
Abstract
Despite using lots of progression in hypersensitivity, it is not yet a cure. Current treatment for allergy focuses on two approaches, including treats only the symptoms of allergy by utilizing drugs and the other is desensitization therapy (immunotherapy), which involves administration of specific allergens. The main obstacle to hypersensitivity permanent treatment is the IgE producing subclass of B-lymphocytes which is the source of IgE- producing in hypersensitive patients. Removal of these lymphocytes causes noticeable reduction in the levels of IgE and allergic responses significantly. In this paper, we proposed a novel complementary approach, which we have called the “nano-liposomes-based target toxicity machine (NBTTM)”, which controls hypersensitivity by removing the IgE producing B-lymphocytes. In this regard, our proposed nano-liposomes (pollen allergens/Aptamer/SCFV/MIP loaded lipid bilayer enclosing toxin interior) will be able to bind to any IgE markers in the lymphatic circulation. The nano-liposomes will bind to the IgE + atopic B cells and cause cell death by internalizing into B cells.
Introduction
Through a hundred years ago, Von Pirquet came up with allergy term to discern the harmful immune responses for the host, in case of protective immunity (Flöistrup et al. Citation2006). During the past decades, the allergic diseases have become one of the major health disturbances of American Academy of Allergy, Asthma, & Immunology (AAAAI).
The allergic response is mainly affected by environmental proteins (known as allergens) which lead to different clinical responses including allergic rhinitis (more commonly known as hay fever), allergic asthma, food allergy, allergic skin inflammation, ocular allergy, and mortal reaction named as anaphylaxis (Valenta et al. Citation2004).
Type I allergic diseases, caused by IgE-mediated hypersensitivity, affect more than 25% of the population in industrialized countries (Larche et al. Citation2006).
In general, effective event in the initiation of allergy is cross-linking of antigen-specific IgE bound to mast cells as an inducer of mast cell degranulation and rapid release of histamine, leukotrienes, and prostaglandins. Although several immunological pathways other than IgE, that can act in allergic responses induced-inflammation, need to be considered, focusing on Ag-specific IgE might be helpful for developing new therapeutic options and treatment of allergic diseases (He et al. Citation2013).
According to the important roles of B cells in humoral immunity and antibody production specially IgE along with antigen presentation, cytokine production, T-cell differentiation, and dendritic cell (DC) function, these cells account as the most important mediators of allergic responses thus manipulating IgE producing plasma cells can be defined as a new approach in allergy treatment (Watanabe et al. Citation2007).
Up to now, the current allergy therapies show promise, but also have some problems. Most important include of the long treatment period, the possibility of anaphylaxis shock, lack of safety, efficacy, and standardization (Barnes Citation2012).
Nano-liposomes are a small vesicle; the structure is similar to the cell membrane. Nano-liposomes can be filled with agents and used to deliver agents for disease especially in cancer treatment (Daraee et al. Citation2016; Rahman et al. Citation2016).
In this proposed manner, using nano-liposomes containing toxins, by targeting IgE + atopic B cells, the toxin is usually a cytotoxic protein derived from a bacterial or plant protein. In this hypothesis, we suggested to use the catalytic form of toxin neither other subunits, for example; the B chain is believed to mediate the entry of the A chain into a cell. In order to dampen the side effects, we will use the A chain of the toxin. In this method, the nano-liposome directs the toxin A chain to the target B cell and we planned to omit the production source of a topic IgE + via intra-lymphatic administration and its profit conclude of lack of hypersensitivity response (anaphylaxis shock), reduced toxin dose and reduced liposome administration times.
Hypothesis
The most important feature of these nano-liposomes is by delivering into the lymphatic stream, preferably by an intra-lymphatic injection. The nano-liposomes selectively bind to the atopic B cells bearing IgE. Once it bonds, the nano-liposomes is introduced into the cytoplasm where it eventually causes cell death. The IgE producing population of B cells responsible for hypersensitivity is eliminated. And the hypersensitive response which depends upon the production of IgE is likewise eliminated. To that end, three approaches could be used: pollen allergens, M1 prime SCFV, or aptamer and molecularly imprinted polymers (MIP) against CXCL13. In the present hypothesis, we suggest the novel application of the pollen allergens as a new, emerging technology in designing our special nano-liposomes, which we have called the "Targeted nano-liposomes" (T-nano-liposomes). In this way, the pollen allergens will serve as a template that can act as antigens to binding IgE + atopic B cells. For specific targeting of IgE + atopic B cells, SCFV, or aptamer against the prime M1 subunit of IgE serve auxiliary agents, and, the MIP will serve as a template that can precisely mimic the actions of an antibody or aptamer against chemokine ligand (e.g. CXCL13). MIPs could be simply synthesized through self-assembling and it’s cheaper than Ab and aptamers (Cheong et al. Citation2013; Li et al. Citation2006; Seidi et al. Citation2014). These target ligands provide the conditions for the presence of nano-liposomes in adjacent atopic B cells in lymph node. The pollen allergens could be easily collected from different geographical regions.
SCFV and aptamer have been considered as the most successful affinity tools with diagnostic and therapeutic applications in both fundamental and applied research nowadays. Another potential therapeutic target that we recommend should be selected. The homing of naive B cells (but not naive T cells) to the follicles during homeostasis depends on the expression of CXC-chemokine ligand 13 (CXCL13) by follicular dendritic cells (FDCs) and CXC-chemokine receptor 5 (CXCR5) as a cognate cellular receptor by naive B cells (Girard et al. Citation2012). Hence, CXCL13 MIP which is coated on the T-nano-liposomes surface can be a suitable ligand to trap the nano-liposomes in the B zone of the lymph node.
Nonetheless, they have several limitations, such as immune system activation. Pollen allergens are alternative affinity tools, which have received extensive attention. A major advantage of pollen allergens includes the efficient region specific natural production which needs to collect procedures and it is specific for IgE+ atopic B cell.
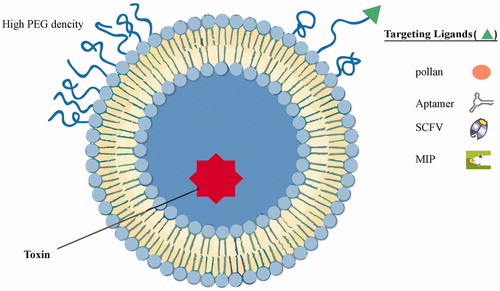
Thanks to the small size of nano-liposomes, which allow them to enter any small capillaries within tissues, and also their enhanced circulation time, it is expected that the application of pollen allergens in the structure of nano-liposomes will eliminate IgE+ atopic B cells in the body, especially in the lymph node. These nano-liposomes will circulate in the patients’ lymph node and bind intelligently to IgE+ Atopic B cells. The NBTTM requires the simple and easily accessible materials to start, i.e. targeted-nano-liposomes (pollen allergens-loaded lipid bilayer enclosing toxin interior). This protocol consists of two steps as follows: synthesis of targeted nano-liposomes and injection of targeted nano-liposomes to the patients’ lymph node. The proposed structure for T-nano-liposomes is shown in .
Figure 1. An illustrative structure of proposed T-nano-liposome. Liposomes can be surface functionalized to endow stealth through PEGylation and to promote receptor-mediated endocytosis by using targeting ligands such as pollen antigens, SCFV, aptamer, and MIP. PEGylation extends liposomal circulation half-life in vivo by reducing clearance, immune recognition, and the non-specific absorption of serum proteins. Polyethylene glycol (PEG) density determines its structure at the liposome surface.
General specifications
The efficacy of the approach can be affected by various factors listed below, which need to be optimized:
The size of the toxin that will determine its super activity.
The preferred shape of the toxin.
The thickness of the PLGA-PEG coating and the loading efficiency of the toxin are parameters that will influence the toxicity.
The preferred surface density of PEG, pollen, allergens, SCFV, MIP.
The administration doses should be optimized in order to avoid the hypersensitivity reaction.
Of note the promising advantages for NBTTM include the following:
Providing a treatment for seasonal allergies by specific pollen allergens.
It is not limited to affecting the hypersensitive response to a particular allergen.
Affecting the underlined cause of allergy rather than just treating symptoms.
It does not require repeated doses.
Shifting from Th2 to Th1 response.
Reducing undesirable side effects.
It is capable of being administered prior to the detection of any of the symptoms of allergy.
Evaluation of the hypothesis
Synthesis of targeted nanoparticles
Nano-liposome preparation
Currently, identified nanoparticles have provided an interesting basis for future applications. One group of nanoparticles is known as liposomes. Liposomes are spherical phospholipid structures which can range in size from submicrometer to several micrometers in diameter. The liposome can be used as a vehicle for drugs, therapeutic proteins, and nucleic acids (Allen and Cullis Citation2013). By shielding the surface of nanoparticles, including liposomes with hydrophilic components like polyethylene glycol (PEG) can minimize the toxicity and also enhance the cellular uptake of these nanoparticles. Additionally, PEGylation can control particle size, and also prevent aggregation of particles during storage. In this paper, the PEG-coated liposome is considered (Belliveau et al. Citation2012; Sawant and Torchilin Citation2012).
Finally, the nano-liposome encapsulated toxin will be prepared by a modified dehydration–rehydration vesicle (DRV) method. According to this method, phospholipid, cholesterol, PEG, and targeting ligands are dissolved in chloroform (solvent) in a round-bottomed flask and the chloroform is removed using a rotary evaporator at 50 °C. The thin lipid film is flushed with nitrogen gas to ensure complete chloroform removal prior to rehydration with distilled water/sucrose. Sucrose is used to stabilize the liposomes during freeze drying. The lipid suspensions are vortexed for 2 min to form multilamellar vesicles (MLVs) and then sonicated for 5 min. The resulting mixtures are centrifuged at low speed (400 × g, 10 min at 4 °C) to remove large vesicles. The suspension of small unilamellar vesicles (SUVs) is then mixed with the toxin. The mixture is rapidly frozen (−20 °C) and freeze-dried overnight under vacuum in a Hetosicc freeze-dryer. The powdered formulations are stored at 4 °C until use ().
Figure 2. Schematic representation of the steps involved in the nano-liposomes-based target toxicity machine.
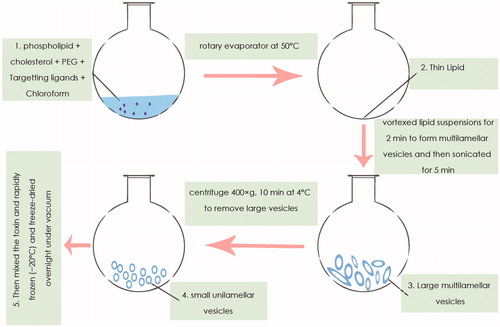
For rehydration, distilled water is added, vortexed, and incubated for 30 min at 50 °C. The step is repeated with phosphate-buffered saline (PBS) and after that, PBS is added again and the mixture is vortexed and incubated for another 30 min at 50 °C. The non-entrapped material is removed following three rounds of PBS wash (18,300 × g for 15 min at 4 °C). The extent of the liposome-entrapped toxin is quantitated by agar diffusion microbiological assay. For this purpose, lipid vesicles are lysed with 0.2% Triton X-100 (Gregoriadis et al. Citation1999; Mugabe et al. Citation2006).
Conjugation could be approached via different modifications including amine modification, carbohydrate modification, disulfide modification, and indirect methods. Procedures have described, in the following order.
Amine modification
Modification of the protein amine groups is defined as the most commonly used method to produce protein-conjugated liposome (especially in antibody–liposome conjugates). In early procedures, the used crosslinking agents, such as l-ethyl-3–(3-dimethylaminopropyl) carbodiimide hydrochloride (EDC) in the presence of preformed liposomes containing a lipophilic carboxylic acid (Endoh et al. Citation1981). N-Hydroxysuccinimide (NHS) is another early approach which is involved in protein modification of proteins via activated fatty acids (Huang et al. Citation1980). EDC have been used in conjunction with NHS to activate acidic utilities on liposomes, which were then conjugated to the amino groups on proteins. Another conjugation reaction can be achieved using heterobifunctional strategy, in this method, thiol or the maleimide group was protected, 3-(2-pyridyldithio)propionic acid-N-hydroxysuccinimide ester (SPDP), S-acetylthioglycolic acid N-hydroxysuccinimide ester (SATA), and 4-(p-maleimidophenyl) butyric acid N-hydroxysuccinimide ester (SMPB) are examples of cross-linkers (Barbet et al. Citation1981; Hansen et al. Citation1995) ().
Figure 3. Scheme of molecular imprinting against CXCl 13. CXCL13 receptors are chosen as a target and an imprint molecule for this hypothesis. The first step is to consider the CXCL13 monomers and template (template is a constant, short peptide sequence illustrative of an available fragment of a larger protein, holes on the template is representative of the active sites in the CXCL13 template). Afterward, all elements of the MIPs are combined and allowed to be self-assembled to form the cross-linked polymer with the template (pre-polymerization). After polymerization, monomers, and the surrounding matrix are cleaved from the template molecules. The resulting targeted MIPs will be able selectively to bind to CXCL13.
Carbohydrate modification
Oxidation of the proteins carbohydrate by sodium periodates produces aldehyde groups that can be used to conjugate the proteins to liposomes. In this method, the glycosylation region is important which is in the structure-depending region but not on functional region (Domen et al. Citation1990).
Disulfide modification
In this method, disulfide bonds cleaved with DTT or 2-mercaptoethylamine (2-ME), these sulfhydryl groups have been used to conjugate protein to maleimide-derivatized liposomes (Kirpotin et al. Citation1997).
Indirect methods
In this method, protein conjugated to the liposome with the secondary receptor–ligand systems such as liposome–PEG and liposome–avidin. Lipid–PEG–hydrazide derivative has been reported to couple oxidized protein to liposomes but effectively target biotinylated proteins. In this article, we proposed lipid–PEG–hydrazide because of the board application of PEG in the conjugated agent (Longman et al. Citation1995; Susan Citation2000).
Consequences of the hypothesis – clinical significance
An optimized dose of T-nano-liposomes will be injected into patients intra-lymphatically and, a patient’s IgE + atopic B cells will be applied to the NBTTM. As mentioned earlier, NBTTM is a device that uses a toxin to kill IgE + atopic B cells in a lymphatic system. By the intra-lymphatic injection of T-nano-liposomes, NBTTM will find their target IgE + Atopic b cells of lymph node B cells by IgE-pollen, Aptamer/SCFV-prime M1, and MIP-CXCl13 binding, then nano-liposomes containing toxin will internalize to atopic B cells and eventually they are going to cause the cell death ().
Figure 4. Schematic diagram of B cell activation by allergens and proposed “NBTTM” to eliminate of IgE + B cell from the patients’ body treated with T-nano-liposome.
Conclusion and perspective
Allergic sensitization is known as a multifactorial process which involves an exaggerated immune response to allergens such as pollens. An inhaled form of immediate allergic reactions is included by most common allergens, the plant pollens. Here and now, up to 30% of industrialized countries population put up with the IgE-mediated allergic disease, which termed as type I hypersensitivity reaction or type I allergies (Fooke Citation2012; Senti and Kundig Citation2016). This process is characterized by the primary production of allergen-specific IgE antibodies against the allergen which bound to their receptors on the surface of mast cells and basophils, which is called the sensitization. The direct consequence of this cross-linking will be the release of histamine of basophils and subsequent symptoms will occur, which forms the immediate phase of the allergic reaction () (Larche et al. Citation2006; Lessof Citation2012).
Over the years, whether allergic therapies achieved durable and curative responses in patients or not, they are still hotly debated. Common therapies in allergy reactions include anti-histamine and chemokine blocking like. The promising blocking concludes of TH2 cytokines, IL-4 and IL-13 which are responsible for class switching of IgE production by B cells. Also, production of IL-4, IL-5, IL-9, and IL-13 by TH2 cells provides the development and recruitment of Eosinophils and differentiation of mast cells (Larche et al. Citation2006). These blockers hinder some mediator and metabolites, but in next exposure to the same antigen, we should use these blockers over and over. So we just have the sectional treatment, not the defining one. But in our plan, the treatment period last longer than this sectional treatment and patients would be protected against allergy in all their longevity.
The next therapy is using various kinds of immunotoxins. Like other therapies, they have some drawbacks like the toxins side effects, this toxin does not have any typical shield, therefore, leads to lots of side effects like destructive reactions to natural cells. This toxin will target one part so the efficacy is low. In our plan, the nanoparticle is against several targets, so will appear in more effective way. As immunotoxins are not used in intra-lymphatic manner, then we were set to provide this kind of administration in this plan.
State-of-the-art allergy treatments include allergy immunotherapy that is considered as a vital treatment for allergic patients and can induce an allergen-specific immune response. In 1911, allergen-specific immunotherapy (SIT) is defined as repeated administration of disease-extorting allergens to induce tolerance clinically. The side effects of SIT which limit its broad application are mostly based on the allergen type, include of allergen applied, the formulation of extract, its potency, selecting the up dose regime, the state of disease severity prior to treatment, and premedication. Also, the highly cross-reactive allergens exist in many biological sources, for instance, profiling which is present in plant pollen. Therefore, because of the diversity in allergen extracts, the precise distinguishing the allergen which is responsible for the clinical symptom is yet a hurdle. All in all urticarial, local allergic reactions and frequent anaphylaxis have been reported. The improved approaches for providing the efficacy specific-allergen immunotherapy came up with using novel adjuvants, modified allergens, mostly used highly purified or recombinant allergens and surrogate means of administration (Burks et al. Citation2013; Focke-Tejkl et al. Citation2015; Fooke Citation2012; Jutel and Akdis Citation2011; Larche et al. Citation2006; Sicherer and Leung Citation2015; Winther et al. Citation2006). In the case of allergen-SIT, there is no clue of direct B-cell tolerance (Akdis et al. Citation1997). Recent studies have provided insights into modern tools of molecular biology against allergy include of using immunogenic and well-characterized epitopes in vaccine strategies and novel cellular test capable of elevated diagnostic avail and histamine measurement in specialized laboratories (Fontenot and Peebles Citation2013). In our plan, the predicted time for abating the B cell colony is estimated about 15 d. In such wise, patients go through sooner outcome.
The NBTTM offers new avenues for early and late treatment of allergy and hypersensitivity reactions.
Here, we highlighted some remarkable advantages: using reduced toxin dosage that turns out in elevated efficacy, using reduced liposome administration times, the intra-lymphatic injection, which renders to rapid feedback and the nanoparticle attaches to its target sooner. In spite of current therapies like immunotoxins, the toxins in contact with other living cells cause a lot of side effects for natural cells, but in our plan, toxins are kept inside the liposomes so the side effects will be shrunk, and in immunotoxin therapy, the toxin sets the goal on just one target, but in our proposed method, we mark several targets such as SCFV, aptamer, pollen itself to bound IgE in B cells, and also MIP to bind its ligand, CXCL13. So the B cell trafficking will occur.
References
- Akdis C, Akdis M, Blesken T, Wymann D, Alkan S, Muller U, Blaser K. 1997. Epitope-specific T cell tolerance to phospholipase A 2 in bee venom and peptide immunotherapy. Recovery by IL-2 and IL-15 in vitro. Immunol Lett. 56:331.
- Allen TM, Cullis PR. 2013. Liposomal drug delivery systems: from concept to clinical applications. Adv Drug Deliv Rev. 65:36–48.
- Barbet J, Machy P, Leserman LD. 1981. Monoclonal antibody covalently coupled to liposomes: specific targeting to cells. J Supramol Struct Cell Biochem. 16:243–258.
- Barnes PJ. 2012. Severe asthma: advances in current management and future therapy. J Allergy Clin Immunol. 129:48–59.
- Belliveau NM, Huft J, Lin PJ, Chen S, Leung AK, Leaver TJ, et al. 2012. Microfluidic synthesis of highly potent limit-size lipid nanoparticles for in vivo delivery of siRNA. Mol Ther—Nucleic Acids. 1:e37.
- Burks AW, Calderon MA, Casale T, Cox L, Demoly P, Jutel M, Nelson H, Akdis CA. 2013. Update on allergy immunotherapy: American academy of allergy, asthma & immunology/European academy of allergy and clinical immunology/PRACTALL consensus report. J Allergy Clin Immunol. 131:1288–1296.e3.
- Cheong WJ, Yang SH, Ali F. 2013. Molecular imprinted polymers for separation science: a review of reviews. J Sep Sci. 36:609–628.
- Daraee H, Etemadi A, Kouhi M, Alimirzalu S, Akbarzadeh A. 2016. Application of liposomes in medicine and drug delivery. Artif Cells Nanomed Biotechnol. 44:381–391.
- Domen PL, Nevens JR, Mallia AK, Hermanson GT, Klenk DC. 1990. Site-directed immobilization of proteins. J Chromatogr a. 510:293–302.
- Endoh H, Suzuki Y, Hashimoto Y. 1981. Antibody coating of liposomes with 1-ethyl-3-(3-dimethyl-aminopropyl)carbodiimide and the effect on target specificity. J Immunol Methods. 44:79–85.
- Flöistrup H, Swartz J, Bergström A, Alm JS, Scheynius A, van Hage M, et al. 2006. Allergic disease and sensitization in Steiner school children. J Allergy Clin Immunol. 117:59–66.
- Focke-Tejkl M, Weber M, Niespodziana K, Neubauer A, Huber H, Henning R, et al. 2015. Development and characterization of a recombinant, hypoallergenic, peptide-based vaccine for grass pollen allergy. J Allergy Clin Immunol. 135:1207–1217.e11.
- Fontenot AP, Peebles RS Jr. 2013. Allergy and hypersensitivity. Curr Opin Immunol. 25:736.
- Fooke M. 2012. Novel approaches for the in-vitro diagnosis of type I allergies. Multidisciplinary Approaches to Allergies. Berlin: Springer.
- Girard JP, Moussion C, Förster R. 2012. HEVs, lymphatics and homeostatic immune cell trafficking in lymph nodes. Nat Rev Immunol. 12:762–773.
- Gregoriadis G, Mccormack B, Obrenovic M, Saffie R, Zadi B, Perrie Y. 1999. Vaccine entrapment in liposomes. Methods. 19:156–162.
- Hansen CB, Kao GY, Moase EH, Zalipsky S, Allen TM. 1995. Attachment of antibodies to sterically stabilized liposomes: evaluation, comparison and optimization of coupling procedures. Biochim Biophys Acta – Biomembranes. 1239:133–144.
- He SH, Zhang HY, Zeng XN, Chen D, Yang PC. 2013. Mast cells and basophils are essential for allergies: mechanisms of allergic inflammation and a proposed procedure for diagnosis. Acta Pharmacol Sin. 34:1270–1283.
- Huang A, Huang L, Kennel SJ. 1980. Monoclonal antibody covalently coupled with fatty acid. A reagent for in vitro liposome targeting. J Biol Chem. 255:8015–8018.
- Jutel M, Akdis C. 2011. Immunological mechanisms of allergen-specific immunotherapy. Allergy. 66:725–732.
- Kirpotin D, Park JW, Hong K, Zalipsky S, Li WL, Carter P, Benz CC, Papahadjopoulos D. 1997. Sterically stabilized anti-HER2 immunoliposomes: design and targeting to human breast cancer cells in vitro. Biochemistry. 36:66–75.
- Larche M, Akdis CA, Valenta R. 2006. Immunological mechanisms of allergen-specific immunotherapy. Nat Rev Immunol. 6:761–771.
- Lessof MH. 2012. Immunological and Clinical Aspects of Allergy. Berlin: Springer Science & Business Media.
- Li Y, Yang HH, You QH, Zhuang ZX, Wang XR. 2006. Protein recognition via surface molecularly imprinted polymer nanowires. Anal Chem. 78:317–320.
- Longman SA, Cullis PR, Bally MB. 1995. A model approach for assessing liposome targeting in vivo. Drug Deliv. 2:156–165.
- Mugabe C, Azghani AO, Omri A. 2006. Preparation and characterization of dehydration-rehydration vesicles loaded with aminoglycoside and macrolide antibiotics. Int J Pharm. 307:244–250.
- Rahman M, Kumar V, Beg S, Sharma G, Katare OP, Anwar F. 2016. Emergence of liposome as targeted magic bullet for inflammatory disorders: current state of the art. Artif Cells Nanomed Biotechnol. 44:1–12.
- Sawant RR, Torchilin VP. 2012. Challenges in development of targeted liposomal therapeutics. AAPS J. 14:303–315.
- Seidi K, Eatemadi A, Mansoori B, Jahanban-Esfahlan R, Farajzadeh D. 2014. Nanomagnet-based detoxifying machine: an alternative/complementary approach in hiv therapy. J AIDS Clin Res. 5:304
- Senti G, Kundig TM. 2016. Novel delivery routes for allergy immunotherapy: intralymphatic, epicutaneous, and intradermal. Immunol Allergy Clin North Am. 36:25–37.
- Sicherer SH, Leung DY. 2015. Advances in allergic skin disease, anaphylaxis, and hypersensitivity reactions to foods, drugs, and insects in 2014. J Allergy Clin Immunol. 135:357–367.
- Susan S. 2000. Antibody conjugation methods for active targeting of liposomes. Methods Mol Med. 25:51–68.
- Valenta R, Ball T, Focke M, Linhart B, Mothes N, Niederberger V, et al. 2004. Immunotherapy of allergic disease. Adv Immunol. 82:105–153.
- Watanabe R, Fujimoto M, Ishiura N, Kuwano Y, Nakashima H, Yazawa N, et al. 2007. CD19 expression in B cells is important for suppression of contact hypersensitivity. Am J Pathol. 171:560–570.
- Winther L, Arnved J, Malling HJ, Nolte H, Mosbech H. 2006. Side‐effects of allergen‐specific immunotherapy. A prospective multi‐centre study. Clin Exp Allergy. 36:254–260.