
Abstract
Human activity can lead to changes in animal behavior and distribution patterns if the level of activity is high enough to cause disturbance. Both humans and other animals commonly use human-made trails in lowland tropical forest and, therefore, it is possible that use of trails by humans might affect the likelihood that animals would use those same trails. We investigated this possibility at a site in lowland forest of eastern Ecuador using camera traps to document numbers of people and numbers of other animals at a series of camera locations during January–March, 2014–2016. Human activity was higher on trails used by all visitors to the research station than on two research plots where human activity is restricted. Ungulates were the only group to show a strong negative relationship with human activity but only on trails open to all visitors; there was no apparent negative impact of human activity on research plots. Results suggest that the level of human activity along trails is not now causing negative impacts to most mammals but also that levels of activity should be monitored to ensure that any increase in human presence does not lead to negative responses by mammals.
Las actividades humanas pueden producir cambios en el comportamiento y los patrones de distribución de los animales silvestres, siempre y cuando dichas actividades sean lo suficientemente altas como para crear un disturbio. En los bosques bajos tropicales, tanto humanos como otros animales usan los senderos hechos por personas; y, es posible que el uso de los senderos por humanos pueda afectar a la probabilidad de que los animales ocupen esos mismos senderos. Se ha investigado esta posibilidad en un bosque bajo tropical el Este de Ecuador mediante el uso de cámaras trampa para documentar el número de personas y animales registrados en una serie de estaciones de muestreo durante enero a marzo de 2014 a 2016. La actividad humana fue mayor en los senderos usados por todos los visitantes de la estación y menor en dos parcelas permanentes de uso restringido. Sólo los ungulados fueron los únicos en mostrar una relación negativa fuerte con la actividad humana, pero sólo en los senderos abiertos a todo el público. No existió un impacto negativo aparente de la actividad humana en las parcelas permanentes. Los resultados sugieren que el nivel de actividad humana en los senderos no está causando un impacto negativo a la mayoría de mamíferos, pero también que los niveles de actividad deberían ser monitoreados para asegurar que cualquier incremento de la presencia humana no tenga un efecto negativo en las respuestas de los mamíferos.
Introduction
Human activity can have negative impacts on many species, leading to changes in distribution (moving away from human activity), abundance, and activity patterns [Citation1–4]. Further, responses of animals to human activity may vary among species [Citation2,5,6] and thereby affect the assemblage of species found in a given location. Predators, for example, may avoid areas with high levels of human activity, whereas the reverse may be true for their prey (i.e. human activity may create predator-free spatial refuges for prey) [Citation7]. Biological reserves, such as field stations, often have multiple uses, including research, education, and ecotourism. Depending on the levels of human activity associated with these different uses, it is possible that populations of species that the reserves are designed to protect may be adversely affected. Thus, development of adequate management plans may need to take into account levels of human activity.
Measuring levels of activity or patterns of distribution can be difficult for many species. Cameras triggered by heat and motion (camera traps) are an important tool for wildlife studies and are particularly useful in tropical forests where animals often are difficult to observe or quantify [Citation8,9]. Camera placement and spacing can affect the likelihood that animals will encounter cameras, and thereby be recorded [Citation9]. Many studies, particularly those that seek to estimate densities of felids (e.g. jaguars, Panthera onca, [Citation10–12]; ocelots, Leopardus pardalis, [Citation13–15]), place cameras along travel routes, which may include game trails, human-use trails, creek beds, and dirt roads [Citation8,10–12,14,16–18], some of the same trails that may be used by people. Use of travel routes for camera placement has been questioned because of the possibility that results would not be representative of the community at large because not all species are equally likely to use trails (e.g. [Citation19–21]). Effects, if present, of trails may depend on the type of ‘trail’ and its width, which may affect which species use it or how frequently [Citation20,22]. Similarly, it is possible that the level of human traffic along trails might affect the numbers and identities of animals captured by camera traps when set along the same trails that humans use [Citation1,4].
In a previous study [Citation22], we did not find a correlation between numbers of people and other animals photographed by cameras on trails in lowland tropical forest but results were based on a single sample period (60 days). Here, we use data gathered across three years from camera traps set along narrow trails (created by humans) in lowland forest of eastern Ecuador to examine the hypothesis that presence of people on trails has a negative influence (leads to reduced presence) on use of those same trails by other animals. The basic question, then, is whether the number of animals recorded in camera traps is negatively correlated with number of people recorded at the same camera location. Our study area is a research station (~650 ha) located within a large expanse of forest with comparatively little human impact in the form of logging, fragmentation, or hunting. On the other hand, the station is visited by many educational groups each year and a smaller number of researchers. Thus, human presence along trails can be substantial but variable. The mission of the station is to support research, education and conservation, so it is important to know how or if the level of human activity affects animals; such information can be important for management (e.g. minimizing use of and impacts to trails) both for the station itself and, more broadly, for conservation policies implemented within Yasuní National Park.
Study site and methods
Study site
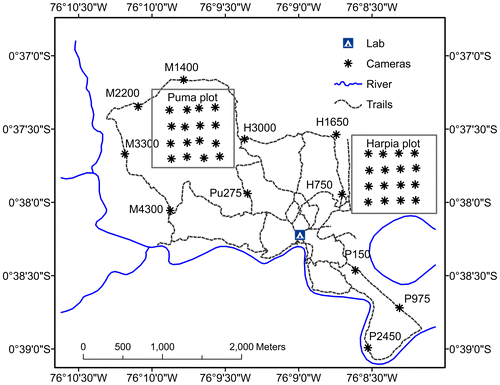
Research was conducted at Tiputini Biodiversity Station (TBS), Orellana Province, Ecuador (ca 0°37′ S, 76°10′ W, 190–270 meters above sea level). TBS encompasses ~650 ha and is located adjacent to and partially within Yasuní National Park on a tract of largely undisturbed (by human activities) lowland rain forest within the biologically diverse Yasuní Biosphere Reserve [Citation23]. The biosphere reserve encompasses Yasuní National Park (~1,000,000 ha) [Citation24], the adjacent Waorani Ethnic Reserve (~700,000 ha), and a 10-km buffer zone. The station and nearby areas are dominated by terra firme forest; várzea forest, palm swamps, and various successional habitats also are present. Mean annual precipitation at Yasuní Research Station, approximately 30 km WSW of TBS, is about 3100 mm. Trails at the station (~30 km; Figure ) have been in existence for ~20 years and are regularly maintained (treefalls cleared, vegetation cut back somewhat) but are narrow, typically <0.5 m wide, sometimes up to 1 m wide. Consequently, canopy cover is not affected by trail presence and trails are not particularly noticeable when approached perpendicularly to the direction of the trail. Trails are used by students, researchers, and station personnel. In addition, there are two 100-ha study plots [Citation25] that were established in 2001 (Figure ). Study plots have transects every 100 m in east–west directions and every 200 m in north-south. Transects are approximately the same width as regular trails and are typically cleared during November–December of each year. Study plots are used by researchers but not by student groups.
Figure 1. Camera locations on station trails and on 100-ha study plots at TBS, Ecuador.
Camera traps
Cameras triggered by an infrared heat-and-motion detector were located approximately 1–1.2 km apart along station trails used by student groups as well as researchers within terra firme forest (Figure ) from mid-January until mid-March, 2014–2016 (~60 days/year). Although the station is generally busy throughout the year, January–May are the busiest months and August–September are the slowest. If human activity has a negative impact on animal activity, we might expect it to occur during the period when the station has the most visitors.
Cameras were placed at sites that showed evidence of animal activity (e.g. tracks) or at sites where topography made occurrence likely; we did not use any chemical or other attractants at camera locations. Two cameras (one each of Reconyx PC800 Hyperfire Professional IRTM and PC850 Hyperfire Pro White FlashTM; Reconyx, Inc., Holmen, Wisconsin, USA), were placed at each location, on opposite sides of the trail, approximately 0.5 m off the ground. There were 11 separate locations at which cameras were placed (Figure ) but only 10 pairs of cameras were in operation each year. Cameras at Pu275 (Figure ) were operational in 2014 and 2015 but not 2016; cameras at M1400 were operational only in 2016. On study plots, where student activity is restricted, cameras were located ~200 m apart in a grid of 16 cameras on each plot; only one camera was placed at each location on the study plots. All cameras were set to take 5 photographs at each detection, with a 1-s delay between images. We set cameras with a minimum time between photographs of 5 min. Cameras remained continuously activated (except when malfunctions occurred); date and time were automatically stamped on each photograph. Cameras were visited at approximately monthly intervals to check batteries and camera placement (cameras were occasionally disturbed by white-lipped peccaries Tayassu pecari).
Cameras along trails were typically identified by small signs alerting groups to their presence. Students, researchers, and guides were told to ignore the cameras (i.e. not try to avoid having photos taken) but to simply continue walking along trails or transects.
Analyses
We summarized images by species and date. We classified photographs as belonging to independent records if more than 30 min elapsed between consecutive photographs of the same species at a given location [Citation26]. We indexed activity in terms of number of photographs/100 trap days (hereafter referred to as capture rates; i.e. captures of images) irrespective of the number of individuals in each image (e.g. peccaries and people often were represented by multiple individuals). We do not assume that such rates are a true estimate of actual abundance [Citation9] but simply assume that, for comparative purposes, such rates provide a reasonable estimate of activity at a camera location [Citation2,7,27]. We calculated number of trap days for each camera location as the time the camera was placed in operation until it was taken down or until the last photograph was taken (based on the date and time stamp on the photographs) if batteries had failed or other malfunctions had occurred. We combined records across months during each year. We used repeated measures ANOVA to compare human activity (images/100 trap days) among plot and trail cameras and across years. We used correlation analyses to examine relationships between number of images of people and for separate groups of mammals (ungulates, carnivores, rodents, xenarthrans). We also examined the relationship between numbers of images of people and other animals with distance (straight-line distance determined from GPS data) from station buildings to cameras along trails. We also calculated distance based on the shortest distance used by walking along trails; these two distance measures were highly correlated (r = 0.989), so we used straight-line distance only in analyses. We used linear regression to examine the relationship between human and ungulate activity, with all years combined. Capture rates (images/100 days) were transformed (Ln + 1) prior to analyses to meet assumptions of tests. All statistical tests were performed using Statistix 10 [Citation28].
Results
Human activity
Independent records of humans along station trails varied both among years and among camera locations (Table ). Human activity was higher and more variable, particularly in 2015 and 2016, among cameras located on trails when compared to cameras located on plots (Figure ; repeated measures ANOVA, camera location [trails, Harpia plot, Puma plot], F = 10.89, df = 2,119, p < 0.001; year effect, F = 6.91, df = 2,119, p < 0.002). Mean activity across three years (mean of annual mean captures across cameras) on trails (63.4 ± 7.5 SD/100 trap days) was more than double that on either plot (Harpia, 23.4 ± 8.0; Puma, 27.8 ± 5.9). Similarly, mean coefficient of variation of activity was higher on trails (90.0 ± 4.2) than on Puma (65.4 ± 4.7) or Harpia (50.4 ± 7.7). Human activity along trails was negatively related to distance from station buildings (Table ).
Table 1. Numbers of trap days and numbers of independent records of people, other mammals, and birds from camera traps located on station trails at Tiputini Biodiversity Station, Ecuador, 2014–2016. Cameras were operated from mid-January to mid-March; no camera was operated at M1400 in 2014 or 2015 and not in 2016 for Pu275. See Figure 1 for camera locations.
Figure 2. Human activity (photographic images/100 trap days; means ± se across cameras within a year) along main station trails and on two 100-ha study plots at TBS, Ecuador.
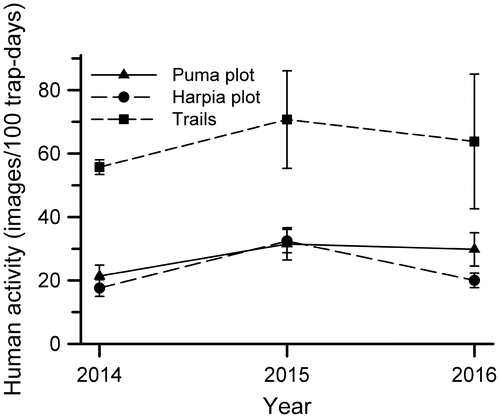
Table 2. Correlation coefficients between capture rates (images/100 trap days) of different animal groups and human activity along trails (Trails) and on two 100 ha study plots (Harpia, Puma; see Figure 1) at TBS, Ecuador, 2014–2016. Also shown are correlation coefficients between capture rates of animals and people and distance from the station to cameras on trails (Trails – distance).
Effects on animals
Independent records of mammals also varied among years and among camera locations along trails (Table ) but with a few exceptions, there was little evidence that animal activity (as measured by images/100 trap days) was negatively related to human activity (Tables and ). Ungulate activity showed the strongest negative correlation with human activity along trails (Figures and ), although the strength of the relationship was much weaker in 2014 than in subsequent years (Tables and ), perhaps because there were fewer people recorded on trails during 2014 (Figure ). In contrast, there was no evidence of negative effects on ungulates in relation to human activity on either plot (Table ). Ungulate activity on trails was positively associated with distance from the main station buildings, likely reflecting the negative relationship between distance and human activity (Table ).
Table 3. Correlation coefficients between capture rates (images/100 trap days) of four ungulate species and human activity along trails at TBS, Ecuador, 2014–2016.
Figure 3. Ungulate and human activity (ln + 1 transformed photographic images/100 trap days) at cameras along trails at TBS, Ecuador. Years are indicated separately but the regression equation is based on all years combined (n = 30).
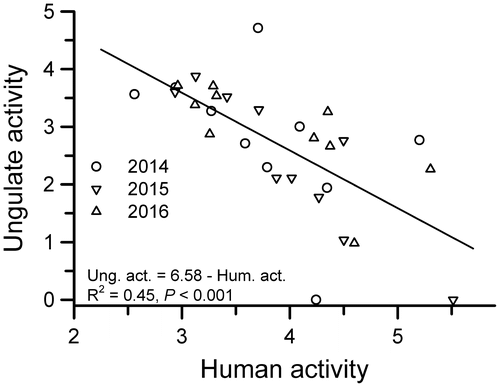
Figure 4. Activity of four species of ungulates in relations to human activity (ln + 1 transformed photographic images/100 trap days) at cameras along trails at TBS, Ecuador. Regression equations are based on all years combined (n = 30): P. tajacu activity = 3.93–0.59Human activity, R2 = 0.14, p < 0.05; T. pecari activity = 4.43–0.84Human activity, R2 = 0.32, p < 0.001; M. americana activity = 3.64–0.55Human activity, R2 = 0.13, p < 0.05; T. terrestris activity = 2.93–0.59Human activity, R2 = 0.25, p < 0.005.
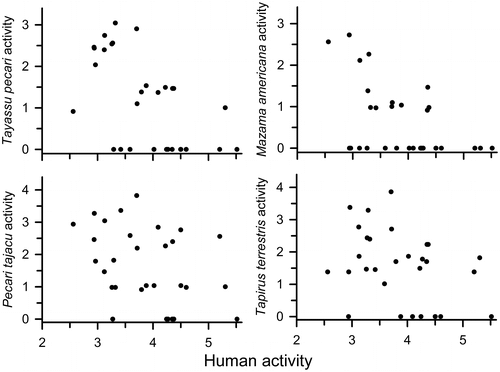
Given the overall negative relationship with human activity on trails seen for all ungulates combined, we examined the relationship separately for four species of ungulates (Mazama americana, Pecari tajacu, T. pecari, and Tapirus terrestris); there were too few records of M. nemorivaga to analyze separately. As was true for all ungulates combined, correlations between animal activity and human activity were strongest in 2015 for three of the four species; M. americana showed the strongest negative correlation in 2016. When data from all three years were combined, all species showed significant negative relationships with levels of human activity (Figure ). The relationship was strongest for T. pecari and T. terrestris and weaker but still negative for the other two species.
Rodent activity was negatively correlated with human activity on the Harpia plot but only in 2016, whereas xenarthrans were positively correlated with human activity on Harpia but only in 2014. There was no evidence of negative or positive effects of human activity on any groups on Puma.
Discussion
Human activity along station trails appears to have had little impact on use of those trails by other animals. Ungulates were the only group that appeared to show a reasonably strong negative reaction to human activity but only in two of three years. Although the difference among years was not significant, there tended to be fewer people recorded on trails in 2014, when there was less evidence of a negative impact on ungulate activity. Ungulate species also differed in their apparent responses to humans, with T. pecari showing the strongest overall negative relationship with human activity. T. pecari occurs in much larger groups, often >100 individuals than any of the other species, including the much smaller P. tajacu. A larger group size may make it easier to detect humans and might help explain the negative relationship with human activity. In general, human activity declined at farther distances from the station and may help account for the positive relationship between peccaries and distance from the station. In contrast to trails open to all station visitors, there was little to no indication of negative effects of human activity along trails on two 100-ha research plots where access is limited and overall human activity was one-third that of trails.
The overall lack of apparent impact on animal use of trails might indicate that human activity, although common, was not sufficiently intense to elicit negative responses from most species. In a broad-scale survey of human and wildlife activity along hiking trails in eastern North America, Kays et al. [Citation6] found no strong or consistent avoidance of hiking trails by animals. They suggested that species might become habituated to periodic occurrence of humans with short-term responses (moving away from trails temporarily) rather than long-term avoidance of areas with human presence. Such short-term responses would be difficult to detect with camera trap records. In that study, overall human activity on trails (~72 people/100 trap days) was similar to the overall rate in the current study (~64/100 trap days). In contrast, Muhly et al. [Citation7] found that human activity, depending on the level, could have positive impacts on prey species activity and negative impacts on predators. The positive impacts were strongest at locations with >32 people/day, whereas the impact on predators could be felt at sites with >18 people/day, levels of activity much greater than in the present study.
In this study, activity was indexed in terms of number of photographs separated by at least 30 min for the same species, including humans, irrespective of the number of individuals captured in an image. People, as is true for peccaries, often travel in groups (particularly for students) and larger groups might have a greater impact on animal activity (i.e. easier to detect and avoid). Larger groups also may be louder and, again, might have a large impact on animal behavior. Images from camera traps do not, however, provide a good assessment of the numbers of individuals in a particular group as the number of people in a given image depends on rates of travel and how spread out the group is. Thus, we were not able to assess the impact of group size on animal activity. In contrast to student groups, researchers on study plots typically are in small groups or single individuals and, as a consequence, may have less of an impact on animal behavior. Furthermore, we only examined effects on animals in terms of their activity as measured by number of photographs. It also is possible that presence of humans could have other impacts that might not be reflected in capture rates. For example, with increased human activity, individuals may need to spend more time being vigilant, leaving less time for other activities such as foraging, which could lead to increased levels of stress, with possible long-term consequences.
No cameras in the present study were located less than ~750 m from the station center of activity. Thus, it is possible that negative impacts might be greater – but not detected in this study – closer to station buildings where human activity is certainly higher (more noise, including generator; more people working close to the station lab building, etc.). Perhaps there is some threshold of activity above which negative effects are seen but below which there is little impact.[Citation7] Further studies that include cameras located closer to the station buildings would be necessary to determine whether impacts of human activity are greater closer to the station. Given that activity on trails tended to be greater farther away from the station, it is entirely possible that an even stronger relationship might be present closer to the station.
Similarly, although this study encompassed three years, it is possible that negative impacts of human activity, assuming they exist, may take longer to accumulate and become detectable. Impacts of human activity also may depend on the type of activity. Simple periodic occurrence of people along trails may not be sufficient to cause much negative reaction (e.g. moving away from trails, fewer photographs). Animals might also simply move away from trails temporarily when humans are present but not move out of the area altogether or may change activity patterns to become more nocturnal [Citation1]. Where human activity is more intense (along roads, hunting) animals may display stronger reactions [Citation6,29,30]. Occurrence of T. pecari, for example, was greater farther from a road in Yasuní National Park [Citation31]; roads provide increased access to hunters and peccaries are a favored target of Waorani. Overall mammal activity at mineral licks also was lower at sites closer to Waorani settlements than at sites located within TBS, where hunting is largely absent [Citation32]. Similarly, brocket deer (M. americana) altered their temporal activity patterns in areas subject to hunting when compared to areas protected from human activity [Citation33]. In the study by Kays et al. [Citation6], cameras were used to measure animal activity along hiking trails in areas open to hunting and in areas without hunting in 32 protected forest areas in the eastern US. Although they found no strong evidence to suggest avoidance of hiking trails in the absence of hunting, hunting pressure did apparently cause some avoidance of areas by the four most frequent targets of hunters.
Human activity may, if too intense, have a variety of negative consequences for wildlife. Consequently, in biological reserves that have to balance human activity with protection of wildlife, the level (and types) of human activity may need to be managed or regulated to prevent negative impacts to wildlife. In the present study, there was little evidence to suggest that current levels of human activity had a significant negative impact on most wildlife. Yet, the apparent negative response of some ungulates does suggest that the level of activity may be reaching a point where more negative responses are likely to occur. Furthermore, because we did not sample areas close to the center of human activity (i.e. close to station buildings), it is possible that negative influences may be present in a more concentrated area. Yet, primate troops (e.g. Ateles belzebuth, Lagothrix lagotricha, Saimiri sciureus) are common in the vicinity of station buildings, often passing over or by the lab, cabins, and other buildings, suggesting that some animals are either not bothered by or become habituated to human activity. Similarly, although anecdotal evidence (e.g. observations of natural history guides present at the station for many years) suggests that some large animals (e.g. T. terrestris) are not as common close to the station as they once were, others still are regularly encountered (e.g. Cuniculus paca, Dasypus novemcinctus, Priodontes maximus). Continued monitoring of the levels of human activity and the responses of animals will be needed to ensure that interactions between humans and wildlife are managed properly. If negative effects of human activity increase over time, it is possible that the station may need to consider limiting human activity along some trails or temporarily closing some trails to human use.
Funding
This work was supported by Universidad San Francisco de Quito and University of Florida.
Disclosure statement
No potential conflict of interest was reported by the authors.
Associate Editor: Esteban Suarez
Acknowledgments
We appreciate the help of Consuelo Barriga in facilitating our work at Tiputini and the staff that make working there such a pleasure. Work at Tiputini Biodiversity Station was conducted in accordance with research permit No. 0005-FAU-MAE-DPOPNY, Ministerio de Ambiente, Ecuador.
Related Research Data
References
- Griffiths M, van Schaik CP. The impact of human traffic on the abundance and activity periods of sumatran rain forest wildlife. Conserv Biol. 1993;7(3):623–626.10.1046/j.1523-1739.1993.07030623.x
- Foster RJ, Harmsen BT, Doncaster CP. Habitat use by sympatric jaguars and pumas across a gradient of human disturbance in Belize. Biotropica. 2010;42:724–731.10.1111/j.1744-7429.2010.00641.x
- Ciuti S, Northrup JM, Muhly TB, et al. Effects of humans on behaviour of wildlife exceed those of natural predators in a landscape of fear. PLoS One. 2012;7:e50611.10.1371/journal.pone.0050611
- Zhou Y, Buesching CD, Newman C, et al. Balancing the benefits of ecotourism and development: the effects of visitor trail-use on mammals in a protected area in rapidly developing China. Biol Conserv. 2013;165:18–24.10.1016/j.biocon.2013.05.007
- George SL, Crooks KR. Recreation and large mammal activity in an urban nature reserve. Biol Conserv. 2006;133:107–117.10.1016/j.biocon.2006.05.024
- Kays R, Parsons AW, Baker MC, et al. Does hunting or hiking affect wildlife communities in protected areas? J Appl Ecol. 2016;54:242–252. DOI:10.1111/1365-2664.12700.
- Muhly TB, Semeniuk C, Massolo A, et al. Human activity helps prey win the predator-prey space race. PLoS One. 2011;6:e17050.10.1371/journal.pone.0017050
- Blake JG, Mosquera D, Loiselle BA, et al. Temporal activity patterns of terrestrial mammals in lowland rainforest of eastern Ecuador. Ecotropica. 2012;20:137–146.
- Burton AC, Neilson E, Moreira D, et al. REVIEW: Wildlife camera trapping: a review and recommendations for linking surveys to ecological processes. J Appl Ecol. 2015;52:675–685.10.1111/1365-2664.12432
- Kelly MJ. Jaguar monitoring in the Chiquibul Forest. Belize Carib Geogr. 2003;13:19–32.
- Wallace RB, Gómez H, Ayala G, et al. Camera trapping for jaguar (Panthera onca) in the Tuichi Valley. Bolivia Mastozool Neotrop. 2003;10:133–139.
- Maffei L, Cullar E, Noss A. One thousand jaguars (Panthera onca) in Bolivias Chaco? Camera trapping in the Kaa-Iya National Park. J Zool. 2004;262:295–304.10.1017/S0952836903004655
- Trolle M, Kéry M. Estimation of ocelot density in the pantanal using capture–recapture analysis of camera-trapping data. J Mammal. 2003;84:607–614.10.1644/1545-1542(2003)084<0607:EOODIT>2.0.CO;2
- Di Bitetti MS, Paviolo A, De Angelo C. Density, habitat use and activity patterns of ocelots (Leopardus pardalis) in the Atlantic Forest of Misiones. Argent J Zool. 2006;270:153–163.
- Mosquera D, Blake JG, Romo D, et al. Ocelot (Leopardus pardalis) density in Eastern Ecuador based on capture–recapture analyses of camera trap data. Neotrop Biodivers. 2016;2:51–58.10.1080/23766808.2016.1168593
- Karanth KU, Nichols JD, Kumar NS, et al. Assessing tiger population dynamics using photographic capture–recapture sampling. Ecology. 2006;87:2925–2937.10.1890/0012-9658(2006)87[2925:ATPDUP]2.0.CO;2
- Blake JG, Mosquera D, Guerra J, et al. Yasuní – a hotspot for jaguars Panthera onca (Carnivora: Felidae)? Camera-traps and jaguar activity at Tiputini Biodiversity Station. Ecuador Rev Biol Trop. 2014;62:689–698.
- Blake JG, Mosquera D, Loiselle BA, et al. Spatial and temporal activity patterns of ocelots Leopardus pardalis in lowland forest of eastern Ecuador. J Mammal. 2016;97:455–463.10.1093/jmammal/gyv190
- Weckel M, Giuliano W, Silver S. Jaguar (Panthera onca) feeding ecology: distribution of predator and prey through time and space. J Zool. 2006;270:25–30.
- Harmsen BJ, Foster RJ, Silver S, et al. Differential use of trails by forest mammals and the implications for camera-trap studies: a case study from Belize. Biotropica. 2010;42:126–133.10.1111/j.1744-7429.2009.00544.x
- Cusack JJ, Dickman AJ, Rowcliffe JM, et al. Random versus game trail-based camera trap placement strategy for monitoring terrestrial mammal communities. PLOS One. 2015;10:e0126373.10.1371/journal.pone.0126373
- Blake JG, Mosquera D. Camera trapping on and off trails in lowland forest of eastern Ecuador: does location matter? Mastozool Neotrop. 2014;21:17–26.
- Bass MS, Finer M, Jenkins CN, et al. Global conservation significance of Ecuador's Yasuní National Park. PLoS One. 2010;5:e8767.10.1371/journal.pone.0008767
- Minesterio del Ambiente. Plan de Manejo del Parque Nacional Yasuní [Management Plan for Yasuni National Park]. Quito: Ecuador; 2011.
- Blake JG, Loiselle BA. Enigmatic declines in bird numbers in lowland forest of eastern Ecuador may be a consequence of climate change. PeerJ. 2015;3:e1177.10.7717/peerj.1177
- Blake JG, Mosquera D, Guerra J, et al. Mineral licks as diversity hotspots in lowland forest of eastern Ecuador. Diversity. 2011;3:217–234.10.3390/d3020217
- Rovero F, Marshall AR. Camera trapping photographic rate as an index of density in forest ungulates. J Appl Ecol. 2009;46:1011–1017.10.1111/jpe.2009.46.issue-5
- Software Analytical. Statistix 10. Tallahassee (FL): Analytical Software; 2013.
- Kerley LL, Goodrich JM, Miquelle DG, et al. Effects of roads and human disturbance on Amur tigers. Conserv Biol. 2002;16:97–108.10.1046/j.1523-1739.2002.99290.x
- Johnson A, Vongkhamheng C, Hedemark M, et al. Effects of human-carnivore conflict on tiger (Panthera tigris) and prey populations in Lao PDR. Anim Conserv. 2006;9:421–430.10.1111/acv.2006.9.issue-4
- Salvador J. Effects of human disturbance on occurrence patterns and behavior of terrestrial mammals in eastern Ecuador [masters dissertation]. Gainesville (FL): University of Florida; 2015.
- Blake JG, Mosquera D, Salvador J. Use of mineral licks by mammals and birds in hunted and non-hunted areas of Yasuní National Park, Ecuador. Anim Conserv. 2013;16:420–437.
- Di Bitetti MS, Paviolo A, Ferrari CA, et al. Differential responses to hunting in two sympatric species of brocket deer (Mazama americana and M. nana). Biotropica. 2008;40:636–645.10.1111/btp.2008.40.issue-5