
ABSTRACT
Numbers of listeriosis illnesses have been increasing in Germany and the European Union during the last decade. In addition, reports on the occurrence of antibiotic resistance in Listeria monocytogenes in clinical and environmental isolates are accumulating. The susceptibility towards 14 antibiotics was tested in a selection of clinical L. monocytogenes isolates to get a more precise picture of the development and manifestation of antibiotic resistance in the L. monocytogenes population. Based on the population structure determined by core genome multi locus sequence typing (cgMLST) 544 out of 1220 sequenced strains collected in Germany between 2009 and 2019 were selected to cover the phylogenetic diversity observed in the clinical L. monocytogenes population. All isolates tested were susceptible towards ampicillin, penicillin and co-trimoxazole – the most relevant antibiotics in the treatment of listeriosis. Resistance to daptomycin and ciprofloxacin was observed in 493 (91%) and in 71 (13%) of 544 isolates, respectively. While all tested strains showed resistance towards ceftriaxone, their resistance levels varied widely between 4 mg/L and >128 mg/L. An allelic variation of the penicillin binding protein gene pbpB3 was identified as the cause of this difference in ceftriaxone resistance levels. This study is the first population structure-guided analysis of antimicrobial resistance in recent clinical isolates and confirms the importance of penicillin binding protein B3 (PBP B3) for the high level of intrinsic cephalosporin resistance of L. monocytogenes on a population-wide scale.
Introduction
Listeria monocytogenes is an important foodborne pathogen and the causative agent of listeriosis, an illness with symptoms ranging from gastroenteritis to septicemia, meningoencephalitis and miscarriage in pregnant women. L. monocytogenes infections are mostly associated with ready-to-eat foods, as well as milk products, meat, fish and vegetables [Citation1]. Case numbers of listeriosis have been increasing during the last years. While between 2001 and 2010, 372 ± 101 listeriosis cases were reported per year in Germany, the average listeriosis case number between 2011 and 2019 rose to 617 ± 135 cases, with 699 notified cases in 2018 [Citation2]. The incidence of listeriosis is relatively low (0.3–0.6 per 100,000 persons in Europe and North America) compared to other gastrointestinal infections [Citation3]. However, fatality rates range between 7% and 30% despite antibiotic treatment [Citation4,Citation5]; even though L. monocytogenes is susceptible to a variety of antibiotics in vitro, it is one of the most fatal gastrointestinal foodborne bacterial pathogens.
The incubation period of listeriosis ranges from 1 to 67 days [Citation6]. This rather long time frame complicates back-tracing of food vehicles through patient interviews and thus often has hampered the identification of outbreak sources. Whole genome sequencing (WGS)-based subtyping techniques, such as core genome multi locus sequence typing (cgMLST), have been implemented recently in many countries to improve disease cluster recognition and compare clinical and food isolates. This has enormously facilitated the identification of infection sources of listeriosis outbreaks [Citation7–12].
The standard therapy for listeriosis is ampicillin or penicillin, frequently combined with gentamicin. While ampicillin or penicillin alone was reported to be only bacteriostatic, a bactericidal synergism of these antibiotics with gentamicin has been observed against L. monocytogenes in vitro [Citation13,Citation14]. However, the effectivity of the combination therapy has been questioned in retrospective studies investigating the outcome of listeriosis treated either with the combination of both antibiotics versus penicillin monotherapy, with no benefit of the combined treatment on the patient´s outcome [Citation15,Citation16] as well as in a recent study in a listeriosis mouse model [Citation17]. As an alternative, treatment with trimethoprim/sulfamethoxazole (hereinafter referred as co-trimoxazole) has been applied successfully in patients allergic to β-lactam antibiotics [Citation18]. Meropenem is occasionally applied in listeriosis treatment, but therapy failure and mortality rate is higher under these conditions [Citation19,Citation20].
As previously reported, resistance to the clinically used antibiotics is rare in clinical isolates of L. monocytogenes [Citation21–23], however, recent studies report increasing numbers of antibiotic-resistant environmental isolates, including isolates from animals, food and food-processing plants [Citation24–26]. This observation is alarming since there is evidence that the increase of minimal inhibitory concentrations (MIC) observed in environmental strains also manifested in clinical strains later on [Citation27]. Therefore, monitoring the development of antibiotic resistance in clinical isolates is of utmost importance to ensure appropriate antibiotic therapy of listeriosis in the future.
Beside the potential emergence of resistance to antibiotics used in standard therapy, L. monocytogenes is intrinsically resistant to third-generation cephalosporins such as ceftriaxone [Citation14,Citation28], often used to treat bacterial meningitis. Hence, as long as L. monocytogenes cannot be ruled out as the causative agent, co-administration of ceftriaxone or other cephalosporins with ampicillin is required [Citation29]. Several factors including the penicillin binding protein PBP B3, encoded by the lmo0441 gene, contribute to the intrinsic cephalosporin resistance of L. monocytogenes [Citation30,Citation31]. A L. monocytogenes mutant lacking lmo0441 has strongly reduced cephalosporin resistance but did not reveal any other obvious phenotypes [Citation30,Citation32], suggesting that PBP B3 has a function specifically required during cephalosporin exposure.
Based on genome sequence data, we here designed a selection of 544 clinical L. monocytogenes strains. This strain selection covers the entire phylogenetic biodiversity observed among strains isolated from human infections in Germany between 2009 and 2019, as it includes representatives of listeriosis outbreak clusters as well as isolates obtained from all sporadic cases. This selection was screened for antibiotic susceptibility against 14 clinically relevant antibiotics to describe the current antibiotic resistance levels of clinical L. monocytogenes strains on a population-wide scale, which led to the discovery of pbpB3 mutations associated with reduced levels of cephalosporin resistance.
Materials and methods
L. monocytogenes strains and growth conditions
All L. monocytogenes strains used within this study were originally received from different senders of the German health care system by the consultant laboratory for Listeria of the Robert Koch Institute. Identity and molecular PCR serogroups were determined by multiplex PCR as previously described [Citation33–35] at arrival and the received strains were archived in an in-house strain collection in 50% glycerol at −80°C. For antibiotic susceptibility testing, individual strains were grown on brain heart infusion (BHI) broth (# 211059, BD-BBL, Franklin Lakes, USA) or BHI agar plates (# CM0375, Oxoid, Basingstoke, UK) at 37°C. The strains used in this study are summarized in the supplementary Table S1.
Construction of plasmids and strains
For expression of pbpB3 variants in L. monocytogenes, pbpB3 alleles of the strains 17-04405 (allele type 49), 18-00287 (allele type 20), 18-00792 (allele type 4), 18-02573 (allele type 56) and 18-04540 (allele type 13) were amplified by PCR using the primers MF19 (5’- CGCGCCATGGATGGCTAGTTATGGTGGGAAAAAG) and MF20 (5’- CGCGGTCGACTTATTTATACATACTTTCAATAACTGGTTTTAGC). Fragments were cloned into plasmid pIMK3 [Citation36] using NcoI/Sall (NEB, Ipswich, USA). The sequence of the cloned inserts was confirmed by Sanger sequencing, the corresponding plasmid was introduced into strain LMJR41 (ΔpbpB3), which was constructed in a previous study [Citation32], by electroporation [Citation36] and transformants were selected on BHI agar plates containing 50 mg/L kanamycin. Correct plasmid insertion at the attB site of the tRNAArg was confirmed by PCR. The sequences of the above mentioned pbpB3 alleles were submitted to NCBI GenBank (MT383155-MT383119).
Genome sequencing
For genome sequencing, chromosomal DNA was extracted using the GenElute Bacterial Genomic DNA Kit (Sigma-Aldrich, St. Louis, USA). One ng of the chromosomal DNA obtained was used in a library preparation using the Nextera XT library preparation kit (Illumina, San Diego, USA) according to manufacturer’s instructions. Sequencing was performed on Illumina MiSeq, NextSeq or HiSeq 1500 instruments, using either the MiSeq Reagent Kit v3 (600-cycle kit) or the HiSeq PE Rapid Cluster kit (version 2) in combination with an HiSeq Rapid SBS (version 2) sequencing kit (500-cycle PE or 150-cycle SE kit).
Population structure analysis
Genome sequencing reads were assembled using the velvet algorithm. MLST sequence types (ST) and cgMLST complex types (CT) according to the seven housekeeping gene MLST scheme [Citation37] and the 1701 locus cgMLST scheme [Citation7], respectively, were extracted from the assembled contigs by automated allele submission to the L. monocytogenes cgMLST server (http://www.cgmlst.org/ncs/schema/690488/). Clusters were defined as groups of strains with ≤10 different alleles between neighbouring strains. Generation of the minimal spanning tree was performed in the “pairwise, ignore missing values” mode. All of the aforementioned steps were performed using the built-in functions of the Ridom® SeqSphere Software package version 6.0.0 (2019/04, Ridom GmbH, Münster, Germany).
Antibiotic susceptibility testing
Antibiotic susceptibility testing was performed as a microdilution assay in accordance with the EUCAST guidelines in the January 2019 version [Citation38]. Briefly, selected L. monocytogenes strains were streaked out on BHI agar plates and incubated at 37°C for 24 h. Three to five colonies from each plate were picked, joined and further incubated in 3 mL BHI broth for 6 h. This culture was used to adjust NaCl solution (0.9%, w/w) to an OD600 of 0.005, representing a concentration of approximately 5·106 colony forming units (CFU) per mL. Ten µl of this solution were used to inoculate the individual wells of a 96-well microtiter plate containing 90 µl Mueller-Hinton fastidious (MH-F) broth with different concentrations of each individual tested antibiotic; 1 mM IPTG was added where necessary. The overall plate design was adopted from a study by Noll and colleagues [Citation26], produced in house and included ampicillin (AMP; MIC < 2 mg/L), benzylpenicillin (PEN; MIC < 2 mg/L), ceftriaxone (CRO; MIC < 4 mg/L), meropenem (MEP; MIC < 0.5 mg/L), daptomycin (DAP; MIC < 2 mg/L), ciprofloxacin (CIP; MIC < 2 mg/L), erythromycin (ERY; MIC < 2 mg/L), gentamicin (GEN; MIC < 2 mg/L), linezolid (LNZ; MIC < 8 mg/L), rifampicin (RAM; MIC < 1 mg/L), tetracycline (TET; MIC < 4 mg/L), tigecycline (TGC; MIC < 1 mg/L), vancomycin (VAN; MIC < 4 mg/L) and co-trimoxazole (SXT; MIC < 0.125 mg/L). Antibiotics were purchased form Sigma-Aldrich (St. Louis, USA), with the exception of LIN and DAP, which were purchased from Molekula GmbH (Munich, Germany). Their concentrations were selected to cover the EUCAST-defined MIC breakpoints [Citation38]. In cases where no breakpoint was defined for L. monocytogenes, the MIC breakpoints of Streptococcus pneumoniae or Staphylococcus aureus were used [Citation38]. The plates were quickly mixed and incubated in a sealed polyethylene bag at 37°C for 20 ± 2 h. Results were determined using a mirror for precise optical detection of growth. MICs were reported as the first concentration of the respective antibiotic where no visible growth was detected after the defined incubation period. Besides the Listeria monocytogenes reference strain EGD-e, a set of reference strains recommended by EUCAST guidelines (Escherichia coli ATCC 259226, Pseudomonas aeruginosa ATCC 278538, Staphylococcus aureus ATCC 292139 and Enterococcus faecalis ATCC 29212) with known antibiotic resistance profiles were used to assure effectivity of the antibiotics under the chosen testing conditions.
Statistical analysis
The Kruskal–Wallis rank sum test was performed to determine if there were significant differences between the determined MICs of the tested antibiotics between samples in serogroups IIa, IIb and IVb as well as between different sequence types (where ≥4 strains were available). To further test which groups significantly (p < 0.05) differed from one another, the pairwise Mann–Whitney-U test was performed. Adjusted p-values were obtained using a Bonferroni–Holm correction. All statistical analysis was performed using the stats package in R version 3.6.1 [Citation39].
Identification of alleles associated with reduced ceftriaxone resistance
Group-specific single nucleotide variations (SNV) were sought using the SNV tool implemented in SeqSphere (Ridom GmbH, Münster, Germany). For this purpose, isolates with reduced ceftriaxone resistance belonging to a particular ST were defined as target and isolates outside this phylogenetic group as non-target. Moreover, isolates belonging to one of the other low-ceftriaxone resistance STs were excluded from the non-target group to increase sensitivity. SNVs occurring in 100% of the target group and which were different to 99% of the non-target group were accepted and only SNVs leading to non-synonymous amino acid exchanges were considered for further analysis.
Results
Population structure-guided isolate selection
The collection of clinical L. monocytogenes strains from the German consultant laboratory was used as the source of genetic diversity within the L. monocytogenes population. At the time this project was started, the collection contained 1220 genome sequenced L. monocytogenes strains, isolated from human infections in Germany between 2009 and 2019. Of these strains 1004 had been isolated from blood or cerebrospinal fluid and the remaining strains from other sources. Therefore, the majority of the strains (82%) were associated with invasive disease. Most of the strains were collected in 2016 (n = 266), 2017 (n = 395) and 2018 (n = 453) (Figure S1).
The population structure of this strain collection was determined using MLST and cgMLST [Citation7,Citation12], allowing identification of disease clusters and sporadic cases. All strains belonged to phylogenetic lineage I (57%, n = 700) and lineage II (43%, n = 520); cgMLST grouped the 1220 isolates into 122 cgMLST complexes containing 798 isolates and 422 singletons. The 122 complexes varied in size from at least two up to 104 isolates, with a median size of 3 per complex (Figure S2). In order to cover all L. monocytogenes subtypes with current clinical relevance comprehensively, the following selection strategy was applied: At least one representative strain from each of the 122 identified complexes was selected. In cases where more than two genotypes belonged to a cluster, its most central isolate was chosen. If strains with different CTs formed a joined complex, a representative strain belonging to the most abundant CT within this complex was selected. An observation further justifying the selection of cluster representatives was obtained in a previous study, showing that isolates belonging to an outbreak cluster possess highly similar antibiotic resistance profiles [Citation40]. In addition to the cluster representatives, all sporadic isolates (422 of 1220) were included to further increase the genetic diversity within the selection of L. monocytogenes isolates. This procedure led to a selection of 544 L. monocytogenes strains from 2009 to 2019 with the majority of strains from 2016 to 2019 including representatives of the molecular serogroups IIa (39.7%), IIb (10.8%), IIc (1.3%), IVa (0.2%), IVb (46.7%), IVb-v1 (0.7%) and IVc (0.2%, Figure S1). Representatives of all 62 STs in the original strain collection were also present in this selection, with ST1, ST6 and ST2 representing the three most abundant STs (Figure S3). Of the 587 CTs identified in the original strain collection, 539 (92%) were also included. Thus, the strain selection for antibiotic profiling contained 544 L. monocytogenes isolates in total and represents a miniaturized model collection of the clinical L. monocytogenes population currently causing infections in Germany (Table S1, Figure S2).
Antibiotic profiling of the miniaturized model population
Each strain of the model population was tested for resistance against 14 clinically relevant antibiotics. No resistance was observed against the antibiotics currently recommended for the treatment of listeriosis; ampicillin, penicillin and co-trimoxazole (, (A)). Still, two of the tested strains were susceptible to increased concentrations (formerly described as intermediate resistance) of ampicillin and penicillin and three isolates were susceptible to increased concentrations of co-trimoxazole. Among all strains tested one showed resistance to gentamicin. No resistance was observed to erythromycin, linezolid, meropenem, rifampicin, tigecycline and vancomycin. Furthermore, all isolates tested (544/544, 100%) were resistant to ceftriaxone (). This observation is in full agreement with the intrinsic cephalosporin resistance of L. monocytogenes. Moreover, the majority of the screened strains (493/544, 91%) also showed resistance to daptomycin, a cyclic lipopeptide antibiotic. Around 13% of the isolates (71/544) showed resistance against the gyrase inhibitor ciprofloxacin. One strain was found to be resistant against tetracycline, an antibiotic to which most of the strains (518/544, 95%) showed intermediate resistance. Susceptibility to increased concentrations was also observed for most isolates in case of linezolid (515/544, 95%) and ciprofloxacin (451/544, 83%), while it was less common with vancomycin (203/544, 37%), gentamicin (55/544, 10%), daptomycin (45/544, 8%) and meropenem (17/544, 3%, ). Sixteen strains showed growth in the presence of 0.6125 mg/L rifampicin, the lowest tested concentration, and must thus be considered as susceptible to increased doses.
Figure 1. Identification of phylogenetic groups with reduced ceftriaxone resistance. (A) Phylogeny of isolates shown as Neighborhood-Joining tree based on the 1701 locus cgMLST scheme for the model population used for the antibiotic susceptibility testing. Starting from the centre, the rings represent the serogroups and the antibiotics tested (CRO, DAP, CIP, RAM, TET, LNZ, VAN, GEN, MEP, SXT, AMP, PEN, ERY, TGC). The colour code for the antibiotics represent resistant (red), intermediate susceptible (yellow) and susceptible (blue) strains for the individual antibiotics. The two outer rings show MIC values determined for CRO from 4 mg/L (green) to >128 mg/L (red), as well as the positions of the isolates belonging to the STs further investigated. Data was visualized using iTOL v4 [Citation41]. (B) Ceftriaxone resistance levels among 544 selected L. monocytogenes isolates according to MLST STs. Only STs for which MICs of ≥4 isolates were available were considered in this analysis.
![Figure 1. Identification of phylogenetic groups with reduced ceftriaxone resistance. (A) Phylogeny of isolates shown as Neighborhood-Joining tree based on the 1701 locus cgMLST scheme for the model population used for the antibiotic susceptibility testing. Starting from the centre, the rings represent the serogroups and the antibiotics tested (CRO, DAP, CIP, RAM, TET, LNZ, VAN, GEN, MEP, SXT, AMP, PEN, ERY, TGC). The colour code for the antibiotics represent resistant (red), intermediate susceptible (yellow) and susceptible (blue) strains for the individual antibiotics. The two outer rings show MIC values determined for CRO from 4 mg/L (green) to >128 mg/L (red), as well as the positions of the isolates belonging to the STs further investigated. Data was visualized using iTOL v4 [Citation41]. (B) Ceftriaxone resistance levels among 544 selected L. monocytogenes isolates according to MLST STs. Only STs for which MICs of ≥4 isolates were available were considered in this analysis.](/cms/asset/291fbb8f-f7e0-4729-8247-7548d5524b11/temi_a_1799722_f0001_oc.jpg)
Table 1. Antibiotic resistance profiles of the L. monocytogenes model population.
The most common co-occurrence of antibiotic resistance was observed with ceftriaxone in addition to daptomycin (493/544, 91%). Out of these, 66 strains (12%) showed additional resistance to ciprofloxacin. Only two isolates were found to be resistant to ceftriaxone and ciprofloxacin while being susceptible to daptomycin. Forty-five isolates (8%) were only resistant to ceftriaxone but none of the other antibiotics tested. Thus, they only showed intrinsic resistance against cephalosporins.
Identification of phylogenetic groups with different antibiotic resistance profiles
The majority of isolates within the model population belonged to the molecular serogroups IIa, IIb and IVb (529 of 544, 97%). On the binary observation level of resistant versus sensitive, resistances were equally distributed between these three main molecular serogroups. To increase the resolution, the MIC values for each antibiotic were compared between isolates belonging to the different molecular serogroups. The average MICs for ampicillin (IVb = 0.36 mg/L, IIa = 0.18 mg/L), penicillin (IVb = 0.38 mg/L, IIa = 0.19 mg/L), daptomycin (IVb = 2.94 mg/L, IIa = 2.46 mg/L), linezolid (IVb = 3.78 mg/L, IIa = 2.13 mg/L), tetracycline (IVb = 1.49 mg/L, IIa = 1.11 mg/L), tigecycline (IVb = 0.12 mg/L, IIa = 0.08 mg/L) were significantly higher (Mann–Whitney U Test, n1 = 216, n2 = 254, P < 0.05) for serogroup IVb isolates compared to isolates of serogroup IIa (Figure S4).
Despite this observation, we also found that the MICs for ceftriaxone varied between 4 mg/L up to >128 mg/L, with a median MIC of >128 mg/L considering all tested isolates (). While this classifies all strains as ceftriaxone-resistant, reduced median MIC values for ceftriaxone of ≤32 mg/L were found for certain STs ((B)). The largest phylogenetic group with lowered ceftriaxone resistance was ST4 (n = 24 isolates), showing a reduced median MIC of 32 mg/L in contrast to >128 mg/L for the remaining population. Likewise, lowered ceftriaxone MICs were observed for ST29 (median MIC = 24 mg/L, n = 7), ST388 (median MIC = 24 mg/L, n = 4) and ST403 isolates (median MIC = 16 mg/L, n = 8, (B)).
Reduced ceftriaxone resistance levels were also observed in ST7 (median MIC = 64 mg/L), ST9 (median MIC = 64 mg/L), ST101 (median MIC = 128 mg/L) and ST204 isolates (median MIC = 64 mg/L, (B)).
Identifying pbpB3 alleles linked to reduced ceftriaxone resistance
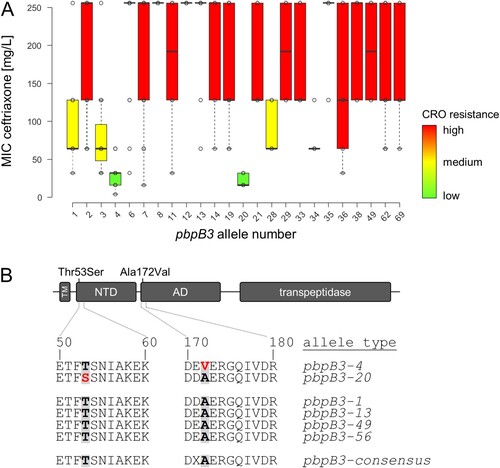
Single nucleotide variant analysis revealed that ST4, ST29, ST388 and ST403 isolates associated with lowered levels of ceftriaxone resistance carried group-specific non-synonymous mutations in various coding regions. However, the only gene carrying one mutation common to all isolates belonging to the STs with reduced ceftriaxone resistance was lmo0441, encoding PBP B3, which showed a mutation within the allelic version found in ST4 and ST388 (pbpB3 allele type 4, Ala172Val) and ST403 and ST29 (pbpB3 allele type 20, Thr53Ser, (A,B)). This suggests that certain pbpB3 alleles are associated with reduced resistance against ceftriaxone. Remarkably, all ST4 and ST388 isolates carried the pbpB3 Ala172Val substitution characteristic for pbpB3 allele no. 4 in the Ruppitsch cgMLST scheme and this pbpB3 allele was not found in any other strain. Likewise, all our ST403 isolates carried the pbpB3 Thr53Ser variant (allele no. 20), also found in four out of six ST29 isolates tested with lowered ceftriaxone resistance levels. The two ST29 isolates tested with a ceftriaxone resistance above the median value observed in this group had a different pbpB3 allele. Despite its presence in these two subgroups, pbpB3 allele no. 20 was not found in any other of the 1220 strains of the original strain collection. We thus conclude that pbpB3 alleles 4 and 20 are associated with reduced ceftriaxone resistance.
Figure 2. Identification of pbpB3 alleles associated with reduced ceftriaxone resistance. (A) Ceftriaxone resistance levels among 544 selected L. monocytogenes isolates according to their pbpB3 allele in the Ruppitsch cgMLST scheme. Only those pbpB3 alleles for which MICs of ≥4 isolates were available were considered in this analysis. (B) Scheme illustrating PBP B3 domains and position of the amino acid exchanges found in the pbpB3 alleles no. 4 (Ala172Val) and 20 (Thr53Ser), which are associated with reduced ceftriaxone resistance. Abbreviations: TM – transmembrane helix, NTD – N-terminal domain; AD – allosteric domain.
Effect of novel pbpB3 mutations on ceftriaxone resistance
Even though the sequence alterations in the two pbpB3 alleles were rather conservative at the protein level, their contribution to ceftriaxone resistance was tested in a complementation assay. For this purpose, a ΔpbpB3 deletion mutant constructed in the background of L. monocytogenes EGD-e (strain LMJR41) [Citation32] was complemented with different pbpB3 alleles and ceftriaxone resistance of the resulting strains was determined. Ceftriaxone resistance was greatly reduced in the Δlmo0441 mutant (2 mg/L) compared to wild type strain EGD-e (64 mg/L, ). Reintroduction of the wild type pbpB3 allele (allele type 1) from EGD-e restored this phenotype almost completely (32 mg/L). In contrast, expression of pbpB3 allele type 4 associated with reduced ceftriaxone resistance in the ΔpbpB3 background led to a lower ceftriaxone resistance level of only 16 mg/L (). When pbpB3 allele type 49, originating from a closely related but fully ceftriaxone-resistant ST217 isolate (MIC >128 mg/L, n = 6), was expressed in the ΔpbpB3 background, ceftriaxone resistance increased to 32 mg/L. This level of ceftriaxone resistance further increased to 64 mg/L, when pbpB3 allele type 13 from ST6 strain 18-04540, showing the highest observed level of ceftriaxone resistance in this study, was used for complementation (). The complementation of the deletion mutant with pbpB3 allele type 56 increased the ceftriaxone MIC to 32 mg/L (). This allele type is identical to pbpB3 allele type 4 except for a single mutation at the aforementioned position 172, where it still carries the original alanine. These results further underline the apparent importance of this single amino acid for the resistance against ceftriaxone. As for the pbpB3 allele type 4, complementation mutants carrying pbpB3 allele type 20 showed higher ceftriaxone compared to the deletion mutant but lower ceftriaxone resistance compared to the complementation mutants carrying pbpB3 alleles of the wild type strain or from the high level resistance strain (). In conclusion, pbpB3 alleles from strains with low and high levels of ceftriaxone resistance confer low and high levels of ceftriaxone resistance upon their heterologous expression in the ΔpbpB3 mutant, respectively. This confirms the association of certain pbpB3 alleles with ceftriaxone resistance and demonstrates the population-wide validity of the concept that PBP B3 is an important determinant for ceftriaxone resistance in L. monocytogenes.
Table 2. Effect of pbpB3 on ceftriaxone resistance.
To estimate the overall relevance of this observation for the entire L. monocytogenes population, the frequency of pbpB3 allele types 4 and 20 was calculated for the model population of 544 strains (55 unique pbpB3 allele types), for the initially used clinical strain collection of 1220 strains (58 unique pbpB3 allele types) as well as for 27,118 L. monocytogenes genomes available on the National Center for Biotechnology Information (NCBI) pathogen detection pipeline at the time of this study (1033 unique pbpB3 allele types). Allele type 4 was detected in 28 strains of the model collection (expected: 10), in 39 strains of the clinical strain collection (expected: 21) and 340 times in the NCBI dataset (expected: 26). Allele type 20 was detected in 12 strains of the model collection, 62 strains of the clinical strain collection and 156 strains of the NCBI dataset. Therefore, the abundance of both allele types was above the theoretically expected values and hence the presence of theses pbpB3 allele types does not seem to provide an evolutionary disadvantage.
Discussion
Our results represent the first comprehensive determination of antibiotic resistance patterns of clinical L. monocytogenes strains isolated in Germany. The complexity of this strain collection was reduced by the generation of a non-redundant model population using cgMLST subtyping data. This model population contains less than half of the original isolates but still maintains the large biodiversity observed in the original L. monocytogenes clinical strain collection; determination of antibiotic resistance patterns in this model population greatly facilitated experimental determination of antibiotic resistance patterns without losing phylogenetic resolution.
An important finding of this study is the sustained effectivity of the standard antibiotics recommended for the treatment of listeriosis. None of the L. monocytogenes strains tested here showed full resistance against ampicillin and penicillin and only one was resistant towards gentamicin. However, gentamicin is not used as a stand-alone antibiotic in listeriosis therapy and only administered in combination with ampicillin or penicillin. Moreover, none of the isolates tested showed full resistance against co-trimoxazole, which is used as an alternative in patients with β-lactam allergy. However, susceptibility only to increased concentrations of penicillin (2/544), ampicillin (2/544) and co-trimoxazole (3/544) was observed in few cases. Therefore our results are in accordance with observations made with other clinical strain collections from Europe where intermediate resistance levels against these three antibiotics were also reported to occur with low frequency [Citation23,Citation27].
The highest level of resistance within our model population was observed for ceftriaxone (100%), to which L. monocytogenes is intrinsically resistant [Citation14,Citation28], daptomycin (91%) and ciprofloxacin (13%). However, breakpoints have not been established for daptomycin and ciprofloxacin in L. monocytogenes (as none of them is recommended to treat listeriosis) and applications of cephalosporins and ciprofloxacin have caused therapy failure in the past [Citation42–44].
A large variation of ceftriaxone MICs ranging from 4 mg/L up to >128 mg/L was observed between isolates belonging to different STs and could be traced back to amino acid exchanges in pbpB3. Interestingly, an almost similar degree of variation in ceftriaxone resistance was observed within the ST1, ST155, ST451 strains included here ((B)), even though no association between ceftriaxone resistance and pbpB3 allele variation was found in these STs. Cephalosporin resistance is a multifactorial process in L. monocytogenes [Citation31], and genetic variations in other cephalosporin resistance determinants, such as other PBPs, certain transporters or regulators [Citation31,Citation45], may account for the variability of ceftriaxone resistance in these phylogenetic groups.
PBP B3 of L. monocytogenes belongs to the same subclass of class B PBPs as Bacillus subtilis PBP3, Staphylococcus aureus PBP2a (encoded by mecA) and Enterococcus faecalis PBP5, which all are low-affinity penicillin binding proteins and as such critical determinants of cephalosporin or methicillin resistance in these bacteria [Citation46–49]. The two pbpB3 mutations lowering cephalosporin resistance described here affect the N-terminal domain and the allosteric domain (non-penicillin binding domain) of PBP B3 ((B)). The function of these non-catalytic domains is not entirely clear, but amino acid exchanges in the allosteric domain of S. aureus PBP2a (such as N146 K and E150 K) are associated with increased resistance of S. aureus to ceftaroline, a fifth-generation cephalosporin [Citation50–53]. Ceftaroline non-covalently interacts with this allosteric domain inducing a conformational change that makes the active site in the transpeptidase domain accessible for acylation and thus for inhibition by a second ceftaroline molecule [Citation54]. The N146 K and E150 K mutations of S. aureus PBP2a map to the same stretch in the beginning of the allosteric domain as the A172 V exchange in PBP B3 of L. monocytogenes. Apparently, amino acid exchanges in this region of the allosteric domain improve or impair cephalosporin binding in low affinity PBPs and thus resistance of different Gram-positive pathogens to this important group of antibiotics.
While the low level of resistance towards currently clinically applied antibiotics is a relief, the situation in environmental and food isolates is more alarming. L. monocytogenes strains with multidrug resistance or resistance to ampicillin, penicillin or co-trimoxazole have repeatedly been isolated from the environment and from different food types [Citation24,Citation26,Citation55–60]. It can be expected that the antibiotic resistances observed in environmental and food strains today will later manifest in clinical strains. Therefore, surveillance of antimicrobial resistance development in clinical L. monocytogenes strains in the future is of great importance, especially since average resistance levels against several β-lactams have been continuously increasing since the 1920s in clinical L. monocytogenes isolates from France [Citation27].
Fischer_et_al._Table_S1_-_Antibiotic_susceptibilities_of_the_L._monocytogenes_model_population.xlsx
Download ()Fischer_et_al._supplementary_material_finanl.doc
Download ()Acknowledgements
The authors would like to acknowledge Matthias Noll and Ingo Klare for fruitful discussions, Andrea Thürmer for genome sequencing and Karsten Großhennig, Petra Hahs and Claudia Lampel for technical assistance.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Buchanan RL, Gorris LGM, Hayman MM, et al. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control. 2017;75:1–13.
- Robert-Koch-Institut. Infektionsepidemiologisches Jahrbuch meldepflichtiger Krankheiten für 2018; 2019.
- de Noordhout CM, Devleesschauwer B, Angulo FJ, et al. The global burden of listeriosis: a systematic review and meta-analysis. Lancet Infect Dis. 2014;14(11):1073–1082.
- Koopmans MM, Bijlsma MW, Brouwer MC, et al. Listeria monocytogenes meningitis in the Netherlands, 1985–2014: A nationwide surveillance study. J Infect. 2017;75(1):12–19.
- Charlier C, Perrodeau E, Leclercq A, et al. Clinical features and prognostic factors of listeriosis: the MONALISA national prospective cohort study. Lancet Infect Dis. 2017;17(5):510–519.
- Goulet V, King LA, Vaillant V, et al. What is the incubation period for listeriosis? BMC Infect Dis. 2013;13:11.
- Ruppitsch W, Pietzka A, Prior K, et al. Defining and evaluating a core genome multilocus sequence typing scheme for whole-genome sequence-based typing of Listeria monocytogenes. J Clin Microbiol. 2015;53(9):2869–2876.
- Kwong JC, Mercoulia K, Tomita T, et al. Prospective whole-genome sequencing enhances national surveillance of Listeria monocytogenes. J Clin Microbiol. 2016;54(2):333–342.
- Chen Y, Gonzalez-Escalona N, Hammack TS, et al. Core genome multilocus sequence typing for identification of globally distributed clonal groups and differentiation of outbreak strains of Listeria monocytogenes. Appl Environ Microbiol. 2016;82(20):6258–6272.
- Kleta S, Hammerl JA, Dieckmann R, et al. Molecular tracing to find source of protracted invasive listeriosis outbreak, Southern Germany, 2012–2016. Emerging Infect Dis. 2017;23(10):1680–1683.
- Moura A, Tourdjman M, Leclercq A, et al. Real-time whole-genome sequencing for surveillance of Listeria monocytogenes, France. Emerging Infect Dis. 2017;23(9):1462–1470.
- Halbedel S, Prager R, Fuchs S, et al. Whole-genome sequencing of recent Listeria monocytogenes isolates from Germany reveals population structure and disease clusters. J Clin Microbiol. 2018;56(6):e00119.
- Moellering Jr., RC, Medoff G, Leech I, et al. Antibiotic synergism against Listeria monocytogenes. Antimicrob Agents Chemother. 1972;1(1):30–34.
- Hof H. Listeriosis: therapeutic options. FEMS Immunol Med Microbiol. 2003;35(3):203–205.
- Mitja O, Pigrau C, Ruiz I, et al. Predictors of mortality and impact of aminoglycosides on outcome in listeriosis in a retrospective cohort study. J Antimicrob Chemother. 2009;64(2):416–423.
- Munoz P, Rojas L, Bunsow E, et al. Listeriosis: An emerging public health problem especially among the elderly. J Infect. 2012;64(1):19–33.
- Koopmans MM, Engelen-Lee J, Brouwer MC, et al. Characterization of a Listeria monocytogenes meningitis mouse model. J Neuroinflammation. 2018;15(1):257.
- Grant MH, Ravreby H, Lorber B. Cure of Listeria monocytogenes meningitis after early transition to oral therapy. Antimicrob Agents Chemother. 2010;54(5):2276–2277.
- Stepanovic S, Lazarevic G, Jesic M, et al. Meropenem therapy failure in Listeria monocytogenes infection. Eur J Clin Microbiol Infect Dis. 2004;23(6):484–486.
- Thonnings S, Knudsen JD, Schonheyder HC, et al. Antibiotic treatment and mortality in patients with Listeria monocytogenes meningitis or bacteraemia. Clin Microbiol Infect. 2016;22(8):725–730.
- Su X, Zhang J, Shi W, et al. Molecular characterization and antimicrobial susceptibility of Listeria monocytogenes isolated from foods and humans. Food Control. 2016;70:96–102.
- Althaus D, Lehner A, Brisse S, et al. Characterization of Listeria monocytogenes strains isolated during 2011–2013 from human infections in Switzerland. Foodborne Pathog Dis. 2014;11(10):753–758.
- Kuch A, Goc A, Belkiewicz K, et al. Molecular diversity and antimicrobial susceptibility of Listeria monocytogenes isolates from invasive infections in Poland (1997–2013). Sci Rep. 2018;8(1):14562.
- Conter M, Paludi D, Zanardi E, et al. Characterization of antimicrobial resistance of foodborne Listeria monocytogenes. Int J Food Microbiol. 2009;128(3):497–500.
- Harakeh S, Saleh I, Zouhairi O, et al. Antimicrobial resistance of Listeria monocytogenes isolated from dairy-based food products. Sci Total Environ. 2009;407(13):4022–4027.
- Noll M, Kleta S, Al Dahouk S. Antibiotic susceptibility of 259 Listeria monocytogenes strains isolated from food, food-processing plants and human samples in Germany. J Infect Public Health. 2018;11(4):572–577.
- Morvan A, Moubareck C, Leclercq A, et al. Antimicrobial resistance of Listeria monocytogenes strains isolated from humans in France. Antimicrob Agents Chemother. 2010;54(6):2728–2731.
- Hof H, Nichterlein T, Kretschmar M. Management of listeriosis. Clin Microbiol Rev. 1997;10(2):345–357.
- El Bashir H, Laundy M, Booy R. Diagnosis and treatment of bacterial meningitis. Arch Dis Child. 2003;88(7):615–620.
- Guinane CM, Cotter PD, Ross RP, et al. Contribution of penicillin-binding protein homologs to antibiotic resistance, cell morphology, and virulence of Listeria monocytogenes EGDe. Antimicrob Agents Chemother. 2006;50(8):2824–2828.
- Krawczyk-Balska A, Markiewicz Z. The intrinsic cephalosporin resistome of Listeria monocytogenes in the context of stress response, gene regulation, pathogenesis and therapeutics. J Appl Microbiol. 2016;120(2):251–265.
- Rismondo J, Möller L, Aldridge C, et al. Discrete and overlapping functions of peptidoglycan synthases in growth, cell division and virulence of Listeria monocytogenes. Mol Microbiol. 2015;95(2):332–351.
- Doumith M, Buchrieser C, Glaser P, et al. Differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J Clin Microbiol. 2004;42(8):3819–3822.
- Kerouanton A, Marault M, Petit L, et al. Evaluation of a multiplex PCR assay as an alternative method for Listeria monocytogenes serotyping. J Microbiol Methods. 2010;80(2):134–137.
- Leclercq A, Chenal-Francisque V, Dieye H, et al. Characterization of the novel Listeria monocytogenes PCR serogrouping profile IVb-v1. Int J Food Microbiol. 2011;147(1):74–77.
- Monk IR, Gahan CG, Hill C. Tools for functional postgenomic analysis of Listeria monocytogenes. Appl Environ Microbiol. 2008;74(13):3921–3934.
- Ragon M, Wirth T, Hollandt F, et al. A new perspective on Listeria monocytogenes evolution. PLoS Pathog. 2008;4(9):e1000146.
- EUCAST. Breakpoint tables for interpretation of MICs and zone diameters, Version 9.0 European Committee on Antimicrobial Susceptibility Testing; 2019 [updated 01.01.2019; cited 2019 05.12.2019]. Available from: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_9.0_Breakpoint_Tables.pdf.
- Team R. A language and environment for statistical computing. Computing. 2006 01/01;1.
- Halbedel S, Wilking H, Holzer A, et al. Large nationwide outbreak of invasive listeriosis associated with blood sausage, Germany, 2018–2019. Emerging Infect Dis. 2020;26(7):1456–1464.
- Letunic I, Bork P. Interactive tree of life (iTOL) v4: recent updates and new developments. Nucleic Acids Res. 2019;47(W1):W256–W259.
- Lorber B, Santoro J, Swenson RM. Letter: Listeria meningitis during cefazolin therapy. Ann Intern Med. 1975;82(2):226.
- Kawaler B, Hof H. Failure of cephalosporins to cure experimental listeriosis. J Infect. 1984;9(3):239–243.
- Grumbach NM, Mylonakis E, Wing EJ. Development of listerial meningitis during ciprofloxacin treatment. Clin Infect Dis. 1999;29(5):1340–1341.
- Wamp S, Rutter ZJ, Rismondo J, et al. PrkA controls peptidoglycan biosynthesis through the essential phosphorylation of ReoM. eLife. 2020;9:e56048.
- Chambers HF. Methicillin-resistant staphylococci. Clin Microbiol Rev. 1988;1(2):173–186.
- Arbeloa A, Segal H, Hugonnet JE, et al. Role of class A penicillin-binding proteins in PBP5-mediated beta-lactam resistance in Enterococcus faecalis. J Bacteriol. 2004;186(5):1221–1228.
- Sauvage E, Kerff F, Terrak M, et al. The penicillin-binding proteins: structure and role in peptidoglycan biosynthesis. FEMS Microbiol Rev. 2008;32(2):234–258.
- Sassine J, Xu M, Sidiq KR, et al. Functional redundancy of division specific penicillin-binding proteins in Bacillus subtilis. Mol Microbiol. 2017;106(2):304–318.
- Mendes RE, Tsakris A, Sader HS, et al. Characterization of methicillin-resistant Staphylococcus aureus displaying increased MICs of ceftaroline. J Antimicrob Chemother. 2012;67(6):1321–1324.
- Alm RA, McLaughlin RE, Kos VN, et al. Analysis of Staphylococcus aureus clinical isolates with reduced susceptibility to ceftaroline: an epidemiological and structural perspective. J Antimicrob Chemother. 2014;69(8):2065–2075.
- Kelley WL, Jousselin A, Barras C, et al. Missense mutations in PBP2A Affecting ceftaroline susceptibility detected in epidemic hospital-acquired methicillin-resistant Staphylococcus aureus clonotypes ST228 and ST247 in Western Switzerland archived since 1998. Antimicrob Agents Chemother. 2015;59(4):1922–1930.
- Bongiorno D, Mongelli G, Stefani S, et al. Genotypic analysis of Italian MRSA strains exhibiting low-level ceftaroline and ceftobiprole resistance. Diagn Microbiol Infect Dis. 2019;95(3):114852.
- Otero LH, Rojas-Altuve A, Llarrull LI, et al. How allosteric control of Staphylococcus aureus penicillin binding protein 2a enables methicillin resistance and physiological function. Proc Natl Acad Sci U S A. 2013;110(42):16808–16813.
- Srinivasan V, Nam HM, Nguyen LT, et al. Prevalence of antimicrobial resistance genes in Listeria monocytogenes isolated from dairy farms. Foodborne Pathog Dis. 2005;2(3):201–211.
- Jamali H, Paydar M, Ismail S, et al. Prevalence, antimicrobial susceptibility and virulotyping of Listeria species and Listeria monocytogenes isolated from open-air fish markets. BMC Microbiol. 2015;15:144.
- Abdollahzadeh E, Ojagh SM, Hosseini H, et al. Antimicrobial resistance of Listeria monocytogenes isolated from seafood and humans in Iran. Microb Pathog. 2016;100:70–74.
- Li L, Olsen RH, Ye L, et al. Characterization of antimicrobial resistance of Listeria monocytogenes strains isolated from a pork processing plant and its respective meat markets in Southern China. Foodborne Pathog Dis. 2016;13(5):262–268.
- Sala C, Morar A, Tirziu E, et al. Environmental occurrence and antibiotic susceptibility profile of Listeria monocytogenes at a slaughterhouse raw processing plant in Romania. J Food Prot. 2016;79(10):1794–1797.
- Roedel A, Dieckmann R, Brendebach H, et al. Biocide-Tolerant Listeria monocytogenes isolates from German food production plants do not show cross-resistance to clinically relevant antibiotics. Appl Environ Microbiol. 2019;85(20):e01253-19.