrient deficiency were observed in any field-year. Still, to more fully address questions about genetically-caused cultivar nutrient differences and dilution effects, a nutrient balance and accumulation approach with nutrient analyses of all maize plant components and higher sample size would be needed. Treatments with separate irrigation and fertilization would also help in evaluating water and nutrients independently as factors potentially limiting maize yield and composition.
Overall, most nutrient concentrations in grain and leaves for both Zuni and Hybrid maize were within ranges measured for highly-managed modern hybrid maize, and in ranges considered sufficient for maize production (Brady and Weil Citation2008, 720; Flynn, Ball, and Baker Citation1999 for New Mexico). Sufficiency ranges are primarily based on nutrient concentrations in ear-leaves at flowering. Leaf N concentrations were slightly below the standard sufficiency range listed in Brady and Weil (Citation2008) for Zuni maize in non-fertilized treatments at the Laate Field (), but within nutrient sufficiency ranges listed in other sources (e.g., Jones, Eck, and Voss Citation1990). Also, this lower leaf N concentration should be considered in light of higher proportion of N in grain vs. leaves characteristic of the Zuni maize. Grain N concentrations of Zuni and Hybrid maize in this experiment are within ranges given for well-fertilized modern hybrid maize in several studies (e.g., Cerrato and Blackmer Citation1990; Ciampitti and Vyn Citation2013; Haegele et al. Citation2013, converting protein to N with the standard factor of 6.25).
Of the 11–12 essential nutrient elements measured in grain and leaves in this study, the only consistent exception to nutrient sufficiency was for grain S. Grain S was consistently low in both cultivars and fields, and for all treatments () compared with sufficiency ranges given in Brady and Weil (Citation2008) and other sources. Although grain S concentration was low, leaf S concentration was above critical levels listed for maize in New Mexico (Flynn, Ulery, and Lindemann Citation2010). Sulfur deficiency seems unlikely given the Cretaceous sedimentary rock soil parent material (Orr Citation1987). The reason for the relatively low grain S concentration is unknown.
Zuni blue maize grain nutrient concentrations were mostly within ranges reported in other studies of American Indian landrace maize in the Southwest and elsewhere (Calloway, Giauque, and Costa Citation1974; Dickerson Citation2003; Jaradat Citation2013; Nankar et al. Citation2016; Oas and Adams Citation2021; Ryu et al. Citation2013). The values of protein – N and mineral nutrients vary widely within and across these studies, which include a variety of maize endosperm types such as floury, flint, dent, sweet, and pop, as well as a range of growing environments and water and nutrient management. Where data for individual nutrients are available from two or more other studies, the Zuni blue maize has a higher range in Fe and Zn, and lower range in Na. Compared with the raw grain, nutrient concentrations commonly increase when maize is ground with mano and metate and when modified during cooking such as with addition of culinary ash and salt (Calloway, Giauque, and Costa Citation1974; Kuhnlein Citation1981; Oas and Adams Citation2021).
Field and soil comparisons
Differences in certain maize nutrients between fields probably reflect fine-scale soil and ecosystem differences between these field sites. Higher N concentration (grain) in the Weekoty Field and higher P concentration (Hybrid grain and leaves in both cultivars) in the Laate Field in the precipitation-only and non-fertilized treatments corresponded with soil N and P differences between the fields (, Supplementary Tables S4 and S5). Possible causal factors for maize and soil nutrient differences between fields are differences in geology and watershed geomorphology, soils, hydrology, and vegetation. Although both fields are within the same soil map unit at the broader scale (1:24,000) of the county soil survey (Zschetzsche Citation2005), at the finer scale of individual sites the Laate Field soil has a higher pH and contains more clay, whereas the Weekoty soil contains more sand (Supplementary Table S5). This overall soil texture difference relates to the higher proportion of shale-derived sediment in the Laate watershed versus more sandstone-derived sediment in the Weekoty watershed (Orr Citation1987). Specific chemical composition of the Cretaceous sedimentary rocks (Gallup Sandstone and Crevasse Canyon Formation) at these sites was not measured and would be needed to better evaluate possible differences relating to the observed soil and maize chemical composition differences.
It is also possible that geomorphic and hydrologic differences between fields partly explain the soil and maize chemical composition trends. For example, the Weekoty Field watershed (125 ha) is larger and more energetic than the Laate Field (7 ha), which may make for differences in the alluvial fan sediments that comprise the soil parent materials (Homburg, Sandor, and Norton Citation2005; Norton, Sandor, and White Citation2003, Citation2007a; Norton et al. Citation2007b; Sandor et al. Citation2007).
Although the Laate and Weekoty Field watersheds have similar elevations and general vegetation composition as indicated by their occurrence in the same map unit in the soil survey, at the finer field-watershed scale their slope aspects differ (Laate Field southwest-facing, and Weekoty Field east-facing), and specific differences in watershed vegetation and other ground cover may also cause differences in soil and maize chemical composition (Norton, Sandor, and White Citation2003, Citation2007a). For example, differences in soil organic matter and N may relate to watershed differences in N-fixing biological crust and shrubs, as well as other vegetation, and to differences in watershed sediment and organic material supply and transport to field soils. More detailed evaluation of soil and maize differences related to possible watershed geologic and ecosystem factors is needed.
In the case of phosphorus, available P in the Laate Field soil is higher despite its higher pH (mean 7.5 surface soil and range 7.5–8.2 in soil profile upper 1 m), which would decrease P availability compared with the Weekoty Field soil, whose pH is more in the optimal range for available P (mean 7.1 surface soil and range 6.8–7.6 in upper 1 m) (Supplementary Table S1 and S5; Brady and Weil Citation2008). By soil testing standards in New Mexico (Flynn Citation2015), the Laate Field surface soil averaged “Moderate” (range “Low” to “Moderate”) and values in the Weekoty Field averaged “Low” (range “Deficient” to “Low”).
In this study, soil differences in total and available N and P between the two fields during the same year corresponded with maize N and P concentration differences in the precipitation-only and combined non-fertilized treatments. Few other studies of ancient and traditional agriculture in the Southwest have explored relationships between soil and crop nutrient content. In a greenhouse test growing a traditional maize (Chapalote) in soils from paired prehistoric fields and uncultivated areas from Mimbres (New Mexico) without added fertilizer, differences in soil total N between sites corresponded with differences in N concentration and growth in young maize plants (Sandor and Gersper Citation1988). As in this study, the maize N difference between sites was eliminated with fertilization. Numerous modern agricultural studies relating maize and soil nutrient levels have been conducted, especially directed at calibrating maize yield response to soil levels of available N, P, K, and other nutrients (e.g., Brady and Weil Citation2008; Chatterjee and Clay Citation2016).
Conclusions
In this experimental study of maize nutrient composition and quality in traditional Zuni fields, we found patterns with water and nutrient treatments, maize cultivars, and maize-soil relationships. Among the most definitive findings is that Zuni blue maize grain was generally higher in nutritional quality than Hybrid maize. Zuni grain had a higher N (protein) content than Hybrid maize, and a higher proportion of N in grain vs. leaf, indicating an ability to partition more N and other nutrients into grain.
The key findings concerning management, maize, and soils point to further inquiry needed to better appreciate and understand Zuni and other traditional agricultural systems. More field-years with runoff application and monitoring are especially important. Application of runoff for just one year at each field was not enough to assess longer-term soil nutrient renewal processes. Also, treatments designed to more clearly separate effects of water and nutrients on maize production in dryland contexts would be essential to test their relative importance in maize production. Monitoring effects of runoff on soil and crop nutrients in different fields and environmental conditions would be helpful, given the considerable spatial and temporal variation in weather and climate, runoff hydrology, and soils.
Another research need is a more complete evaluation of nutrient content in all maize components, both aboveground and in roots, over growing seasons. This would allow a more quantitative assessment of nutrient accumulation by maize, and better testing of how maize landraces differ from hybrid maize, which would provide information for improving maize quality and maintaining biodiversity (Khoury et al. Citation2021). Coupled maize-soil studies would also allow quantitative nutrient balance evaluation of traditional maize production in terms of inputs and outputs. Interdisciplinary approaches to this maize research with tandem empirical studies and modeling would be most productive (Gillreath-Brown and Bocinsky Citation2017). Studies of traditional agroecosystems and crops in partnership with farmers and communities are always an important priority (Altieri Citation2004; Bocinsky and Varien Citation2017).
This part of the maize experiment at Zuni emphasized nutrients and their status in maize and soils. One of the key findings was that nutrient concentrations in both Zuni and Hybrid maize indicated nutrient sufficiency. This reflects the traditional Zuni agricultural system that has functioned for many centuries without conventional irrigation or fertilization in an arid to semiarid environment. Zuni soil and water management and nutrient renewal are connected to watershed ecosystem processes that supply organic matter and nutrients through runoff. This has been achieved by careful placement of fields on landscape positions such as alluvial fans and hillslope positions that receive runoff, and traditional methods to retain runoff and its nutrient-rich sediments (Sandor et al. Citation2007). Such sustainable agricultural strategies, and the diversity of maize and other crops adapted to arid conditions developed by the Zuni and other traditional peoples, are relevant to solving agricultural challenges in the face of climate change (Adams Citation2015; Nabhan Citation2013).
Although this agroecosystem is “low-tech” relative to intensive modern agriculture, there are important lessons to learn from its principles that are rooted in soil health and sustainability. Traditional Zuni dryland agriculture comprises sustainable practices intended to maintain productivity through conservation of soil and water resources.
Supplemental Material
Download Zip (7.5 MB)Acknowledgments
We are grateful to the Pueblo of Zuni for permitting us to conduct the study, and to all those with whom we worked on the project. We thank the Iowa State University Department of Horticulture Plant Nutrition Laboratory (Diane Shogren and others) for conducting the analyses of chemical elements in maize and fertilizer. We also thank Carl White and Chris Thomas (Department of Biology, University of New Mexico) for the sampling and analysis of soil potentially mineralizable nitrogen. Please see Muenchrath et al. (Citation2017) for more detailed acknowledgements about the maize experiment at Zuni. We thank the journal Editor and anonymous reviewers for their help.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/21683565.2022.2135670
Additional information
Funding
References
- Adams, K. R. 2015. The archaeology and agronomy of ancient maize (Zea mays L.). In Arid lands agriculture: Understanding the past for the future, ed. S. E. Ingram and R. C. Hunt, 15–53. Tucson: University of Arizona Press.
- Altieri, M. A. 2004. Linking ecologists and traditional farmers in the search for sustainable agriculture. Frontiers in Ecology and the Environment 2:35–42. doi:10.1890/1540-9295(2004)002[0035:LEATFI]2.0.CO;2.
- Barreiro, J., ed. 1989. Indian corn of the Americas: Gift to the world. Northeast Indian Quarterly 6: 1-2.
- Benson, L. V. 2010. Factors controlling Pre-Columbian and early historic maize productivity in the American Southwest, part 2: The Chaco Halo, Mesa Verde, Pajarito Plateau/Bandelier, and Zuni archaeological regions. Journal of Archaeological Method and Theory 18:61–109. doi:10.1007/s10816-010-9083-y.
- Bocinsky, R. K., and M. D. Varien. 2017. Comparing maize paleoproduction models with experimental data. Journal of Ethnobiology 37:282–307. doi:10.2993/0278-0771-37.2.282.
- Bousselot, J. M., D. A. Muenchrath, A. D. Knapp, and J. D. Reeder. 2017. Emergence and seedling characteristics of maize native to the Southwestern US. American Journal of Plant Sciences 8:1304–18. doi:10.4236/ajps.2017.86087.
- Brady, N. C., and R. R. Weil. 2008. The Nature and Properties of Soils. 14th ed. Upper Saddle River, NJ: Prentice Hall.
- Calloway, D. H., R. D. Giauque, and F. M. Costa. 1974. The superior mineral content of some American Indian foods in comparison to federally donated counterpart commodities. Ecology of Food and Nutrition 3:203–11. doi:10.1080/03670244.1974.9990382.
- Cerrato, M. E., and A. M. Blackmer. 1990. Relationships between grain nitrogen concentrations and the nitrogen status of corn. Agronomy Journal 82:744–49. doi:10.2134/agronj1990.00021962008200040019x.
- Cerrato, M. E., and A. M. Blackmer. 1991. Relationships between leaf nitrogen concentrations and the nitrogen status of corn. Journal of Production Agriculture 4:525–31. doi:10.2134/JPA1991.0525.
- Chatterjee, A., and D. Clay. 2016. Soil Fertility Management in Agroecosystems. Madison, WI: American Society of Agronomy, Inc. doi:10.2134/soilfertility.
- Ciampitti, I. A., and T. J. Vyn. 2013. Grain nitrogen source changes over time in maize: A review. Crop Science 53:366–77. doi:10.2135/cropsci2012.07.0439.
- Colín-Chávez, C., J. J. Virgen-Ortiz, L. E. Serrano-Rubio, M. A. Martínez-Téllez, and M. Astier. 2020. Comparison of nutritional properties and bioactive compounds between industrial and artisan fresh tortillas from maize landraces. Current Research in Food Science 3:189–94. doi:10.1016/j.crfs.2020.05.004.
- Cushing, F. H. 1920. Zuni Breadstuff. Indian Notes and Monographs 8. Museum of the American Indian. New York, NY: Heye Foundation.
- Damp, J. E. 2007. Zuni Emergent Agriculture: Economic Strategies and the Origins of the Zuni. In Zuni Origins: Toward a New Synthesis of Southwestern Archaeology, ed. D. A. Gregory and D. R. Wilcox, 118–32. Tucson: The University of Arizona Press.
- Davis, D. R. 2009. Declining fruit and vegetable nutrient composition: What is the evidence? HortScience 44:15–19. doi:10.21273/hortsci.44.1.15.
- Dickerson, G. 2003. Nutritional Analysis of New Mexico Blue Corn and Dent Corn Kernels. Las Cruces: New Mexico State University Cooperative Extension Service Guide H-233.
- Epperson, P. M., J. V. Sweedler, R. B. Bilhorn, G. R. Sims, and M. B. Denton. 1988. Applications of charge transfer devices in spectroscopy. Analytical Chemistry 60:327–35. doi:10.1021/ac00156a001.
- FAOSTAT. 2021. Crops and livestock products. URL: https://www.fao.org/faostat/en/#data/QCL. Accessed on November 15, 2021.
- Fish, S. K. 2004. Corn, crops, and cultivation in the North American Southwest. In People and Plants in Ancient Western North America, ed. P. E. Minnis, 115–66. Washington, DC: Smithsonian Books.
- Fisher, C. 2020. Archaeology for sustainable agriculture. Journal of Archaeological Research 28:393–441. doi:10.1007/s10814-019-09138-5.
- Flynn, R. 2015. Interpreting soil tests: Unlock the secrets of your soil. Las Cruces: New Mexico State University Cooperative Extension Service Circular 676.
- Flynn, R., S. T. Ball, and R. D. Baker. 1999. Sampling for Plant Tissue Analysis. Las Cruces: New Mexico State University Cooperative Extension Service Guide A-123.
- Flynn, R., A. Ulery, and W. C. Lindemann. 2010. Sulfur and New Mexico Agriculture. Las Cruces: New Mexico State University Cooperative Extension Service Circular 650.
- Gillreath-Brown, A., and R. K. Bocinsky. 2017. A dialogue between empirical and model-based agricultural studies in archaeology. Journal of Ethnobiology 37:167–71. doi:10.2993/0278-0771-37.2.167.
- Gliessman, S. R. 2014. Agroecology: The Ecology of Sustainable Food Systems. Third ed. Boca Raton, FL: CRC Press. doi:10.1201/b17881.
- Green, J., ed. 1979. Zuni: Selected Writings of Frank Hamilton Cushing. Lincoln: University of Nebraska Press.
- Haegele, J. W., K. A. Cook, D. M. Nichols, and F. E. Below. 2013. Changes in nitrogen use traits associated with genetic improvement for grain yield of maize hybrids released in different decades. Crop Science 53:1256–68. doi:10.2135/cropsci2012.07.0429.
- Havener, C. I. 1999. The Influence of Zuni Runoff Agriculture on Microbial Populations in Cultivated Fields. M.S. thesis, Department of Renewable Resources, University of Wyoming, Laramie. UMI Number EP22174.
- Homburg, J. A., J. A. Sandor, and J. B. Norton. 2005. Anthropogenic influences on Zuni soils. Geoarchaeology 20:661–93. doi:10.1002/gea.20076.
- Jaradat, A. A. 2013. Perceptual distinctiveness in Native American maize (Zea mays L.) landraces has practical implications. Plant Genetic Resources 11:266–78. doi:10.1017/S1479262113000063.
- Jarrell, W. M., and R. B. Beverly. 1981. The dilution effect in plant nutrition studies. In Advances in Agronomy 34, ed. N. C. Brady, 197–224. New York, NY: Academic Press. doi:10.1016/S0065-2113(08)60887-1.
- Johannessen, S., and C. Hastorf, eds. 1994. Corn and Culture in the Prehistoric New World. Boulder, CO: Westview Press.
- Jones, J. B. 1977. Elemental analysis of soil extracts and plant tissue ash by plasma emission spectroscopy. Communications in Soil Science and Plant Analysis 8:349–65. doi:10.1080/00103627709366727.
- Jones, J. B. 1991. Kjeldahl method for nitrogen determination. Athens, GA: Micro-Macro Publishing.
- Jones, J. B., H. V. Eck, and R. Voss. 1990. Plant analysis as an aid in fertilizing corn and grain sorghum. In Soil Testing and Plant Analysis, ed. R. L. Westerman, 521–47. Madison, WI: Soil Science Society of America. doi:10.2136/sssabookser3.3ed.c20.
- Khoury, C. K., S. Brush, D. E. Costich, H. A. Curry, S. de Haan, J. M. M. Engels, L. Guarino, S. Hoban, K. L. Mercer, A. J. Miller, et al. 2021. Crop genetic erosion: Understanding and responding to loss of crop diversity. New Phytologist 233:84–118. doi:10.1111/nph.17733.
- Kistler, L., H. B. Thakar, A. M. VanDerwarker, A. Domic, A. Bergström, R. J. George, T. K. Harper, R. G. Allaby, K. Hirth, and D. J. Kennett. 2020. Archaeological Central American maize genomes suggest ancient gene flow from South America. Proceedings of the National Academy of Sciences 117:33124–29. DOI: 10.1073/pnas.2015560117.
- Kuhnlein, H. 1981. Dietary mineral ecology of the Hopi. Journal of Ethnobiology 1:84–94.
- Merrill, W. L., R. J. Hard, J. B. Mabry, G. J. Fritz, K. R. Adams, J. R. Roney, and A. C. MacWilliams. 2009. The diffusion of maize to the southwestern United States and its impact. Proceedings of the National Academy of Sciences 106:21019–26. DOI: 10.1073/pnas.0906075106.
- Mihesuah, D. A., and E. Hoover, eds. 2019. Indigenous food sovereignty in the United States: Restoring cultural knowledge, protecting environments, and regaining health. Norman: University of Oklahoma Press.
- Mt.Pleasant, J., and R. F. Burt. 2010. Estimating productivity of traditional Iroquoian cropping systems from field experiments and historical literature. Journal of Ethnobiology 30:52–79. doi:10.2993/0278-0771-30.1.52.
- Muenchrath, D. A., M. Kuratomi, J. A. Sandor, and J. A. Homburg. 2002. Observational study of maize production systems of Zuni farmers in semiarid New Mexico. Journal of Ethnobiology 22:1–33.
- Muenchrath, D. A., and R. J. Salvador. 1995. Maize productivity and agroecology: Effects of environment and agricultural practices on the biology of maize. In Soil, water, biology, and belief in prehistoric and traditional Southwestern Agriculture, New Mexico Archaeological Council Special Publication No. 2. ed. H. W. Toll, 303–33. New Mexico Archaeological Council, C & M Press, Denver, CO.
- Muenchrath, D. A., J. A. Sandor, J. B. Norton, and J. A. Homburg. 2017. A maize experiment in a traditional Zuni agroecosystem. Journal of Ethnobiology 37:172–95. doi:10.2993/0278-0771-37.2.172.
- Munter, R. C., and R. A. Grande. 1981. Plant tissue and soil extract analysis by ICP-atomic emission spectroscopy. In Developments in atomic plasma spectrochemical analysis, ed. R. M. Barnes, 653–73. London: Heyden and Son.
- Nabhan, G. P. 2013. Growing Food in a Hotter, Drier Land: Lessons From Desert Farmers on Adapting to Climate Uncertainty. White River Junction, VT: Chelsea Green Publishing.
- Nankar, A., L. Grant, P. Scott, and R. C. Pratt. 2016. Agronomic and kernel compositional traits of blue maize landraces from the Southwestern United States. Crop Science 56:2663–74. doi:10.2135/cropsci2015.12.0773.
- Nelson, D. W., and L. E. Sommers. 1980. Total nitrogen analysis of soil and plant tissues. Journal of Association of Official Analytical Chemists 63:770–78.
- Norton, J. B., J. F. Bowannie, P. Peynetsa, W. Quandelacy, and S. F. Siebert. 2002. Native American methods for conservation and restoration of semiarid ephemeral streams. Journal of Soil and Water Conservation 57:250–58.
- Norton, J. B., J. A. Sandor, and C. S. White. 2003. Hillslope soils and organic matter dynamics within a Native American agroecosystem on the Colorado Plateau. Soil Science Society of America Journal 67:225–34. doi:10.2136/SSSAJ2003.2250.
- Norton, J. B., J. A. Sandor, and C. S. White. 2007a. Runoff and sediments from hillslope soils within a Native American agroecosystem. Soil Science Society of America Journal 71:476–83. doi:10.2136/sssaj2006.0019.
- Norton, J. B., J. A. Sandor, C. S. White, and V. Laahty. 2007b. Organic matter transformations through arroyos and alluvial fan soils within a Native American agroecosystem. Soil Science Society of America Journal 71:829–35. doi:10.2136/sssaj2006.0020.
- Nuss, E. T., and S. A. Tanumihardjo. 2010. Maize: A paramount staple crop in the context of global nutrition. Comprehensive Reviews in Food Science and Food Safety 9:417–36. doi:10.1111/j.1541-4337.2010.00117.x.
- Oas, S., and K. R. Adams. 2021. The nutritional content of five Southwestern U.S. indigenous maize (Zea Mays L.) landraces of varying endosperm type. American Antiquity. doi:10.1017/aaq.2021.131.
- Olson, R. A., and D. H. Sander. 1988. Corn Production. In Corn and Corn Improvement, ed. G. F. Sprague and J. W. Dudley, 3rd ed., Agronomy Monograph 18. 639–86. Madison, WI: American Society of Agronomy.
- Orr, B. R. 1987. Water resources of the Zuni Tribal Lands, McKinley and Cibola Counties, New Mexico. Water Supply Paper 2227. U.S. Geological Survey, Denver, CO.
- Palacios-Rojas, N., L. McCulley, M. Kaeppler, T. J. Titcomb, N. S. Gunaratna, S. Lopez-Ridaura, and S. A. Tanumihardjo. 2020. Mining maize diversity and improving its nutritional aspects within agro-food systems. Comprehensive Reviews in Food Science and Food Safety 19:1809–34. doi:10.1111/1541-4337.12552.
- Reed, K., and P. Ryan. 2019. Lessons from the past and the future of food. World Archaeology 51:1–16. doi:10.1080/00438243.2019.1610492.
- Riedell, W. 2010. Mineral‐nutrient synergism and dilution responses to nitrogen fertilizer in field‐grown maize. Journal of Plant Nutrition and Soil Science 173:869–74. doi:10.1002/jpln.200900218.
- Ryu, S. H., L. Werth, S. Nelson, J. C. Scheerens, and R. C. Pratt. 2013. Variation of kernel anthocyanin and carotenoid pigment content in USA/Mexico borderland land races of maize. Economic Botany 67:98–109. doi:10.1007/s12231-013-9232-9.
- Sandor, J. A., and P. L. Gersper. 1988. Evaluation of soil fertility in some prehistoric agricultural terraces in New Mexico. Agronomy Journal 80:846–50. doi:10.2134/agronj1988.00021962008000050033x.
- Sandor, J. A., and J. A. Homburg. 2015. Agricultural soils of the prehistoric Southwest: Known unknowns. In Arid lands agriculture: Understanding the past for the future, ed. S. E. Ingram and R. C. Hunt, 54–88. Tucson: University of Arizona Press.
- Sandor, J. A., J. B. Norton, J. A. Homburg, D. A. Muenchrath, C. S. White, S. E. Williams, C. I. Havener, and P. D. Stahl. 2007. Biogeochemical studies of a Native American runoff agroecosystem. Geoarchaeology 22:359–86. doi:10.1002/gea.20157.
- Shaw, R. H. 1988. Climate Requirement. In Corn and Corn Improvement, ed. G. F. Sprague and J. W. Dudley, 3rd ed., Agronomy Monograph 18. 609–38. Madison, WI: American Society of Agronomy.
- Smith, K. A., and A. Scott. 1990. Continuous-flow, flow-injection, and discrete analysis. In Soil Analysis, Modern Instrumental Techniques, ed. K. A. Smith, 183–227. 2nd ed. New York, NY: Marcel-Dekker.
- Strawhacker, C., G. Snitker, M. A. Peeples, A. P. Kinzig, K. W. Kintigh, K. Bocinsky, B. Butterfield, J. Freeman, S. Oas, M. C. Nelson, et al. 2020. A landscape perspective on climate-driven risks to food security: Exploring the relationship between climate and social transformation in the prehispanic U.S. Southwest. American Antiquity 85:427–51. doi:10.1017/aaq.2020.35.
- Wall, L. L., C. W. Gehrke, T. E. Neuner, R. D. Cathey, and P. R. Rexroad. 1975. Total protein nitrogen: Evaluation and comparison of four different methods. Journal of Association of Official Analytical Chemists 58:811–17. doi:10.1093/jaoac/58.4.811.
- White, C. S., D. R. Dreesen, and S. R. Loftin. 1997. Cobble mulch: An Anasazi water-conservation gardening technique. Paper presented at the Proceedings, 42nd Annual New Mexico Water Conference. New Mexico Water Resources Research Institute Report No. 304, New Mexico State University, Las Cruces.
- Woli, K. P., J. E. Sawyer, M. J. Boyer, L. J. Abendroth, and R. W. Elmore. 2018. Corn era hybrid macronutrient and dry matter accumulation in plant components. Agronomy Journal 110:1648–58. doi:10.2134/agronj2018.01.0025.
- Zschetzsche, S. A. 2005. Soil Survey of McKinley County Area, New Mexico. Washington, DC: U.S. Department of Agriculture – Natural Resources Conservation Service.