
Abstract
Cocoa (Theobroma cacao L. 1753) is a plant of great cultural, economic and ecological importance globally. By its structure and function maintains a wide variety of animal species. In Latin America, their crops are being affected by the disease called ‘frosty pod rot of cocoa’ caused by the fungus Moniliophthora roreri (Cif & Par) Evans. 2007, considered as the most destructive disease that attacks cocoa and kills it. The increase of temperature or precipitation because of warming global climate change could favor the fungus expansion to areas previously unaffected. We modeled the potential distribution for M. roreri and T. cocoa using MaxEnt and circulation model general HADCM3, A2a scenario with climatic variables for the present and the future (5, 35 and 65 years), in order to find sites suitable for monitoring to prevent the fungus spread, and to identify which variables determine the presence of M. roreri. The suitable areas for both species overlap from present to 2050, while they are different in 2080. M. roreri could extend from southern Ecuador to Venezuela, spread to westernmost Amazon, southwest Peruvian Amazon, and to the Peruvian-Bolivian border and the adjacent areas of Brazil. In both taxa the most influential variable seems to be precipitation of the wettest month. The Brazilian Amazon is the areas of South America with the largest plantations of T. cocoa, and therefore the most sensitive to the presence and proliferation of the fungus. For this reason we recommend to set up a monitoring system allowing early warning and control of incipient outbreaks that could eventually destroy Bolivian and Brazilian cocoa plantations.
El cacao (Theobroma cacao L. 1753) es una planta de gran importancia cultural, económica y ecológica a nivel mundial. Por su estructura y función mantiene una gran diversidad de especies animales. En América Latina sus cultivos están siendo afectados por la enfermedad llamada ‘podredumbre del cacao’ provocada por el hongo, Moniliophthora roreri (Cif. & Par.) (Evans, 2007), considerada como la enfermedad más destructiva que ataca al cacao, causándole la muerte. El aumento de temperatura o cambio de precipitación debido al calentamiento climático global podrían favorecer que este hongo se expanda a áreas no antes afectadas. Se modeló la distribución potencial para M. roreri y T. cacao usando MaxEnt y el modelo de circulación general HADCM3, escenario A2a con variables climáticas para el presente y el futuro (5, 35 y 65 años), con el fin de encontrar sitios de monitoreo idóneos para frenar la expansión del hongo e identificar qué variables determinan la distribución de M. roreri. La distribución de áreas idóneas coinciden para ambas especies desde el presente hasta el año 2050, pero en 2080 las áreas predichas como idóneas para cada una son diferentes. M. roreri se podría extender en el extremo occidental de la Amazonía, desde el sur de Ecuador hasta Venezuela, en el suroeste de la Amazonía peruana, la región fronteriza con Bolivia y en las áreas adyacentes de Brasil. En T. cacao y M. roreri la variable que más influyó es la precipitación del mes más húmedo. La Amazonía brasileña es la zona de Sudamérica con extensiones de T. cacao más importantes, y por tanto sensible a la presencia y proliferación del hongo. Por este motivo se sugiere necesario crear un cordón de monitoreo constante que permita alertas tempranas y control de los brotes incipientes que podría terminar con todos los cultivos de cacao brasileños y también bolivianos.
Palabras claves:
Introducción
El cacao, Theobroma cacao L. (1753), es un árbol perennifolio nativo del trópico americano, especialmente del sur de México y América Central [Citation1]. El cacao es una planta de uso ancestral, cuya difusión de su uso se le atribuye a los mayas, y que debido a un gran movimiento transoceánico ahora se cultiva en varios continentes [Citation1], constituyendo una parte importante dentro de las actividades culturales, ecológicas y económicas a nivel mundial. T. cacao es uno de los motores económicos mundiales para los países productores (Bolivia, Brasil, Colombia, Ecuador, entre otros). Sin embargo, su importancia cultural sobrepasa fronteras políticas alcanzando una trascendencia histórico-ecológica mundial, debido a que las plantaciones de T. cacao forman agro-ecosistemas que por su estructura y función mantienen una diversidad de aves, murciélagos, mamíferos no voladores e invertebrados –especialmente hormigas– similar a la de los bosques naturales y superior a la de los hábitats agrícolas más intervenidos [Citation2–4]. Adicionalmente, las plantaciones de T. cacao también contribuyen a la fijación de CO2 atmosférico, a la conservación del suelo y el agua, a la polinización y a otros servicios ecosistémicos [Citation5].
El cultivo de T. cacao en América Latina está siendo afectado por la enfermedad ‘podredumbre del cacao’, provocada por el hongo Moniliophthora roreri (Cif. & Par.) (Evans, 2007). Considerada como la enfermedad más destructiva que ataca a las poblaciones de cacao del género Theobroma y Herrania, siendo posible que afecte a todas las especies de estos dos géneros [Citation6,7]. El impacto económico y ecológico de esta enfermedad ha llegado a originar pérdidas de más del 90% de la producción, ocasionando la pérdida de hábitat para flora y fauna, fragmentación, erosión del suelo [Citation8] y abandono de áreas de cultivo enteras [Citation9–11]. M. roreri se detectó originalmente en la parte nororiental de Colombia, y posteriormente se expandió hacia Ecuador [Citation12,13]. A partir de 1970 alcanzó Perú (1980) [Citation13,17], Bolivia, Venezuela, Panamá (1956), Costa Rica (1978) [Citation13,15], Honduras (1997), Guatemala (2002), Belice (2004) México (2005) [Citation16], Bolivia [Citation13] y actualmente está expandiéndose hacia Brasil [Citation17].
Su capacidad adaptativa [Citation16] y el cambio climático podrían favorecer su distribución geográfica hacia el Caribe insular y las Guyanas en las Américas [Citation18], así como los continentes productores de cacao a granel: África y Asia [Citation19]. El hongo puede utilizar como vectores de dispersión el agua, los insectos, el viento, así como una gran variedad de actividades antropogénicas [Citation11,20–22]. No obstante, la dispersión y severidad de la enfermedad dependen mucho de las condiciones ambientales, especialmente de la humedad y temperatura, así como del manejo de la plantación [Citation20,23,24], esta es una de las principales causas de la dificultad de controlar la enfermedad.
Resulta urgente entender la relación T. cacao-M. roreri, considerando aspectos ecológicos de las especies, con énfasis en la distribución y posible expansión de la M. roreri sobre T. cacao en sitios aún no afectados, con el fin de establecer medidas para prevenir la infección. El desarrollo de Modelos de Distribución de Especies (MDE), el avance de los Sistemas de Información Geográfica (SIG) y la creciente accesibilidad a información espacial relacionada con variables ambientales ha aumentado considerablemente nuestra capacidad de predecir las áreas de distribución de las especies. En este estudio se usan MDE para evaluar la distribución potencial de M. roreri en cultivos de T. cacao y los impactos del cambio climático sobre esta enfermedad en las próximas décadas; sin embargo no se toma en cuenta factores humanos que podrían ser importantes (e.g., movimientos de plantas de un lugar geográfico a otro). Los resultados apoyan la hipótesis de que el hongo se expenderá hacia áreas que al momento no están siendo infectadas.
Métodos
Especies focales
T. cacao, es un árbol perennifolio, que mide de 4 a 7 m de altura, y que cultivado puede crecer hasta 20 m o más cuando crece sobre sombra intensa y según las condiciones ambientales alcanza diversos tamaños [Citation1]. Crece en áreas con topografía plana u ondulada; necesita temperaturas medias de entre 22° a 26° C, y las extremas dependen de la altitud sin embargo no baja de los 15 °C ni sube de los 32 °C; siendo óptima de 25 °C porque permite la floración y fructificación [Citation25]. El cacao no soporta temperaturas bajo 0° aunque sea por poco tiempo puede provocar la aparición de enfermedades criptogámicas [Citation26]. La precipitación pluvial mínima y máxima entre 1400 a 3000 mm y óptima de 1500 a 2500 mm bien distribuidos a lo largo del ciclo. El déficit a 100 mm afecta a la floración y brote de hojas. La humedad relativa anual promedio corresponde al 70 o 80%. Necesita sombra que le proteja de la radiación y de la evaporación excesivas siendo de del 40 al 50% para el cultivo en crecimiento (menor de 4 años) y del 60 al 75% para plantaciones en producción [Citation1]. Actualmente se cultiva en regiones calientes muy húmedas; constituye una importante fuente generadora de ocupación, de divisas y en especial ha impulsado la economía de los países productores [Citation27]. Se cultiva principalmente en 13 países; en América el mayor productor es Brasil, con el 18% de la producción mundial, le sigue Ecuador con el 6% [Citation4], y finalmente, Colombia y México con el 1%. El continente africano, con Costa de Marfil, Ghana, Nigeria y Camerún genera cerca del 60% de la producción mundial [Citation4].
M. roreri, es un hongo parásito del grupo de los basidiomicetes, perteneciente a la familia Marasmiaceae [Citation28]. M. roreri se conoce con distintos nombres, como enfermedad acuosa, pringue, hielo, pasmo, enfermedad palúdica, ceniza, polvillo [Citation9,29], o enfermedad de Quevedo, en referencia al lugar de aparición de la misma [Citation14,21].
Ciclo de la enfermedad, empieza en los residuos contaminados de cosecha. Las conidias son diseminadas por el viento y la lluvia, ocurriendo también contaminación de frutos o mazorcas con moniliasis de una plantación a otra. Las esporas también son movilizadas durante las labores de cosecha en el aire y bajo condiciones propicias de humedad y temperatura antes señaladas, así estas estructuras infectan constantemente los frutos que recién están formándose [Citation30,31]. La penetración del hongo, ocurre directamente a través de los estomas, creciendo entre las células del córtex, produciendo conidias dentro y en la superficie de los frutos [Citation32]. El período de incubación del hongo es largo, antes de aparecer los síntomas [Citation33]. El tiempo de infección puede ser de 3 a 8 semanas, pudiendo variar según la edad del fruto, la severidad del ataque, la susceptibilidad del árbol y las condiciones de clima, principalmente presencia de lluvias, mientras que en frutos tiernos, en días lluviosos y calurosos, el período de incubación se acorta a tres semanas [Citation34], sin embargo, Cruz en el año 1993 relata que el período de incubación (latente) fluctúa entre 30 y 70 días [Citation35].
Datos de distribución
El área de distribución geográfica en este estudio incluye los siguientes países: Ecuador, Colombia, Perú, Venezuela, Bolivia, Brasil, Panamá, Costa Rica, Nicaragua, Honduras, Guatemala, Belice y México. En todos ellos hay cultivos de cacao, y en todos ellos se ha confirmado la presencia de M. roreri, a excepción de Brasil. Se compiló una base de datos con 136 localidades geográficas para cacao y 67 para M. roreri.
Los datos de distribución de T. cacao fueron tomados de TROPICOS (Missouri Botanical Garden), Royal Botanic Gardens (Kew), New York Botanical Garden (NYBG), Herbario Nacional del Ecuador (QCNE), Comisión Nacional para el Conocimiento y el Uso de la Biodiversidad (CONABIO), Ministerio de Agricultura y Desarrollo Rural – Colombia, Fondo para el Financiamiento del Sector Agropecuario (FINAGRO), Centro Agronómico Tropical de Investigaciones y Enseñanza – Costa Rica (CATIE), Plan Ejecutivo para el Cultivo del Cacao – Brasil (CEPLAC), Instituto Nicaragüense de Tecnología Agropecuaria (INTA), seminarios de la Corporación Colombiana de Investigación Agropecuaria (Corpoica), Organización Internacional del Cacao (ICCO), Federación Nacional del Cacao – Colombia (Fedecacao) y World Cocoa Foundation (WCF).
Los datos de M. roreri fueron tomados del Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), Departamento de Agricultura de los Estados Unidos (USDA), Food and Agriculture Organization of the United Nations (FAO), Sanidad Agraria de Perú (SENASA), Instituto de Cultivos Tropicales – Perú (ICT), Servicio de Información Agroalimentaria y Pesquera (SIAP), Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP) en Ecuador, Fundación Hondureña Investigaciones Agrícolas (FHIA). Los datos que no contaban con coordenadas geográficas fueron georreferenciados utilizando información de gaceteros y mapas a escala 1:50.000.
Variables Climáticas
Se utilizaron 19 variables climáticas actuales y futuras (5, 35 y 65 años) de WorldClim 1.4 correspondientes al modelo de circulación general HADCM3 bajo el escenario A2. Este escenario presupone un mundo heterogéneo con poco desarrollo tecnológico, distribución muy desigual de los recursos y crecimiento demográfico alto [Citation36]. Los datos se descargaron de http://www.worldclim.org/download. Estas 19 variables bioclimáticas se utilizan ampliamente en estudios de modelado por considerarse biológicamente significativas para caracterizar distribuciones de especies, ya que representan tendencias anuales, estacionalidad y factores limitantes o extremos [Citation36]. Todas las variables fueron cortadas usando la extensión GridMachine en Arcview 3.2 para incluir solamente el área de estudio, con una resolución espacial de 0.008333°, equivalente a ~1 km en el Ecuador.
Modelos de distribución de especies
Actualmente existen varios métodos que pueden ser utilizados para generar las distribuciones de especies [Citation37–39]. En este caso se utilizó MaxEnt 3.3.1, ya que es muy eficaz en comparación con otros métodos [Citation36,40–42]. Se utilizó el 70% de los datos de distribución de ambas especies para generar el modelo de distribución y el 30% para evaluarlos. MaxEnt hace predicciones usando únicamente datos de presencia. Este programa modela una distribución probable potencial del hábitat disponible en el área de estudio. Esta distribución modelada considera los datos ambientales y de distribución geográfica conocida para la especie. De acuerdo con MaxEnt, la mejor distribución potencial de una especie es aquella más cercana a tener iguales probabilidades de presencia a lo largo de toda el área de estudio, sujeta a la restricción de que la expectativa para cada variable ambiental, incluida en la distribución modelada, debe concordar con su promedio empírico a lo largo de la distribución real conocida. Debido es esto la probabilidad calculada en MaxEnt no es de presencia, sino de similitud con el hábitat observado. MaxEnt asigna un valor de idoneidad de hábitat en el área de estudio que varía desde 1 –el más idóneo– hasta 0, que indica nula idoneidad para el organismo modelado.
Para cada especie y periodo temporal (presente, 2020, 2050 y 2080) se realizaron 100 modelos, de las que se derivó el modelo promedio [Citation43]. La evaluación sólo se realizó para los modelos del presente, ya que no es posible realizarla sin tener datos de presencias reales, y para ello se utilizó el estadístico ‘área bajo la curva ROC’, o AUC. Sus valores varían entre 0,5 y 1, en donde 0,5 equivale a una clasificación al azar mientras que un valor de 1 indica un ajuste perfecto [Citation44–46]. Los mapas y AUC finales son los promedios.
Análisis de datos
Los modelos resultantes de MaxEnt fueron convertidos a mapas binarios utilizando el umbral específico de corte ‘Maximum training sensitivity plus specificity’, que es el umbral que optimiza sensibilidad, o casos correctamente clasificados como presencias, y la especificidad, o casos correctamente clasificados como ausencias [Citation46–49].
Un requisito importante para manejar, conservar y controlar la dispersión de las especies patógenas es conocer la influencia del ambiente y sus variables climáticas [Citation50–52]. Para identificar cuáles son las variables que determinan la distribución de M. roreri y T. cacao se analizó uno de los test de Maxent: ‘jackknife regularized training gain’ o ganancia regularizada, tanto para M. roreri como para T. cacao. Se seleccionó el de ganancia regularizada ya que trabaja con 70% de los datos del modelo permitiendo comparar y correlacionar variables entre uno y otro modelo en caso de ser necesario. Para saber si las variables climáticas que influían en M. roreri eran las mismas que para T. cacao, se compararon los dos análisis de jackknife.
Resultados
Análisis de los modelos
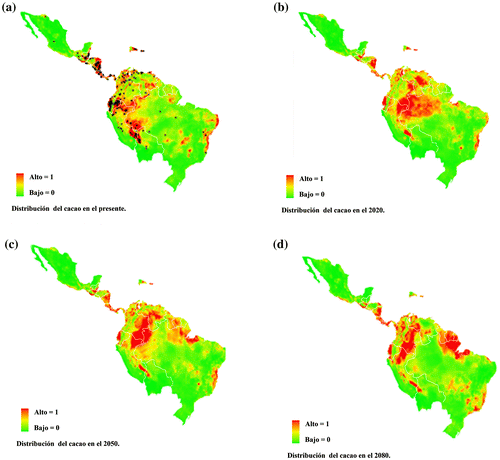
La distribución potencial de T. cacao y M. roreri fue predicha por MaxEnt con valores altos de AUC para el presente: 0.906 para M. roreri y 0.882 para T. cacao. Estos altos valores justificaron que se realizaran las proyecciones para el futuro. En el modelo del presente (Figura (a)), T. cacao se encuentra en regiones tropicales y subtropicales de México y Centro América, en los bosques latifoliados húmedos y subtropicales de Ecuador, Colombia, Perú, las Yungas de Bolivia, Venezuela, Guyanas, Surinam y al sureste en la Amazonía brasileña, en donde también se encuentra bosques tropicales, subtropicales y sabanas con pastizales y matorrales. Las condiciones de hábitat óptimo para esta especie se reducen en 2020 (Figura (b)) en los bosques latifoliados húmedos y subtropicales de Costa Rica, Valle de Magdalena en Colombia, río Napo entre Ecuador y Perú, y las ramas occidental y central de los Andes en Ecuador, pero también se observan condiciones óptimas en los bosques latifoliados húmedos y subtropicales de Iquitos en Perú, y la Amazonía brasileña y venezolana. Para 2050 (Figura (c)) existen zonas favorables para las poblaciones de T. cacao, aunque en Brasil se detecta una disminución de la idoneidad en la Amazonía. Esto determina una gran disminución del hábitat idóneo para plantar T. cacao en este país cacaotero. Aun así, el área alrededor de la frontera con Guyana y Surinam es propicia para esta especie, además de su costa Atlántica. En 2080 (Figura (d)), las condiciones para T. cacao son favorables en los bosques latifoliados húmedos y subtropicales de la costa del Pacífico en México, Costa Rica, Panamá, Surinam, mientras que disminuye en los cerrados de Brasil, la Amazonía colombiana, Guatemala y la República Dominicana. En general, se observa que a lo largo del tiempo cambian las condiciones climáticas y también el hábitat idóneo para T. cacao, con una disminución considerable en el área de cultivo idóneo en el continente americano.
Figura 1. Distribución potencial de T. cacao para los escenarios climáticos del presente (a), 2020 (b), 2050 (c) y 2080 (d). Los puntos representan la distribución conocida de la especie. Los colores indican de verde (0) a rojo (1) la idoneidad de hábitat de acuerdo con MaxEnt.
En la actualidad, M. roreri se conoce de 12 países. De acuerdo con el mapa de distribución potencial con datos de Presente (Figura (a)), la Amazonía brasileña se encuentra en riesgo debido a una posible expansión de M. roreri, lo mismo que puede ocurrir a lo largo del arco formado por el extremo occidental de la Amazonía, desde el sur de Ecuador hasta Venezuela. El riesgo de colonización es inminente en el suroeste de la Amazonía peruana, la región fronteriza que avanza aún más en Bolivia que está siendo afectada actualmente por M. roreri y que para este estudio no se tomó en cuenta debido a que el registro oficial se publicó en este mismo año, pero fue predicho por el modelo; y por supuesto, las áreas adyacentes de Brasil se sitúan riesgo potencial. En Centro América, M. roreri muestra zonas de alta idoneidad en la costa de México, Nicaragua y las zonas fronterizas entre Honduras y Costa Rica. En 2020 (Figura (b)) su dispersión podría ampliarse a otros países de donde aún no hay registros, y se podría expandir con facilidad a través de la frontera entre Colombia, Venezuela para finalmente llegar a Brasil, lo que podría causar la devastación de plantaciones en grandes áreas.
Figura 2. Distribución potencial de M. roreri, para los escenarios climáticos del presente (a) 2020 (b), 2050 (c) y 2080 (d). Los puntos representan la distribución conocida de la especie. Los colores indican de verde (0) a rojo (1) la idoneidad de hábitat de acuerdo con MaxEnt.
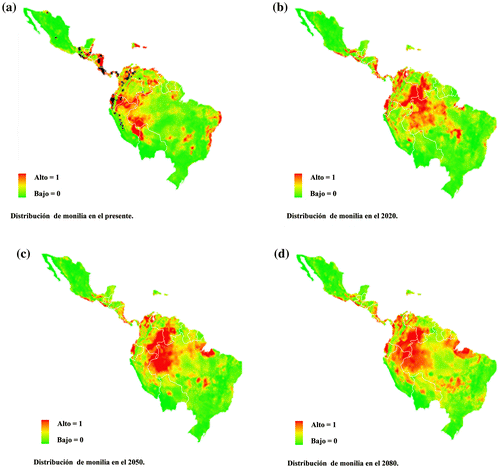
Para 2050 (Figura (c)), las plantaciones de T. cacao en otras regiones de Brasil, especialmente en los Departamentos de Acre, Rondonia y Amapa, podrían infectarse. En Perú se verían afectadas las estribaciones orientales de los Andes, Tumbes y Piura, así como la Amazonía fronteriza con Colombia y Bolivia, lo que facilitaría la entrada de la enfermedad a Brasil. Las zonas más afectadas en Venezuela serían Bolívar, Amazonas y Delta Amacuro en su parte fronteriza con Guyana. En Colombia, los departamentos de Vichada, Guainía, Vaepés y Amazonas, fronterizos con Brasil. Su avance en dirección Sur y Este continúa amenazando también a la casi totalidad de los estados brasileños de Roraima y Amazonas. Finalmente, hacia el norte se extendería por las zonas costeras de los países centroamericanos hasta algunos estados de México.
En 2080 (Figura (d)), el hábitat idóneo para M. roreri se extendería hacia otros Departamentos de Perú (Amazonas, Huanuco, Libertad, Arequipa), en Brasil hacia Maranhao y Matogrosso, mientras que en Venezuela se mantendría en los mismos lugares que en el 2050. Es importante destacar que para 2080 existe una disyunción entre las distribuciones predichas para T. cacao con respecto a la de M. roreri.
Análisis de las variables climáticas
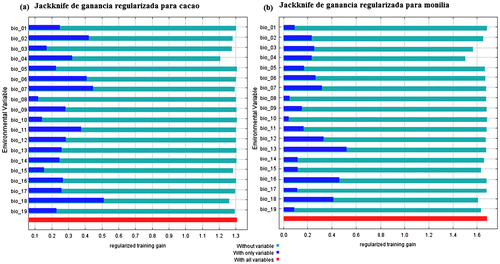
Las variables que de acuerdo al estadístico jackknife tienen una mayor influencia en la distribución del cacao son la precipitación del trimestre más cálido (bio 18), el rango anual de temperatura (bio 7), el rango de temperatura diurno promedio que refiere a la temperatura máxima y mínima (bio2), temperatura mínima del mes más frío (bio 6), temperatura media del trimestre más frío (bio 11) y la estacionalidad de temperatura (bio 4) (Figura (a)). Para M. roreri, las variables con mayor influencia son la precipitación del mes más húmedo (bio 13), la precipitación del trimestre más húmedo (bio16), la precipitación del trimestre más cálido (bio 18), la precipitación anual (bio 12), el rango de temperatura anual (bio 7) y la temperatura mínima del mes más frío (bio 6) son las más influyentes. Al igual que en T. cacao, para M. roreri, la variable más influyente y que interaccionan en las dos especies permitiendo que el patógeno sea específico de cacao son la bio 18, bio 7 y bio 6.
Figura 3. Jackknife de ganancia regularizada para T. cacao (a) y M. roreri (b). La variable que más aporta a ambos modelos es la precipitación del mes más lluvioso.
Análisis de los valores de idoneidad a lo largo del tiempo
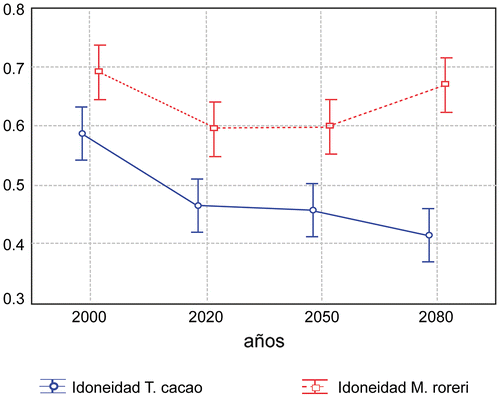
La distribución potencial actual de M. roreri se superpone con la de T. cacao. Para 2020 el área predicha para M. roreri aumenta, y aún más en 2050, aunque se reduce ligeramente en la Amazonía brasileña hacia 2080 (Figura (a)–(d)). En el caso de T. cacao, no hay una disminución sensible del área total, pero sí se observa un desplazamiento de las áreas idóneas a zonas en las que actualmente esta planta no crece. En todos los periodos analizados hay una alta superposición de las áreas de alta idoneidad de M. roreri y T. cacao (Figuras (a–d) and (a–d)).
La idoneidad de ambas especies sigue patrones semejantes hasta 2050, para cambiar totalmente en 2080, cuando se observa un aumento del área geográfica idónea para M. roreri y una disminución para T. cacao (Figura ). Es especialmente preocupante que hay zonas de superposición de alta idoneidad para ambas especies en el norte (especialmente en Amapá, Maranhao) y medio oeste (Matogroso, Pará, Rondonia) de Brasil. Sin embargo se observa claramente que existen zonas (Amazonas, Acre) que no serán favorables para el cultivo de T. cacao aunque en la actualidad son altamente productivas.
Figura 4. Análisis de idoneidad que representan la disposición espacial de los hábitats mínimos potencialmente adecuados para el desarrollo de M. roreri y T. cacao, que muestra las condiciones climáticas afines y distintas para M. roreri y para T. cacao. En 2080 las condiciones son diferentes.
Sitios recomendados para monitoreo ante la posible expansión de M. roreri
La distribución predicha por MaxEnt identifica áreas importantes para la prevención, alerta y control de la enfermedad, especialmente en Brasil, ya que en el resto de países la enfermedad ya es un hecho.
Discusión
La distribución potencial de Theobroma cacao para América Latina se ve comprometida en el futuro por la expansión predicha –con altos valores de idoneidad– de M. roreri. Los modelos predicen para T. cacao una reducción del área considerada idónea, así como una disminución en los valores de idoneidad con respecto al presente. Aunque las áreas predichas para T. cacao y M. roreri no son totalmente coincidentes en 2080, en los periodos previos hay una alta correspondencia, por lo que es posible que en este proceso las poblaciones de T. cacao sufran tales tasas de pérdida que no puedan recuperarse por sí solas, incluso en ausencia de M. roreri. Tres factores importantes hacen imparable la expansión del hongo a las plantaciones de T. cacao: (1) la alta adaptación del hongo a los cultivos de T. cacao, (2) la resistencia del hongo al ambiente cambiante y (3) la principal causa de la dispersión de M. roreri es la actividad humana, imprescindible en cualquier cultivo.
M. roreri se ha especializado durante 200 años en atacar cultivos de Theobroma, ya que ha sido capaz de adaptarse a este ambiente antropizado [Citation53,54], favorecido por las propias condiciones de los cultivos. La resistencia adquirida por M. roreri se detectó inicialmente en los bosques de la Cordillera Occidental de los Andes, al noroeste de Colombia [Citation55]. Los primeros datos, fechados en 1817, proceden de plantaciones de esa área, aunque la adaptación histórica de M. roreri a los nuevos ambientes causó su diferenciación en grupos genéticos con características morfo-fisiológicas propias pero similarmente agresivas frente a T. cacao [Citation12,17]. Phillips-Mora en el 2003[Citation12] descubrió que M. roreri puede crecer y desarrollarse entre 18 y 30 °C, y la espora puede germinar de 10 a 40 °C. Además, puede soportar temperaturas de 55 °C [Citation56]. Estos rangos de temperatura exceden las condiciones normales de crecimiento de T. cacao. La distribución observada de M. roreri, mucho más limitada que lo que estas condiciones indican, pudiera deberse a lo limitado de la distribución de T. cacao, y que cualquier ampliación en la distribución del cultivo iría acompañada por M. roreri, perfectamente capaz de vivir en condiciones mucho más amplias que las que ahora conocemos [Citation57].
La mayor tasa de germinación de esporas ocurre cuando son sometidas durante 8 ó 9 horas a humedades relativas de 80,5–100% y temperaturas de 20–24 °C, mientras que a menor humedad disminuye dicha tasa, hasta hacerse nula en esporas secas [Citation7,29]. El agua es el principal componente para la germinación de las esporas, y a veces el único sustrato necesario para iniciarla [Citation58]. La alta temperatura y humedad que caracterizan las áreas tropicales en las que se cultiva T. cacao, favorecen el inicio y desarrollo de la enfermedad [Citation59], permitiendo que este patógeno pueda reproducirse y diseminarse durante todo el año [Citation53,54].
Los resultados de MaxEnt indican que la variable de mayor importancia para modelar la distribución de T. cacao es la precipitación del trimestr más cálido, seguida del rango anual de temperatura. La explicación de este fenómeno es que, si bien en la cabecera del Amazonas –su centro de diversidad–, las temperaturas varían entre 20–30 °C, en los árboles del piso inferior de la selva tropical la variación diurna se reduce a alrededor de 6 °C [Citation60]. Los parámetros de las dos especies se ajustan perfectamente a la temperatura media diaria en los trópicos, por lo que es posiblemente una consecuencia de la adaptación a las condiciones ambientales donde T. cacao crece normalmente [Citation60].
Las variables de mayor importancia para determinar el hábitat idóneo de M. roreri son la precipitación del mes más húmedo y la precipitación del trimestre más húmedo, que confirma lo encontrado en otros estudios, que concluyeron que en lugares con lluvias frecuentes e intensas la incidencia de M. roreri es mayor (80% infección) que en sitios secos [Citation6,61]. Otras variables contribuyen con la dispersión diurna de M. roreri, el rango anual de temperatura (bio 7) y las temperaturas mínimas del mes más frío (bio 6) [Citation62]; requiere temperaturas frías en los meses más fríos y calientes en los meses más cálidos, superiores a 26 °C las cuales favorecen la liberación y dispersión de los conidios; las lluvias intensas y frecuentes favorecen la presencia de agua libre sobre los frutos, facilitando la germinación y penetración de los conidios [Citation13]. Finalmente, una humedad relativa menor del 80% favorece la dispersión [Citation63], ya que con sólo 4 mm diarios de precipitación que mojen el follaje y saturen el ambiente de humedad producen el ambiente adecuado para el desarrollo de la infección [Citation24].
Las condiciones microclimáticas necesarias para la producción de T. cacao son las óptimas para la infección y para la diseminación del patógeno [Citation24]. Por otro lado, la estacionalidad de la temperatura también favorece que el aumento de M. roreri no sea explosivo sino progresivo, por lo que en plantaciones jóvenes y vigorosas la aparición de la enfermedad es lenta y no causa muchas pérdidas [Citation24], pero éstas aumentan con la edad de la plantación hasta convertirlas en totalmente deficitarias. Es fundamental resaltar que las variables climáticas juegan un papel importante en el desarrollo y en general en todas las funciones vitales de M. roreri y T. cacao, pero en la actualidad aún no se sabe cómo se produce la diseminación del hongo, aunque es poco probable que el hongo se haya propagado de forma natural [Citation13].
Áreas prioritarias para la prevención, alerta y control de M. roreri
Hasta el momento ha sido imposible erradicar por completo M. roreri una vez que se establece en una región. Para frenar la diseminación del hongo es necesario y urgente trabajar entre los municipios de los países fronterizos afectados. Hay acciones que han dado resultados en Venezuela (Mérida, Zulia, Táchira, Apure y Barinas), de donde el hongo es endémico, como la prohibición de trasladar material de T. cacao (excepto almendras beneficiadas), hacia los estados del centro y oriente del país, los cuales se encuentran libres de la enfermedad. En México, para contener la dispersión desde Tabasco se estableció en el 2005 un cinturón de seguridad de 5 km de ancho a lo largo de la frontera con el Estado de Chiapas, aún libre de M. roreri. Esta área ha sido vigilada constantemente por el CESVETAB (Comité de Sanidad Vegetal de Tabasco) para la detección precoz de nuevas plantaciones afectadas y aplicar un conjunto de prácticas de cultivo para el control de la enfermedad y evitar una mayor propagación.
Este es el primer intento que se ha realizado para modelar la distribución potencial y posible efecto de un hongo hacia la planta de cacao. Sin embargo, otros factores adicionales a los climáticos podrían influenciar la distribución de las especies estudiadas. Por ejemplo, otros factores tales como las capacidades de dispersión de las especies focales, sus capacidades evolutivas para adaptarse a condiciones nuevas, y factores antropogénicos, como el cambio y uso de suelo o la translocación de organismos [Citation64]. Este trabajo se enfocó en factores climáticos, ya que estos limitan la distribución de los organismos afectando su crecimiento y su supervivencia (p.g., temperaturas letales altas y mínimas).
Conclusiones
Los modelos demuestran que existe una estrecha relación entre las condiciones climáticas y la distribución de estas especies, lo que confirma los resultados de estudios previos. Las principales condiciones para determinar el crecimiento de M. roreri están relacionadas con la precipitación, la temperatura mínima del mes más frío, la máxima del más cálido y la humedad relativa. Su combinación permite que se den las condiciones microambientales óptimas para el hongo, el propio T. cacao [Citation24] y su distribución en Latinoamérica, lo que hace necesario considerar las variables de cambio climático, mismas que darán a futuro, las condiciones agroecológicas para nuevos sitios de cultivo del cacao y en consecuencia la probable proliferación de la enfermedad. Así también la diseminación del hongo a cultivos no contaminados.
Teniendo en cuenta que M. roreri es un patógeno obligado y por tanto su distribución está grandemente restringida por la distribución del cacao, los modelos predicen una expansión de la M. roreri a regiones ahora libres de la misma, es imperativo establecer medidas para la detección precoz de plantaciones afectadas y aplicar un conjunto de prácticas de cultivo intercalar con otras especies forestales permitiendo la conexión (corredores biológicos) entre áreas fragmentadas, mitigando así efectos de cambio climático, proveyendo de un hábitat y recursos para la conservación de flora y fauna.
La Amazonía brasileña es la zona de Sudamérica con cultivos de T. cacao más extensos e importantes [Citation16] y por tanto sensible a la presencia y aún más a la proliferación del hongo. Es necesario crear un cordón de control y monitoreo constante que permita alertas tempranas y detección de brotes incipientes. En caso contrario, M. roreri podría devastar los cultivos de cacao brasileños y bolivianos, como está comenzando a suceder en la actualidad [Citation7,16,65].
Las acciones o alternativas de prevención deben estar dirigidas a las zonas de frontera y rutas de ríos con Perú, Colombia y Venezuela, donde es posible el transporte de plantas, frutos u otros vectores de la enfermedad. Para ello es importante adoptar medidas de cuarentena y planes de contingencia, sobre todo en los estados de Bahía, Espíritu Santo y Pará, que tienen contacto fluvial, y también en Rondonia y Amazonas, que están cerca de las fronteras de riesgo de M. roreri. El trabajo conjunto con los municipios fronterizos para difundir y concienciar a los agricultores, personal técnico en particular y a la población en general; en el reconocimiento de los síntomas.
Es necesario instaurar prácticas culturales eficaces y el uso adecuado de variedades de café evitando el reemplazo de variedades de origen trinitarios, que si bien es cierto dan mayor producción y una aparente resistencia a esta enfermedad, han provocado la pérdida de diversidad genética [Citation16,66].
Contribución del autor
SMO diseñó el estudio y escribió el primer borrador del manuscrito. TP, PT, JM, participaron en el diseño de los métodos, análisis de los resultados, escritura del articulo y revisión del mismo.
Agradecimientos
Al Misouri Botanical Garden (St. Louis, USA) y a las instituciones auspiciantes, el Consejo Superior de Investigaciones Científicas y la Universidad Internacional Menéndez Pelayo, de España, y la Universidad Central del Ecuador. Vladimir Sandoval revisó versiones previas del manuscrito.
Associate Editor: Veerle Vanacker
Declaración de divulgación
No hay conflicto de intereses y puede ser reproducido.
Financiación
Este estudio fue financiado por el Missouri Botanical Garden (St. Louis, USA).
Related Research Data
Referencias
- Ogata N. Comisión Nacional Para El Conocimiento y uso de la Biodiversidad. El Cacao. Biodiversitas. 2007 May–Jun;72:1–5. Spanish. Disponible en internet: http://www.biodiversidad.gob.mx/Biodiversitas/Articulos/biodiv72art1.pdf
- Bueno J, Álvarez F, Santiago S. Biodiversidad del Estado de Tabasco [Internet]. Version 6.0. México (MEX): Instituto de Biología UNAM- CONABIO [cited 2015 Agosto 12]. Spanish. Disponible en internet: https://books.google.com.ec/books?id=105sMZ6e1dIC&printsec=frontcover&source=gbs_ge_summary_r&cad=0#v=onepage&q&f=false
- Salgado M. Biodiversidad en Agroecosistemas. In: Ramírez G. La moniliasis un desafío para lograr la sostenibilidad del sistema cacao en México. Tecnología en Marcha [Internet] [cited 2015 Agosto 12]; 2008;21(1):97–110. Spanish. Disponible en internet: http://dialnet.unirioja.es/descarga/articulo/4835688.pdf
- Ramírez G. La moniliasis un desafío para lograr la sostenibilidad del sistema cacao en México. Tecnología en Marcha [Internet] [cited 2015 Agosto 12]. 2008;21(1):97–110. Spanish. Disponible en internet: http://dialnet.unirioja.es/descarga/articulo/4835688.pdf
- Somarriba E. Agroforestería con cacao Paper presented at: Seminario Internacional de la Corporación Colombiana de Investigación Agropecuaria (Corpoica); 2007; CATIE, Talamanca (CR). Spanish.
- Evans HC. Pot rot of cacao caused by Moniliophthora roreri. CMI. Phytopathological Papers. 1981;24:44.
- Phillips-Mora W, Baqueros F, Melnick RL, et al. First report of frosty pod rot caused by Moniliophthora roreri on cacao in Bolivia. BSPP. 2015;31:29.
- Sánchez F, Gamboa E, Rincón J. Control químico y cultural de la moniliasis (Moniliophthora roreri Cif y Par) del cacao (Theobroma cacao L) en el estado Barinas. Rev Fac Agron. 2003;20:188–194. Spanish.
- Rorer JB. Enfermedades y Plagas del Cacao en el Ecuador y Métodos Modernos Apropiados al Cultivo del Cacao. Paper presented at: Asociación de Agricultores; 1918; Guayaquil (EC). Spanish.
- Parsons JJ. The antioquenian colonization of western Colombia. Vol. 32. UC Publications in Ibero-Americana; 1949. p. 233. Medellín (CO). English.
- Enríquez GA, Brenes O, Delgado J. Desarrollo e impacto de la moniliasis del cacao en Costa Rica. Paper presented at: Conferencia Internacional de Investigación en Cacao; 1981; Fedecacao; Cartegena (CO). Spanish.
- Phillips-Mora W. Origin, biogeography, genetic diversity and taxonomic affinities of the cacao fungus Moniliophthora roreri as determined using molecular, phytopathological and morpho-physiological evidence [dissertation]. University (UK): Universities in the United Kingdom; 2003.
- Phillips-Mora W. Origen, biogeografía, diversidad genética y afinidades taxonómicas del hongo Moniliophthora roreri Cif y Par y del cacao (Theobroma cacao L.) determinadas mediante evidencia molecular, fitopatológica y morfofisiológica [Internet]; 2005 [cited 2015 Agosto 13]. Available from: www.Catie.ac.cr
- Hernández TA, Aranzazu F, Arévalo E, et al. La moniliasis del cacao en el Perú. Agrotrópica. 1990;2:56–58. Spanish.
- Enríquez G, Suárez C. Moniliophthora roreri disease of cacao in Costa Rica. Rev Interam Psicol. 1978;28:339–340.
- Phillips-Mora W, Wilkinson MJ. Frosty pod of cacao: a disease with a limited geographic range but unlimited potential for damage. Phytopathology. 2007;97(12):1644–1647.10.1094/PHYTO-97-12-1644
- Parra D, Perez S, Sosa D, et al. Avances en las investigaciones venezolanas sobre enfermedades del cacao. RET. 2009;1(2):55–74. Spanish.
- Phillips-Mora W. Avances en CATIE en el conocimiento del hongo causante de la moniliasis: aplicaciones prácticas. Paper presented at: Hacia una cacaocultura competitiva en la Región Andina para su inserción en los mercados internacionales: Experiencias exitosas; 2007; Bucaramanga (CO). Spanish.
- Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO). Perspectivas a plazo medio de los productos básicos agrícolas –proyecciones al año 2010 [Internet]. 2004:95. Spanish. Disponible en internet: ftp://ftp.fao.org/docrep/fao/007/y5143s/y5143s00.pdf
- Krauss U, Hidalgo E, Bateman R, et al. Improving the formulation and timing of application of endophytic biocontrol and chemical agents against frosty pod rot (Moniliophthpora roreri) in cocoa (Theobroma cacao). Biol Control. 2010;54(3):230–240.10.1016/j.biocontrol.2010.05.011
- Barros NO. Investigaciones sobre el hongo Moniliophthora roreri Cif y Par, causante de la pudrición acuosa de la mazorca del cacao: sus daños y su control. El Cacaotero Colombiano. 1977;3:42–52. Spanish.
- Evans HC, Stalpers JA, Samson RA, et al. On the taxonomy of M. roreri roreri, an important Pathogen of Theobroma cacao in South America. Can J Bot. 1978;56:2528–2532.10.1139/b78-305
- Porras VH, González LC. Epifitiología de la moniliasis del cacao (Moniliophthora roreri) y su relación con el ciclo de producción en la zona de Matina, Costa Rica. Fitopatología. 1984;19(2):78–84. Spanish.
- Ampuero C. Moniliophthora roreri pod rot of cocoa. Cocoa Grower’s Bull. 1967;9:15–17.
- CORPEI, ANECACAO, FEDECADE, UNOCACE. (base de datos en línea). Solicitud de Protección de la Denominación de Origen “Cacao Arriba”. MIC, ONUDI, Cacao, Estudio Agroindustrial en el Ecuador: Competitividad de la cadena de valor y perspectivas de mercado, (EC); 2007.
- Enriquez G. Curso sobre el cultivo de cacao. Turrialba (CR): Centro Agronómico Tropical de Investigación y Enseñanza. 1985:339:5–23. Spanish. Disponible en internet: https://books.google.es/books?hl=es&lr=&id=eZgOAQAAIAAJ&oi=fnd&pg=PA5&dq=Enriquez+G.+Curso+sobre+el+cultivo+de+cacao,+Turrialba,+Costa+Rica:+Centro+Agrono%CC%81mico+Tropical+de+Investigacio%CC%81n+y+Ensen%CC%83anza.+1985&ots=IpuMZ0Pg8O&sig=gPQNyPfAaOOGC0BmrMfXzWeHsbg#v=onepage&q&f=false
- Meléndez-Marín L. Microambiente, cantidad de esporas en el aire e incidencia del hongo Moniliophthora roreri (Cif & Par.) Evans et al., bajo tres sistemas de manejo de sombra leguminosa en cacao (Theobroma cacao) [dissertation]. Centro Agronómico Tropical de Investigaciones y Enseñanza (CATIE): Universidad de Costa Rica; 1993. Spanish. Disponible en internet: http://www.worldcocoafoundation.org/scientific-research/researchlibrary/pdf/MelendezMarin1993.pdf
- Phillips-Mora W. Evaluación de la resistencia de cultivares de cacao (Theobroma cacao L.) a Moniliophthora roreri Cif y Par. Evans et al. [dissertation]. Centro Agronómico Tropical de Investigaciones y Enseñanza (CATIE): Universidad de Costa Rica. 1986:100. Spanish. Disponible en internet: http://orton.catie.ac.cr/REPDOC/A1720E/A1720E.PDF
- Aime MC, Phillips-Mora W. The causal agents of witches broom and frosty pod rot of cacao (chocolate, Theobroma cacao. L) form a new lineage of Marasmiaceae. Mycologia. 2005;97:1012–1022.
- Amores F, Agama J, Suárez C, et al. EET 575 y EET 576 nuevos clones de cacao nacional para la zona central de Manabí. Estación Experimental Tropical “Pichilingue” 2009; 346:28; Quevedo (EC). Spanish. Disponible en internet: http://repositorio.iniap.gob.ec/handle/41000/1517
- Campuzano H. Influencia de la temperatura y la humedad en la germinación de esporas de M. roreri. Paper presented at: 8th International Cocoa Research Conference; 1981. p. 493–497; Cartagena (CO). Spanish.
- Albuquerque PSB, Bastos CN, Luz EDMN, et al. Doenças do cacaueiro (Theobroma cacao). Em: Kimati H, Amorim L, Rezende JAM, et al, editors. Manual de Fitopatologia, 2005:2:151–163; Piracicaba (BR). Portugués.
- Johnson J, Bonilla J, Aguero L. Manual de manejo y producción del cacaotero; 2008. Leon (NI). Spanish. Disponible en internet: http://cenida.una.edu.ni/relectronicos/RENF01J71.pdf
- AGRÍCOLAS. Fundación Hondureña Investigaciones. Identificación y control de la moniliasis del cacao. Diseño e Impresión: Centro de Comunicación Agrícola de la FHIA; 2003. La Lima, Cortés (HO). Spanish.
- Cruz BS. Determinación de fuentes de resistencia de cacao de origen nacional al ataque de Monilia roreri (Cif. & Par.) [Tesis Ing. Agr.]. Universidad de Guayaquil (EC); 1993.
- Hijmans RJ, Cameron SE, Parra JL, et al. Very high resolution interpolated climate surfaces for global land areas. Int J Climatol [Internet]. 2005;25:1965–1978. Available from: http://onlinelibrary.wiley.com/doi/10.1002/joc.1276/pdf10.1002/(ISSN)1097-0088
- Levy PE, Cannell MGR, Friend AD. Modelling the impact of future changes in climate, CO2 concentration and land use on natural ecosystems and the terrestrial carbon sink. Glob Environ Change. 2004;14(1):21–30.10.1016/j.gloenvcha.2003.10.005
- Anderson RP, Dudik M, Ferrier S, et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography. 2006;29(2):129–151.
- Guisan A, Zimmermann NE. Predictive habitat distribution models in ecology. Ecol Model. 2000;135(2):147–186.10.1016/S0304-3800(00)00354-9
- Peterson AT, Soberón J, Sánchez-Corder V. Conservatism of ecological niches in evolutionary time. Science. 1999;285(5431):1265–1267.10.1126/science.285.5431.1265
- Peterson AT, Ortega-Huerta MA, Bartley J, et al. Future projections for Mexican faunas under global climate change scenarios. Nature. 2002;416:626–629.10.1038/416626a
- Phillips SJ. Transferability, sample selection bias and background data in presence-only modelling: a response to Peterson et al. (2007). Ecography. 2008;31(2):272–278.10.1111/j.0906-7590.2008.5378.x
- Phillips SJ, Dudík M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography. 2008;31(2):161–175.10.1111/j.0906-7590.2008.5203.x
- Phillips-Mora W, Ortiz CF, Aime M. Cincuenta años de la podredumbre moniliasis en Centroamérica: Cronología de su propagación y el impacto de Panamá a México. Paper presented at: 15a conferencia Internacional de Investigación del Cacao; 2006. p. 1–12; San José (CR). Spanish.
- Fielding AH, Bell JF. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ Conserv. 1997;24(1):49.
- Loiselle B, Howell C, Graham C, et al. Avoiding pitfalls of using species distribution models in conservation planning. Conserv Biol. 2003;17:1591–1600.10.1111/cbi.2003.17.issue-6
- Merow C, Smith MJ, Silander JA. A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter. Ecography [Internet]. 2013;36:1058–1069. Available from: http://docs.google.com/viewer?a=v&pid=sites&srcid=ZGVmYXVsdGRvbWFpbnxuaWNoZXNhbmRhcmVhc29mZGlzdHJpYnV0aW9ufGd4OjU3ZmVkYzNmZWVlOWI2ZmY10.1111/ecog.2013.36.issue-10
- Zweig MH, Campbell G. Receiver-operating characteristics (ROC) plots: a fundamental evaluation tool in clinical medicine. Clin Chem. 1993;39:561–577.
- Liu C, Berry PM, Dawson TP, et al. Selecting thresholds of occurrence in the prediction of species distributions. Ecography. 2005;28:385–393.10.1111/eco.2005.28.issue-3
- Jimenez-Valverde A, Lobo JM. The ghost of unbalanced species distribution data in geographical model predictions. Divers Distribut. 2006;12(5):521–524.10.1111/ddi.2006.12.issue-5
- Fry W. Epidemiology: influence of the biotic environment. In: Principles of plant disease management. New York, NY: Academic Press; 1982. p. 68–82. Available from: https://books.google.com.ec/books?hl=es&lr=&id=n1kxOTitCAgC&oi=fnd&pg=PP1&dq=Principles+of+plant+disease+management.+New+York&ots=9PZjGQUJYl&sig=iDB23oL2IKSAnnafSeCw9JHGgGY#v=onepage&q=Principles%20of%20plant%20disease%20management.%20New%20York&f=false
- Agrios GN. Plant pathology. Vol. 5. Gainesville (FL): Elsevier. 2004. p. 952. ISBN: 9780120445653.
- Thuiller W, Richardson D, Pysek P, et al. Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale. Glob Change Biol. 2005;11(12):2234–2250.10.1111/gcb.2005.11.issue-12
- Carlile MJ, Watkinson SC, Gooday GW. The Fungi. London (UK): Academic Press; 1994. Scienc Direct. ISBN: 978-0-12-738445-0
- Wang XR, Szmidt AE. Population genetic analysis of pathogenic forest fungi. Molecular biology of woody plants. 2000. p. 491–520. The Netherlands: Springer.10.1007/978-94-017-2311-4
- Phillips-Mora W, Krauss U, Evans H, et al. Genetic diversity of the cacao pathogen Moniliophthora roreri (Cif.) Evans et al. in Tropical America. Vol. 2. Paper presented at: 14th International Cocoa Research Conference; 2003. p. 613–618; Acra (GH).
- Ram A. Biology, epidemiology and control of moniliasis (Moniliophthora roreri) [dissertation]. Silwood Park, Berkshire, England: University of London; 1989.
- Evans HC. Invasive neotropical pathogens of tree crops. In: Watling R, Frankland JC, Ainsworth AM, et al., editors. Tropical mycology: Micromycetes. Vol. 2; 2002. p. 83–112. New York (NY): CABI Publishing.
- Glottieb D. The physiology of spore germination in fungi. Bot Rev. 1950;16(5):229–257.
- Claytoni CN. The germination of fungus spores in relation to controlled humidity. Phitopathology. 1942;32:921–943.
- Wood GAR, Lass RA. Cocoa. 4th ed. Cornwall (UK): Blakwell Science; 1985. p. 620.
- Aranzazu HF. La moniliasis del cacao. Vol. 3. Paper presented at: Tercera Reunión del Comité Consultivo de Sanidad Vegetal; 1982. p. 22–26; Caracas (VE). Spanish.
- Dowding P. Water availability, the distribution of fungi and their adaptation to the environment. Paper presented at: Symposium of the British Mycological Society; 1986; University of Cambridge, Cambridge (EU).
- Soberón J, Peterson AT. Interpretation of models of fundamental ecological niches and species’distributional areas. Biodiv Inform. 2005;2:1–10.
- Moraes WB, Júnior J, Peixoto LDA, et al. An analysis of the risk of cocoa moniliasis occurrence in Brazil as the result of climate change. Summa Phytopathol. 2012;38(1):30–35.
- Porras VH. Epidemiología de la moniliasis (M. roreri Cif y Par.) del cacao y su relación con la producción del árbol en la zona de Matina [dissertation]. San José (CR): Universidad de Costa Rica; 1986. p. 51.