
ABSTRACT
Distinct SARS-CoV-2 Omicron sublineages have evolved showing increased fitness and immune evasion than the original Omicron variant BA.1. Here, we report the neutralization activity of sera from BNT162b2 vaccinated individuals or unimmunized Omicron BA.1-infected individuals against Omicron sublineages and “Deltacron” variant (XD). BNT162b2 post-dose 3 immune sera neutralized USA-WA1/2020, Omicron BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA.4/5-, and XD-spike SARS-CoV-2s with geometric mean titres (GMTs) of 1335, 393, 298, 315, 216, 103, and 301, respectively; thus, BA.4/5 SARS-CoV-2 spike variant showed the highest propensity to evade vaccine neutralization compared to the original Omicron variants BA.1. BA.1-convalescent sera neutralized USA-WA1/2020, BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA.4/5-, and Deltacron-spike SARS-CoV-2s with GMTs of 15, 430, 110, 109, 102, 25, and 284, respectively. The unique mutation F486V in the BA.4/5 spike contributes to the increased evasion of antibody neutralization by sublineage BA.4/5. The low neutralization titres of vaccinated sera or convalescent sera from BA.1 infected individuals against the emerging and rapidly spreading Omicron BA.4/5 variants provide important results for consideration in the selection of an updated vaccine in the current Omicron wave.
Trial registration: ClinicalTrials.gov; identifier: NCT04368728.
Main text
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) Omicron variant has emerged as the fifth variant of concern (VOC) after Alpha, Beta, Gamma, and Delta. Omicron exhibited the greatest evasion from vaccine- and infection-elicited neutralizing antibodies among all the VOCs [Citation1,Citation2]. Since the emergence of Omicron in late November 2021, distinct sublineages have evolved around the world: BA.1 was responsible for the initial surge and has now been replaced by BA.2 and BA.2.12.1 in the USA (https://covid.cdc.gov/covid-data-tracker/#variant-proportions); BA.4 and BA.5 have become prevalent in South Africa and, more recently, are becoming prevalent in some areas in Europe and are increasing in the USA; BA.3 currently remains at low frequency. The co-circulation of Omicron and Delta has also led to a hybrid variant “Deltacron” [Citation3]. Since these newly emerged Omicron sublineages and Deltacron have distinct mutations in the spike glycoproteins (Figure S1(A)), it is important to examine their susceptibility to vaccine- or infection-elicited antibody neutralization.
To assess the neutralization titres of sera from BNT162b2-immunized or Omicron BA.1-infected individuals against different variants, we engineered the complete spike gene from Omicron sublineages BA.1 (GISAID EPI_ISL_6640916), BA.2 (GISAID EPI_ISL_6795834.2), BA.2.12.1 (GISAID EPI_ISL_12115772), BA.3 (GISAID EPI_ISL_7605591), BA.4/5 (BA.4: GISAID EPI_ISL_11542270; BA.5: GISAID EPI_ISL_11542604; BA.4 and BA.5 have the identical spike sequence), and Deltacron XD strain (GISAID EPI_ISL_10819657 with the first 158 amino acids from Delta spike and the rest from Omicron BA.1 spike) into the mNeonGreen (mNG) reporter USA-WA1/2020 strain, a SARS-CoV-2 strain isolated in January 2020 (Figure S1(A)). The resulting BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA.4/5-, and XD-spike mNG SARS-CoV-2s produced infectious titres in Vero E6 cells of >106 plaque-forming units per millilitre (PFU/ml), similar to the wild-type USA-WA1/2020 mNG virus. The recombinant viruses developed decreasing sizes of fluorescent foci in the order of USA-WA1/2020 > BA.4/5-spike > BA.2-spike ≈ BA.2.12.1-spike ≈ BA.3-spike > XD-spike ≈ BA.1-spike mNG SARS-CoV-2 (Figure S1(B)). All recombinant viruses were sequenced to ensure no undesired mutations. Only the sequenced passage 1 viruses were used for the neutralization assays.
Using the recombinant SARS-CoV-2s, we determined the 50% fluorescent focus-reduction neutralization titres (FFRNT50) for two panels of human sera. The first serum panel (n = 22) was collected from individuals at one-month post-dose 3 (PD3) of BNT162b2 vaccine. We chose the PD3 sera because (i) many people have already received 3 doses of BNT162b2 and (ii) two doses of BNT162b2 did not elicit robust neutralization against Omicron BA.1 [Citation2]. The second serum panel (n = 20) was collected from unvaccinated COVID-19 patients who had been infected by BA.1 during the initial Omicron emergence in the USA [Citation4]. The genotype of the infecting virus was verified for each patient by Sanger sequencing of nasal swabs. Tables S1 and S2 summarize the data from the PD3 BNT162b2-vaccinated- and BA.1-infected individuals, respectively.
BNT162b2 PD3 immune sera neutralized USA-WA1/2020, BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA.4/5-, and XD-spike mNG SARS-CoV-2s with geometric mean titres (GMTs) of 1335, 393, 298, 315, 216, 103, and 301, respectively ((A) and Table S1). All PD3 sera neutralized all variant-spike SARS-CoV-2s with neutralization titres of >20. However, neutralizing GMTs against BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA.4/5-, and XD-spike SARS-CoV-2s were 3.4-, 4.5-, 4.2-, 6.2-, 13.0-, and 4.4-fold lower than that against USA-WA1/2020 GMT, respectively ((A)). The results support two conclusions. First, BA.1, BA.2, BA.2.12.1, and XD-spike variants have an increased but similar propensity to evade neutralizing antibodies induced by three doses of BNT162b2 and likely other COVID-19 vaccines. This result suggests that the sequential increase in the prevalence of BA.2.12.1 > BA.2 > BA.1 over the past six months was likely not driven by the difference in spike-mediated evasion of neutralization in vaccinated people, but by other factors, such as differences in viral transmission, or other mechanisms of immune evasion [Citation5,Citation6]. Second, BA.4/5 variants are much less efficiently neutralized by BNT162b2 PD3 immune sera than BA.1, BA.2, BA.2.12.1, BA.3, and XD spikes. The BA.3 sublineage may be attenuated in viral fitness as it has remained at very low prevalence; however, BA.4 and BA.5 sublineages, with the additional F486V and reversion of Q493 (compared to BA.2.12.1) that appear to improve infectivity, may potentially replace other Omicron sublineages in circulation. Indeed, reversion of the V486 mutation to the wild-type F486 restored the susceptibility of BA.4 V486F-spike virus to neutralization (Figure S2 and Table S3). These findings suggest that (i) close monitoring of the prevalence of sublineages BA.4 and BA.5 through epidemiological surveillance is critical as rapid increases in these sublineages with higher levels of immune escape could lead to new waves of infections and (ii) the unique mutation F486V in the BA.4/5 spike contributes to the increased evasion of antibody neutralization by sublineage BA.4/5.
Figure 1. Neutralization by sera collected at one-month post-dose 3 BNT162b2 vaccine (A) and by sera collected from unvaccinated individuals who contracted Omicron BA.1 SARS-CoV-2 (B). Scatterplot of neutralization titres against USA-WA1/2020, Omicron sublineage BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA.4/5-, and Deltacron XD-spike mNG SARS-CoV-2s. Both BNT162b2-vaccinated sera (n = 22) and BA.1-infected convalescent sera (n = 20) were tested for their FFRNT50s against the variant-spike mNG SARS-CoV-2s. The variant-spike mNG SARS-CoV-2s were produced by engineering the complete variant-spike genes into the mNG USA-WA1/2020. Each data point represents the geometric mean FFRNT50 (GMT) obtained with a serum specimen against the indicated virus. Tables S1 and S2 summarize the serum information and FFRNT50s for (A) and (B), respectively. The neutralization titres for individual variant-spike mNG SARS-CoV-2s were determined in two or three independent experiments, each with duplicate assays; the GMTs are presented. The bar heights and the numbers above indicate GMTs. The whiskers indicate 95% confidence intervals. The dotted lines indicate the limit of detection of FFRNT50. Statistical analysis was performed with the use of the Wilcoxon matched-pairs signed-rank test. For the BNT162b2-vaccinated sera in (A), the P-values of the GMT differences between USA-WA1/2020 and any variant-spike SARS-CoV-2 are all <.0001. For the BA.1-convelescent sera in (B), the P-value of GMT difference between BA.1- and XD-spike viruses is .0021; the P-values of the GMT differences between BA.1- and any other variant-spike viruses (including USA-WA1/2020) are all <.0001. For both serum panels in (A) and (B), FFRNT50 values with connected lines are presented for individual sera.
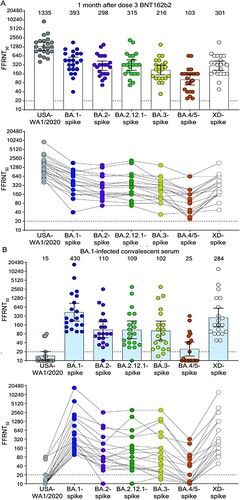
Sera from BA.1-infected individuals neutralized USA-WA1/2020, BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA.4/5-, and XD-spike viruses with GMTs of 15, 430, 110, 109, 102, 25, and 284, respectively ((B) and Table S2). Thus, the neutralizing GMTs against heterologous USA-WA1/2020-, BA.2-, BA.2.12.1-, BA.3-, BA.4/5-, and XD-spike SARS-CoV-2s were 28.7-, 3.9-, 3.9-, 4.2-, 17.2-, and 1.5-fold lower than that against homologous BA.1-spike virus, respectively. All sera neutralized BA.1-spike virus with titres of ≥80. Importantly, however, 13 (56%), 1 (5%), 1 (5%), and 10 (50%) out of 20 sera did not neutralize USA-WA1/2020, BA.2-, BA.2.12.1-, and BA.4/5-spike SARS-CoV-2s, respectively ((B)). These results suggest that sera from BA.1-infected individuals do not neutralize all Omicron sublineages and Deltacron XD with similar efficiencies, and importantly are not efficient at neutralizing BA.4/5. As shown in Figure S2, the additional mutation F486V in the BA.4/5 spike may account for the further evasion of neutralization.
The current study has confirmed and expanded our previous results on Omicron sublineage neutralization [Citation1,Citation2,Citation4]. Our results have potential implications for the strategy of updating the current COVID-19 vaccines in which, even within the Omicron lineages, substantial differences exist in the potential of vaccinated or BA.1-infected individuals to neutralize Omicron sublineages. The data suggest that Omicron BA.4 and BA.5 sublineages are adapted to evade neutralization in both prototype vaccinated and Omicron BA.1-infected individuals. The antigenic distinctions of spike glycoproteins among Omicron sublineages, Deltacron XD, and USA-WA1/2020 as well as prevalence of Omicron sublineages must be carefully considered when deciding to update the vaccine to new variants.
Our study has some limitations. This study lacks analyses of T cells and non-neutralizing antibodies that can mediate Fc-mediated effector functions. Both of these immune arms, in conjunction with neutralizing antibodies, protect patients from severe disease. After vaccination or infection, the majority of the T cell epitopes are preserved in Omicron spikes [Citation7]. Caution should also be taken when comparing the differences in variant neutralization between BNT162b2-vaccinated (PD3 sera) and BA.1-infected individuals. This is because (i) a third dose booster vaccination increases both the magnitude and breadth of neutralization [Citation8], which might explain the observation that the neutralizing GMTs of BA.1 infected convalescent sera against heterologous variants or sublineages were reduced more than those from BNT162b2 PD3 sera. (ii) BA.1-infected sera used in this study (collected on days 8–62 after a positive RT–PCR test) were heterogeneous, with some sera collected at an acute plasma blast stage and other sera at a convalescent IgG dominant phase; and (iii) the relatively small sample size. Regardless, our results consistently showed the most significant reduction in neutralization against BA.4/5 spike, underscoring the importance of closely monitoring the prevalence of sublineages BA.4 and BA.5 globally and taking the data into account when making decisions on updating current COVID-19 vaccines.
Methods
Human sera
Two human serum panels were used in the study. The first panel of serum samples was collected from BNT162b2 vaccines participating in the phase 1 portion of the ongoing phase 1/2/3 clinical trial (ClinicalTrials.gov identifier: NCT04368728). The protocol and informed consent were approved by institutional review boards for each of the investigational centres participating in the study. The study was conducted in compliance with all International Council for Harmonisation Good Clinical Practice guidelines and the ethical principles of the Declaration of Helsinki. The primary outcomes for phase 1 were reported previously [Citation1,Citation9,Citation10]. BNT162b2-vaccinated sera (n = 22) were collected one month after dose 3 and used for neutralization test. Table S1 summarizes the patient information which was previously reported [Citation8,Citation11].
The second serum panel (n = 20) was collected from COVID-19 patients who were not vaccinated but infected by BA.1 sublineage during the initial Omicron emergence at the University of Texas Medical Branch (UTMB) [Citation4]. The research protocol regarding the use of human sera was reviewed and approved by the UTMB Institutional Review Board (IRB number 20-0070). The de-identified human sera were heat-inactivated at 56°C for 30 min before the neutralization test. The genotype of infecting virus was verified by Sanger sequencing. Table S2 summarizes the serum information which was recently reported [Citation4].
Recombinant Omicron sublineage- or Deltacron XD-spike mNG SARS-CoV-2s
Recombinant Omicron sublineage BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA.4/5-, and Deltacron XD-spike mNG SARS-CoV-2s were constructed by engineering the complete spike gene from the indicated variants into an infectious cDNA clone of mNG USA-WA1/2020 [Citation12] (Figure S1(A)). All amino acid substitutions, deletions, and insertions in the variant-spike glycoproteins were introduced into the infectious cDNA clone of mNG USA-WA1/2020 using PCR-based mutagenesis as previously described [Citation13]. The BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA4/5-, and XD-spike sequences were based on GISAID EPI_ISL_6640916, EPI_ISL_6795834.2, EPI_ISL_12115772, EPI_ISL_7605591, EPI_ISL_11542270 (BA.4 spike is identical to BA.5 spike EPI_ISL_11542604), and EPI_ISL_10819657, respectively. Figure S1(A) depicts the spike mutations from different Omicron sublineages and Deltacron XD variant. The full-length cDNA of viral genome bearing the variant spike was assembled via in vitro ligation and used as a template for in vitro transcription. The full-length viral RNA was then electroporated into Vero E6-TMPRSS2 cells. On day 3–4 post electroporation, the original P0 virus was harvested from the electroporated cells and propagated for another round on Vero E6 cells to produce the P1 virus. The infectious titre of the P1 virus was quantified by fluorescent focus assay on Vero E6 cells (Figure S1(B)) and sequenced for the complete spike gene to ensure no undesired mutations. The P1 virus was used for the neutralization test. The protocols for the mutagenesis of mNG SARS-CoV-2 and virus production were reported previously [Citation14].
Fluorescent focus reduction neutralization test (FFRNT)
FFRNT was performed to measure the neutralization titres of sera against USA-WA1/2020, BA.1-, BA.2-, BA.2.12.1-, BA.3-, BA4/5-, and XD-spike mNG SARS-CoV-2s. The FFRNT protocol was reported previously [Citation15]. Vero E6 cells were seeded onto 96-well plates with 2.5 × 104 cells per well (Greiner Bio-one™) and incubated overnight. On the next day, each serum was two-fold serially diluted in culture medium and mixed with 100–150 FFUs of mNG SARS-CoV-2. The final serum dilution ranged from 1:20 to 1:20,480. After incubation at 37°C for 1 h, the serum-virus mixtures were loaded onto the pre-seeded Vero E6 cell monolayer in 96-well plates. After 1 h infection, the inoculum was removed and 100 μl of overlay medium containing 0.8% methylcellulose was added to each well. After incubating the plates at 37°C for 16 h, raw images of mNG foci were acquired using Cytation™ 7 (BioTek) armed with 2.5× FL Zeiss objective with a wide-field of view and processed using the software settings (GFP [469,525] threshold 4000, object selection size 50–1000 µm). The fluorescent mNG foci were counted in each well and normalized to the non-serum-treated controls to calculate the relative infectivities. The FFRNT50 value was defined as the minimal serum dilution to suppress >50% of fluorescent foci. The neutralization titre of each serum was determined in duplicate assays, and the geometric mean was taken. Tables S1 and S2 summarize the FFRNT50 results.
Supplemental Material
Download MS Word (3.1 MB)Acknowledgements
We thank the Pfizer-BioNTech clinical trial C4591001 and NCT04368728 participants, from whom the post-immunization human sera were obtained. We thank many colleagues at Pfizer and BioNTech who developed and produced the BNT162b2 vaccine. The collection and testing of BA.1-infected human sera at UTMB were supported by NIH grant HHSN272201600013C and an award from the Sealy & Smith Foundation. ML was supported by NIAID grant T35AI0778878.
Disclosure statement
X.X. and P.-Y.S. have filed a patent on the reverse genetic system of SARS-CoV-2. C.K., J.Z., H.X., X.X., and P.-Y.S. received compensation from Pfizer to perform the project. Q.Y., M.C., D.C., K.U.J., and K.A.S. are employees of Pfizer and may hold stock options. K.A.S. and Q.Y. are inventors on a patent application related to RNA-based COVID-19 vaccines. U.S. and A.M. are inventors on patents and patent applications related to RNA technology and COVID-19 vaccines. A.M is an employee of BioNTech and may hold stock options.
Additional information
Funding
References
- Kurhade C, Zou J, Xia H, et al. Neutralization of Omicron BA.1, BA.2, and BA.3 SARS-CoV-2 by 3 doses of BNT162b2 vaccine. Nat Commun. 2022;13:3602.
- Xia H, Zou J, Kurhade C, et al. Neutralization and durability of 2 or 3 doses of the BNT162b2 vaccine against Omicron SARS-CoV-2. Cell Host Microbe. 2022;30:485–488.e3.
- Colson P, Fournier PE, Delerce J, et al. Culture and identification of a “Deltamicron” SARS-CoV-2 in a three cases cluster in southern France. J Med Virol. 2022;94(8):3739–3749.
- Zou J, Kurhade C, Xia H, et al. Cross-neutralization of Omicron BA.1 against BA.2 and BA.3 SARS-CoV-2. Nat Commun. 2022;13:2956.
- Lyngse FP, Kirkeby C, Halasa T, et al. Nationwide study on SARS-CoV-2 transmission within households from lockdown to reopening, Denmark, 27 February 2020 to 1 August 2020. Euro Surveill. 2022;27(6):2001800.
- Peacock TP, Brown JC, Zhou J, et al. The SARS-CoV-2 variant, omicron, shows rapid replication in human primary nasal epithelial cultures and efficiently uses the endosomal route of entry. BioRxiv. 2022. doi:10.1101/2021.12.31.474653
- Redd AD, Nardin A, Kared H, et al. Minimal crossover between mutations associated with Omicron variant of SARS-CoV-2 and CD8(+) T-cell epitopes identified in COVID-19 convalescent individuals. mBio. 2022;13(2):e03617–21.
- Falsey AR, Frenck RW, Walsh EE, et al. SARS-CoV-2 neutralization with BNT162b2 vaccine dose 3. N Engl J Med. 2021;385(17):1627–1629.
- Mulligan MJ, Lyke KE, Kitchin N, et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature. 2020;586:589–593.
- Walsh EE, Frenck RW, Falsey AR, et al. Safety and immunogenicity of two RNA-based Covid-19 vaccine candidates. N Engl J Med. 2020;383:2439–2450.
- Liu Y, Liu J, Xia H, et al. Neutralizing activity of BNT162b2-elicited serum. N Engl J Med. 2021;384:1466–1468.
- Xie X, Muruato A, Lokugamage KG, et al. An infectious cDNA clone of SARS-CoV-2. Cell Host Microbe. 2020;27:841–848.e3.
- Xie X, Liu Y, Liu J, et al. Neutralization of SARS-CoV-2 spike 69/70 deletion, E484K and N501Y variants by BNT162b2 vaccine-elicited sera. Nat Med. 2021;27:620–621.
- Xie X, Lokugamage KG, Zhang X, et al. Engineering SARS-CoV-2 using a reverse genetic system. Nat Protoc. 2021;16:1761–1784.
- Zou J, Xia H, Xie X, et al. Neutralization against Omicron SARS-CoV-2 from previous non-Omicron infection. Nat Commun. 2022;13:852.