
Abstract
We provide information about the composition of the terrestrial frogs of the genus Pristimantis in the upper basin of the Upano River, located on the Eastern slope of the Andes Mountain range in the Morona Santiago province, south-east of Ecuador. The compendium of the 37 species comes from collections conducted on an altitudinal gradient of between 1170 to 3750 meters. Four species are described as new: Pristimantis albujai sp. nov, P. churuwiai sp. nov, P. sambalan sp. nov and P. saturtinoi sp. nov. Two species are reported for the first time in Morona Santiago, expanding their known distribution. Eight species have an undetermined taxonomic status, which could represent new additions to Ecuador’s fauna. Of the 28 species formally described, 16 are endemic to the eastern slopes of the Ecuadorean Andes, and eight are restricted to the upper sub-basin of the Upano River. The highest concentration of diversity occurs between 1200 to 1600 meters of elevation, diminishing as the altitude increases. Our survey represents 45.5% of the Pristimantis frogs reported for the upper basin of the Napo, Pastaza and Santiago rivers.
Se proporciona información sobre la composición de las ranas terrestres Pristimantis de la subcuenca alta del río Upano ubicada en la vertiente oriental de la Cordillera de los Andes, provincia de Morona Santiago, suroriente de Ecuador. El compendio de las 37 especies proviene de colecciones realizadas en una gradiente altitudinal comprendida entre 1170 a 3750 m de elevación. Cuatro especies son descritas como nuevas: Pristimantis albujai sp. nov, P. churuwiai sp. nov, P. sambalan sp. nov y P. saturninoi sp. nov. Dos especies: Pristimantis bicantus y P. inusitatus, amplían su distribución y son citadas por primera vez para Morona Santiago. Ocho especies presentan estatus taxonómico que requiere revisión, las cuales pueden representar nuevas adiciones en la fauna de Ecuador. De las 28 especies formalmente descritas, 16 son endémicas de las laderas orientales de los Andes de Ecuador, y ocho están restringidas a la subcuenca alta del Upano. La mayor concentración de diversidad ocurre entre los 1200 a 1600 m de elevación, menguando a medida que aumenta la altitud. Nuestro compendio representa el 45.5% de las ranas Pristimantis reportadas para la Cuenca alta de los ríos Napo, Pastaza y Santiago.
Introducción
Con 505 especies, el género Pristimantis es el más diverso de entre los anfibios [Citation1], siendo en Ecuador el género más especiado y con mayor número de especies endémicas, 194 y 106 especies respectivamente [Citation2]. Su extraordinaria radiación adaptativa en los Andes, es atribuida a su peculiar sistema reproductivo de desarrollo directo, sumado a las barreras geográficas y adaptabilidad a gran variedad de hábitats [Citation3,4]. La mayor concentración de especies de ranas Pristimantis ocurre en los bosques andino montanos [Citation2,3,5], sin embargo, la creciente fragmentación de los hábitats naturales ha puesto en peligro de extinción a numerosas especies [Citation6–8].
Los bosques inmersos en la subcuenca alta del río Upano (SARU) circunscriben a las laderas orientales de los Andes ecuatorianos, resguardando una importante biodiversidad de especies tanto florísticas como faunísticas [Citation9–11]. Expediciones realizadas en los últimos años a ésta región escasamente estudiada, han dado a conocer un creciente número de vertebrados endémicos restringidos a estos ecosistemas [Citation12–17]. Todos estos descubrimientos provienen del interior del Parque Nacional Sangay (PNS), área nominada por la UNESCO en 1983 como Patrimonio Natural de la Humanidad.
Durante los años 2010 al 2016, nuestro grupo de trabajo realizó numerosas expediciones a la SARU, en zonas de amortiguamiento y en el interior del PNS, obteniendo material significativo de ranas terrestres Pristimantis, que ha permitido construir una sinopsis del género en la región, así como identificar especies nuevas y determinar los patrones de diversidad de éstos pequeños vertebrados.
Métodos
Evaluamos ocho localidades de la SARU, previamente seleccionadas, en un rango de elevación altitudinal de 1150 y 3750 m (Figura ), durante el periodo 2010–2016. El esfuerzo de muestreo en cada uno de los sitios fue variable, de tres a 12 días por sitio, con promedio de 500 m de recorrido por punto, influyendo en el esfuerzo las distancias de caminata, severidad del clima, irregularidad topográfica y el número de colectores. En los sitios con mayor dificultad de acceso se empleó menor esfuerzo de muestreo. La metodología para el registro de los anuros sigue a [Citation18–20]. Las coordenadas, rango altitudinal, tipo de ecosistema y esfuerzo de captura se anotan en la Tabla .
Figura 1. Mapa del área de estudio. (1) Domono; (2) Danu; (3) río Sardinayacu; (4) Sardinayacu; (5) Guabisai; (6) Tinguichaca; (7) Cerro Sambalan; (8) Cisnian.
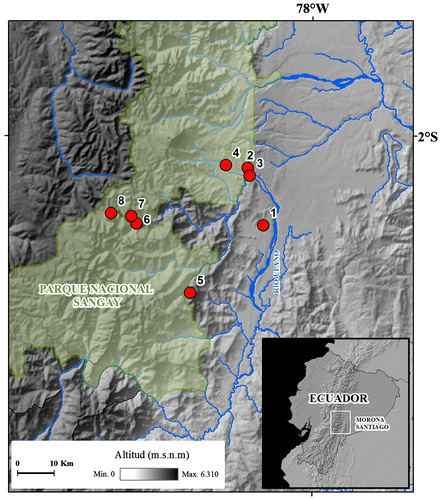
Tabla 1. Detalles de las localidades de estudio en la SARU, Ecuador. (BMB) Bosque Montano Bajo; (BNM) Bosque de Neblina Montano; (BMA) Bosque Montano Alto.
Análisis de diversidad
Los patrones de alfa–beta diversidad y la distribución espacial de la riqueza fueron analizados a partir de las amplitudes altitudinales de las especies determinadas en el estudio. La riqueza de especies, taxa únicos, especies endémicas y amenazadas, fueron distribuidas en seis rangos altitudinales desde los 1200 m a 3600 m, separados entre sí cada 400 m de ancho de banda de elevación. Usando estadística descriptiva, con la ayuda del programa Past® 3.12, [Citation21] se realizaron histogramas para graficar la amplitud altitudinal de las especies y la distribución de riqueza en la gradiente. Para determinar el grado de influencia de la elevación sobre la riqueza de Pristimantis, se aplicó una prueba de regresión lineal simple entre la elevación y el logaritmo natural de la riqueza. A partir de datos de presencia–ausencia de las especies en los seis rangos de altitud, se aplicó un análisis de conglomerados (Cluster) basado en el algoritmo de Jaccard, para determinar el grado de similitud entre los rangos de elevación, también en Past® 3.12. [Citation21] De igual forma bajo el mismo análisis, se compararon los datos de riqueza de la Cuenca del río Santiago, frente a la Cuenca del Pastaza y del Napo.
Nomenclatura
La terminología del género sigue a Hedges et al. [Citation22] y para la familia seguimos a Pyron and Wiens [Citation23] y Padial et al. [Citation24]. Para la descripción de las especies se adopta la terminología estandarizada de Lynch and Duellman [Citation3], Rueda and Lynch [Citation25] y Duellman and Lehr [Citation5].
Morfología
Las medidas se tomaron con un calibrador digital Buffalo Tools® (precisión de ± 0.01 mm). Los caracteres morfológicos utilizados son: (1) Longitud rostro–cloaca; (2) distancia órbita–narina (desde el margen anterior de la órbita hasta el margen posterior de la narina; (3) longitud de la cabeza (desde el borde posterior del escamoso hasta la punta del hocico); (4) ancho cefálico (medido a nivel de las mandíbulas posteriores); (5) distancia interorbital (entre bordes proximales de los párpados); (6) distancia internarial (distancia entre los bordes proximales de las narinas); (7) longitud de la tibia (desde la rodilla flexionada hasta el talón); (8) longitud del pie (tomado desde el margen proximal del tubérculo metatarsal interno hasta la punta del disco del dedo IV); (9) longitud de la mano (distancia desde el borde proximal del tubérculo palmar interno a la punta del dedo III); (10) diámetro horizontal del tímpano; (11) longitud horizontal del ojo; (12) ancho del párpado. El sexo de los especímenes fue determinado por inspección de las gónadas a través de disecciones para observar testículos u oviductos y comprobación de presencia/ausencia de hendiduras vocales.
Los especímenes colectados fueron sacrificados según las recomendaciones de Chen and Combs [Citation26] y preservados según los protocolos de Simmons [Citation27]. La coloración en vida fue tomada de las notas de campo y fotografías a color, siguiendo la terminología de Encicolorpedia [Citation28]. El material examinado (Apéndice 1) y colectado, se encuentra depositado en la sección de Herpetología del Museo Ecuatoriano de Ciencias Naturales (DHMECN), en el Museo de Historia Natural Gustavo Orcés de la Escuela Politécnica Nacional (MEPN) y en el Museo de Zoología de la Pontificia Universidad Católica del Ecuador (QCAZ).
Vocalizaciones
Los cantos se registraron con una grabadora digital Olympus® WS–822 conectada a un sistema modular Sennheiser® K6–C y acoplada a un micrófono de cabezal Sennheiser® (ME 66) 40–20000 Hz ± 2,5dB, a una distancia de grabación de 3 m; utilizando en la digitalización, filtros de ruido y análisis temporales en el programa Adobe Audition® CS6 [Citation14]. Para las diagramaciones y análisis espectrales se utilizó el programa Raven® 1.4 [Citation29], a una frecuencia de muestreo de 44.1 kHz y 16 “bits” de resolución, utilizando la ventana de Hann al 80% de superposición y 512 puntos de resolución de la transformación rápida de Fourier (FFT).
Los parámetros que se analizaron fueron: (1) Frecuencia dominante; (2) Armónicos; (3) Duración de los cantos; (4) Intervalos entre cantos; (5) cantos/minuto; (6) Notas/canto; (7) Duración de las notas; (8) Intervalos entre notas; (9) Notas/segundo. Las definiciones y mediciones realizadas en los parámetros acústicos, se siguieron según los términos de Batallas and Brito [Citation14], Duellman and Pyles [Citation30], Cocroft and Ryan [Citation31], Angulo et al. [Citation32] y Díaz and Cádiz [Citation33].
Resultados
Distribución espacial de la diversidad de Pristimantis en la SARU
Reportamos que la Cuenca del río Upano contiene 37 especies de ranas terrestres Pristimantis en una gradiente de elevación que va desde los 1200 a 3600 m (Figura (A)). De las cuales 25 especies han sido previamente reportadas en los bosques andinos de los Andes orientales de Ecuador, cuatro son descritas como nuevas y ocho taxones son identificados como especies candidatas nuevas. 16 especies son endémicas y 9 están amenazadas.
Figura 2. Distribución espacial de alfa-diversidad de ranas Pristimantis de la subcuenca del Upano. (A) Bandas de amplitud altitudinal de 36 especies de Pristimantis. (BMB) Bosque Montano Bajo; (BNM) Bosque de Neblina Montano; (BMA) Bosque Montano Alto. (B) Histograma de valores de riqueza (S′) de ranas Pristimantis en seis rangos altitudinales. (N°S′) número de especies; (S′ únicas) número de especies únicas del rango; (S′ endémicas) número de especies endémicas; (S′ amenazadas) número de especies amenazadas.
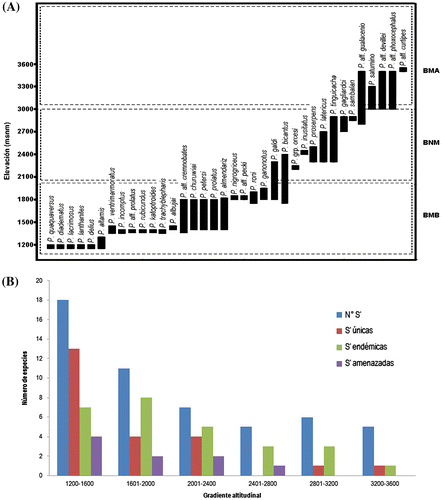
La diversidad en la gradiente se compone de tres diferentes tipos de ensamblajes (Figura (A)) asociados a los siguientes ecosistemas: (1) Bosque Montano Bajo: contiene 25 especies, entre los 1200 a 2000 m de altura; (2) Bosque de Neblina Montano: con 10 especies, en elevaciones de 2100 a 3000 m; y (3) Bosque Montano Alto: 5 especies, sobre los 3100 m.
En promedio, las ranas terrestres Pristimantis exhiben bajas amplitudes de distribución altitudinal; cerca del 47% (que equivale a 16 especies) de las especies no superan amplitudes de 50 m de elevación; un porcentaje similar 44% (15 especies) tienen anchos de banda de 500 m, y tan solo tres especies alcanzan amplitudes de banda de hasta 700 m. Resalta en la gradiente altitudinal, que el mayor número de especies únicas por rango (15 especies), se concentre entre 1200 a 1600 m de elevación y disminuya drásticamente a partir de 1800 m.
Todos los rangos altitudinales contienen un promedio del 50% de especies endémicas, destacando principalmente la banda de 1601 a 2000 m con un máximo de ocho taxa. Entre los 1200 a 2800 m de altura se concentran especies de Pristimantis categorizadas en riesgo de extinción, alcanzando un promedio de 22% de especies amenazadas por banda altitudinal (Figura (B)).
La elevación, influencia significativamente la diversidad de ranas Pristimantis de la SARU (Figura (A)). Bajo una tendencia inversamente proporcional, la riqueza de especies se incrementa a medida de que el rango de elevación disminuye (Figura (A)). La cota de menor elevación en la cuenca (1200 a 1600 m de altura), alcanza el mayor valor de alfa diversidad de ranas Pristimantis (19 especies), comprimiéndose en un 39% sobre los 1600 m de elevación; reduciendo gradualmente su riqueza hasta alcanzar cinco especies sobre los 3200 m de altura (Figuras (B) and 3(A)).
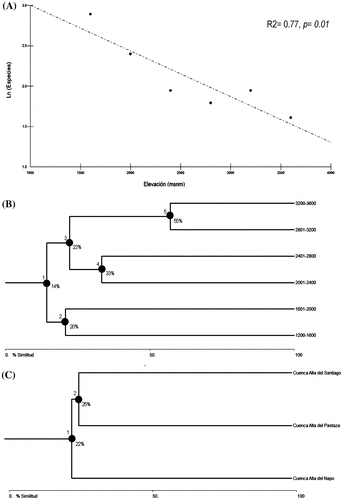
Figura 3. Beta diversidad de ranas Pristimantis de la subcuenca del Upano. (A) Regresión lineal entre la elevación y la riqueza de especies (valores de significancia se exhiben en la parte superior derecha). (B) Análisis de conglomerados (cluster) basado en el algoritmo de Jaccard para riqueza de especies en seis rangos altitudinales (los agrupamientos se muestran con círculos negros); (C) Análisis de conglomerados (cluster) basado en el algoritmo de Jaccard para riqueza de especies en tres cuencas de las estribaciones de los Andes orientales de Ecuador (los agrupamientos se muestran con círculos negros).
La diversidad del género Pristimantis en la gradiente altitudinal de la SARU es heterogénea y sus ensamblajes son especializados a los tres ecosistemas identificados en la región. Determinamos que el 64% de las especies tienen baja frecuencia a lo largo de la gradiente altitudinal, ya que están presentes en un solo rango de los seis delimitados; un 28% de la composición cubre hasta dos rangos de elevación, y un reducido 8% de especies se distribuye en tres rangos (Figura (B)).
El análisis de conglomerados determinó cinco agrupamientos para los seis rangos altitudinales estudiados (Figura (B)): (1) el primer agrupamiento determina que todos los rangos comparten tan solo el 14% de la composición de especies; (2) agrupa a los rangos altitudinales de las comunidades de bosque montano bajo, entre los 1600 a 2000 m, los cuales comparten el 20% de sus especies; (3) asocia los ensamblajes de bosque montano y bosque montano alto, sobre los 2000 m hasta los 3600 m de elevación, compartiendo el 22% de su composición; (4) el agrupamiento se compone por los rangos altitudinales del bosque montano sobre los 2000 m hasta los 2800 m de elevación, los dos ensambles comparten el 33% de sus especies; (5) el último agrupamiento asocia a las cotas de elevación sobre los 2800 hasta los 3600 m, compartiendo el porcentaje más alto de composición de especies (55%) en toda la gradiente.
A escala regional (Apéndice 2), la riqueza de ranas terrestres Pristimantis de la zona de estudio, muestra una baja similitud en la composición de especies en relación a otras dos importantes cuencas de las estribaciones orientales de los Andes de Ecuador (Figura (C)), alcanzando el 22% de similitud entre ellas, y asociándola cercanamente a la región de la Cuenca del Pastaza al compartir un cuarto (25%) de su riqueza de especies.
Compendio de especies
Pristimantis albujai sp nov
Nombre común en español: Cutín de Albuja
Common name in English: Albuja’s Rainfrog
Holotipo (Figuras 4–6)
Hembra adulta DHMECN 12245, colectado por DBR, JBM, Hernán Orellana y Jenny Curay el 18 de septiembre del 2015 en el río Sardinayacu (2º04′45.3″S; 78º09′37.3″W, 1400 m), Parque Nacional Sangay, parroquia Sinaí, cantón Morona, provincia de Morona Santiago, República del Ecuador.
Etimología
El epíteto específico “albujai” es un patronímico en homenaje al eminente mastozoólogo ecuatoriano Luís H. Albuja Viteri, quien ha aportado sustancialmente al conocimiento de los mamíferos neotropicales, ha formado e influenciado a numerosos biólogos ecuatorianos y ha contribuido significativamente al conocimiento de los vertebrados de Ecuador.
Diagnóstico
Una especie de Pristimantis caracterizada por: (1) Piel del dorso finamente granular con verrugas bajas y aplanadas, pronunciadas hacia la parte posterior del dorso y los flancos; vientre levemente areolado y con verrugas elevadas; pliegues dorsolaterales presentes, extendiéndose por detrás de los ojos hasta la cintura pélvica; pliegue discoidal presente bien definido; (2) membrana timpánica y anillo timpánico presentes; (3) hocico corto, subacuminado en vista dorsal, redondeado de perfil; (4) un tubérculo cónico grande sobre el párpado superior y rodeado de varios tubérculos aplanados pequeños; distancia inter-orbital menor que la longitud de la órbita; (5) procesos dentígeros del vómer presentes, oblicuos de contorno y con 1-2 dientes; (6) hendiduras vocales y almohadillas nupciales desconocidos; (7) dedo manual I más corto que el II, discos truncados y con almohadillas dilatadas; (8) dedos manuales con rebordes cutáneos laterales; (9) tubérculos ulnares presentes cónicos y medianos; (10) talón con un tubérculo cónico distintivo; borde externo del tarso con un tubérculo cónico y tres redondeados, pliegue tarsal interno débilmente definido; (11) tubérculo metatarsal interno oval 2.5 veces del tamaño del tubérculo metatarsal externo, redondeado; (12) dedos del pie con rebordes cutáneos laterales, fuertemente definidos en la base de los dedos; membrana interdigital ausente; dedo V mucho más largo que el III; discos del pie de similar tamaño que los de las manos, dedo V alcanza el tubérculo subarticular distal del dedo IV; (13) dorsalmente de aspecto marrón grisáceo con distintivos pliegues dorsolaterales crema (marrón cobrizo en vida), con manchas café oscuras delineadas y encerradas por un borde crema, vientre en fondo marrón oscuro con verrugas elevadas de color café oscuro, garganta en fondo café claro con marca en forma de V en el mentón, color café oscuro; ingles, superficies anteriores de los muslos y piernas crema blanquecino (rojizo en vida); iris café con una distintiva banda media con tonalidad café oscura (14) adultos de tamaño mediano, LRC en machos desconocido, LRC hembra (DHMECN 12245) 18.1 mm, n = 1.
Comparaciones
Dentro del género de ranas Pristimantis de la ladera Oriental de los Andes de Ecuador, Pristimantis albujai sp. nov., forma parte del ecotipo de ocho ranas espinosas [Citation15], que presentan conspicuos tubérculos en el párpado superior, talón, borde externo del tarso, antebrazo y superficies ocultas de ingle incluyendo muslos con coloraciones llamativas. Sin embargo este conjunto de ocho especies (Pristimantis roni, P. katoptroides, P. galdi, P. eriphus, P. inusitatus, P gualacenio, P. bellae y P. rufoviridis), exhiben coloraciones del dorso predominantemente verde; las cuales contrastan con la coloración dorsal café oscuro con distintivas bandas dorsolaterales marrón cobrizo en P. albujai, así como, la presencia de procesos dentígeros del vómer oblicuos (triangulares en las ochos especies citas). Tamaño corporal máximo en hembras <22.5 mm de P. albujai (tamaños máximos >23 mm en las ocho especies citadas).
Las únicas especies con las que podría ser confundido P. albujai sp. nov., es P. verecundus [Citation34] y P. mutabilis [Citation35] de la ladera Occidental de los Andes, distribuidas entre el suroccidente de Colombia y el noroccidente de Ecuador [Citation3,34–36]; las tres especies tienen tamaños corporales pequeños (con un LRC de hasta 23.2 mm), tubérculos cónicos en el párpado, talón, tarso y distintiva coloración inguinal roja. No obstante, P. verecundus difiere de P. albujai (caracteres de ésta especie entre paréntesis) por presentar vomerinos odontóforos subtriangulares de contorno y prominentes (oblicuos y bajos), dedos de las manos con quillas (con rebordes cutáneos bien definidos y extendidos en la base de los dedos), flancos con distintivas barras transversales (sin barras transversales) y pliegues dorsolaterales que se extienden desde la región escapular hasta la región del sacro (se extienden desde la región posterior de los ojos hasta el sacro).
Descripción del holotipo
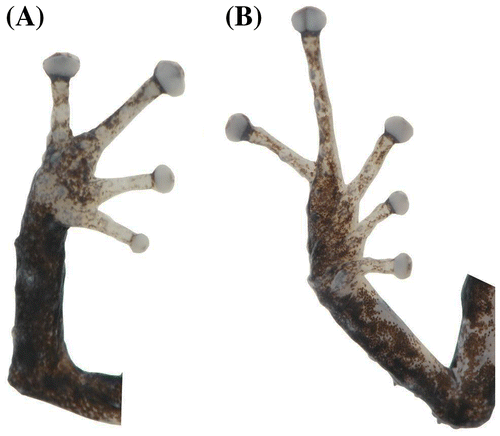
Hembra adulta con la cabeza ligeramente más larga que ancha, 133%; hocico corto, subacuminado en vista dorsal y redondeado de perfil; longitud horizontal de la órbita mucho mayor que la distancia órbita-narina, 260%; narinas protuberantes y dirigidas lateralmente; canto rostral cóncavo; región loreal redondeado; labios ligeramente acampanados; tubérculo cónico grande en el párpado superior y rodeado de varios tubérculos aplanados pequeños; crestas craneales ausentes; un fino pliegue supratimpánico extendido desde la región posterior del ojo hasta el borde del segundo tubérculo postrictal; varios tubérculos postrictales subcónicos detrás del tímpano y hacia la comisura con la mandíbula; membrana timpánica presente; anillo timpánico visible dorsalmente, redondeado, su longitud es el 33.3% de la longitud horizontal del ojo; coanas pequeñas y redondeadas de contorno; procesos dentígeros del vómer presentes, oblicuos de contorno, pequeños y con bordes regulares, con 1–2 dientes; lengua más larga que ancha, con el borde posterior unilobular, el 25% está adherida al piso de la boca. Piel del dorso finamente granular con verrugas bajas y aplanadas, pronunciadas hacia la parte posterior del dorso y los flancos; pliegues dorsolaterales presentes y elevados (reducidos por efectos de preservación), que se extienden desde la parte posterior del ojo hasta la región del sacro; vientre levemente areolado y con verrugas elevadas; pliegue discoidal presente bien definido que se extiende desde las axilas hasta la parte posterior de la ingle; cubierta cloacal mediana; tubérculos cloacales presentes y dispersos. Tubérculos escapulares presentes, subcónicos y pequeños; tubérculos cónicos medianos y esparcidos en la región ulnar y el codo. Superficie palmar mediana; tubérculo palmar bífido y ligeramente elevado (Figura (A)), tubérculo tenar pequeño y ovalado, aproximadamente 1/2 del ancho del tubérculo palmar; tubérculos supernumerarios agrandados, elevados y dispersos; tubérculos subarticulares pequeños, redondeados y ligeramente elevados; dedos manuales con rebordes cutáneos laterales, sin membranas interdigitales; dedo I más corto que el dedo II; disco del dedo I ligeramente expandido, discos de los dedos II–IV expandidos, redondeados y ligeramente más expandidos que los dígitos (Figura (A)); todos los dedos manuales con cojinetes ventrales definidos por hendiduras circunmarginales. Extremidades posteriores delgadas, longitud de la tibia 52.4% de la longitud rostro-cloaca; talón con un tubérculo cónico distintivo; borde externo del tarso con un tubérculo cónico y tres redondeados; tubérculo metatarsal interno ovalado, mediano, elevado y alargado, aproximadamente 2.5 veces más grande que el tubérculo metatarsal externo; pliegue tarsal débilmente definido en el primer tercio del borde interno del tarso; tubérculos supernumerarios plantares pequeños y dispersos, ligeramente elevados de perfil; tubérculos subarticulares pequeños, redondeados y elevados; dedos del pie con fuertes rebordes cutáneos laterales, pronunciados hacia la base de los dedos; sin membranas interdigitales; discos de los dedos del pie ligeramente más pequeños que los discos de los dedos de la mano, todos los discos llevan hendiduras circunmarginales; longitud relativa de los dedos del pie: I < II < III < V < IV; dedo III del pie más pequeño que el dedo V; dedo III del pie llega al borde anterior del tubérculo subarticular distal del dedo IV (Figura (B)).
Figura 4. Pristimantis albujai sp. nov. (DHMECN 12245). (A) Vista palmar de la mano; (B) vista plantar del pie. Longitud del pie 7.9 mm.
Coloración del holotipo en preservado
Dorsalmente de aspecto marrón grisáceo con distintivos pliegues dorsolaterales crema, en la región medial con manchas café oscuras delineadas y encerradas por finas líneas crema, región interorbital con dos manchas café oscuras; barra supratimpánica café oscuro, con una fina línea blanca delineando el borde inferior del tímpano, con dos barras diagonales al ojo de color blanco y café oscuro; extremidades anteriores y posteriores en fondo crema blanquecino con barras transversales café oscuro, superficies de los dedos de las manos I y II y de los dedos del pie I–IV crema (Figura (A)); vientre en fondo marrón oscuro con verrugas elevadas de color café oscuro, garganta en fondo café claro con marca en forma de “V” en el mentón color café oscuro (Figura (B)), superficies palmares, plantares y dígitos pigmentados de blanco; ingles, superficies anteriores de los muslos y piernas, crema blanquecino.
Figura 5. Pristimantis albujai sp. nov., en preservado. (A) vista dorsal; (B) vista ventral del holotipo DHMECN 12245, macho adulto, LRC: 18.1 mm.

Coloración del holotipo en vida
Dorso marrón oscuro, con tonalidades sepia hacia la región sacra; bandas marrón cobrizo (tipo windsor tan) definidas hacia el limite externo de la superficie dorsal y extendiéndose desde el sacro hasta la base de la región orbital; maculaciones negruzcas, prominentes, con bordes almendra, que van desde la región sacra hasta la región interorbital. Región rostral de la misma coloración que el dorso, siendo más oscura el área de las narinas y canto rostral; bandas labiales marrón oscura y crema a nivel infraocular. Párpado superior marrón oscuro, con tonos dorados (tipo golden brown) en el tubérculo supraocular. Superficie dorsal de las extremidades anteriores y posteriores marrón claras, con barras transversales marrón oscuras, más evidentes a nivel del muslo y antebrazo. Miembros manuales y pediales de la misma coloración que las extremidades, con dedos dorado pálido y manchas marrón oscuras en los dedos manuales III–IV. Vientre negruzco de tonalidades marrones, con leves puntos blancos y barras marrón claras hacia la mandíbula; región posteroventral hacia los flancos de tonos rojizos (tipo rojo sangría). Superficie ventral de las extremidades anteriores y posteriores de la misma coloración del vientre, exceptuando el borde externo de los muslos y piernas que son rojizos; manchas marrón oscuras, con bordes amarillentos hacia el borde interno de las piernas (Figura ). Superficies palmar y plantar marrón oscuro, ligeramente más claros que sus superficies dorsales. Iris café con una distintiva banda media con tonalidad café oscura.
Figura 6. Pristimantis albujai sp. nov., en vida. Vista dorsal y ventral del holotipo DHMECN 12245, macho adulto, LRC: 18.1 mm.

Figura 7. (A) Pristimantis almendariz ♀, LRC 34.6 mm, DHMECN 12363; (B) P. altamnis ♂, LRC 17.7 mm, DHMECN 12214; (C) P. bicantus, ♀, LRC 22 mm: MEPN 14454; (D) P. conspicillatus complex ♂, LRC 32.6, MEPN 14675; (E) P. af. cremnobates, ♀, LRC 23.3 mm, MEPN 14370; (F) P. af. curtipes, ♂, LRC 39.4 mm, DHMECN 12305.

Medidas del holotipo (mm). Longitud rostro-cloaca = 18.1; distancia órbita-narina = 1.5; longitud cefálica = 8.8; ancho cefálico = 6.6; distancia interorbital = 2.2; distancia internarinal = 3.3; longitud de la tibia = 9.5; longitud del pie = 7.9; longitud de la mano = 4.9; diámetro del tímpano = 1.3; longitud horizontal del ojo = 3.9; ancho del párpado = 2.2.
Distribución y ecología
La especie nueva es conocida solamente de la localidad tipo, en el río Sardinayacu dentro del Parque Nacional Sangay, provincia de Morona Santiago, Ecuador, a una elevación de 1400 msnm (Figura ). El piso zoogeográfico donde habita Pristimantis albujai, pertenece al Piso Subtropical Oriental [Citation37], el ecosistema corresponde al Bosque siempreverde piemontano del sur de la Cordillera Oriental de los Andes [Citation38]. El hábitat se caracteriza por la presencia de árboles con fustes cargados de abundantes musgos y lianas; en general la altura de la vegetación emergente alcanza hasta 35 m de altura, donde dominan visualmente especies como el Copal (Dacryodes peruviana Loes), la palma real (Dictyocaryum lamarckianum Mart.) y el canelón (Ocotea sp); el sotobosque es denso, dominado por platanillos (Heliconaceae), joyapas (Ericaceae) y variedad de arbustos (Asteraceae y Melastomataceae). Pristimantis albujai fue colectado en las primeras horas de la noche (19:30), en el interior de un bosque prístino perchado sobre una hoja a 0.3 m sobre el suelo. Fue colectado en simpatría con Pristimantis almendariz, P. katoptroides, P. petersi y P. rubicundus.
Comentarios taxonómicos
Pristimantis albujai presenta una morfología externa y patrón de coloración muy similar a P. verecundus de las estribaciones Occidentales de los Andes de Colombia y Ecuador, recientemente Guayasamin et al. [Citation35] reasignaron a P. verecundus dentro del grupo de especies de P. myersi, por ende, nosotros asignamos a P. albujai tentativamente al grupo de especies de P. myersi por presentar los siguientes caracteres: tamaño corporal <27 mm, cabeza ancha y corta, ausencia de cresta craneal, cuerpo robusto, extremidades moderadamente largas, piel del vientre aereolado, vomerinos oblicuos, tímpano presente, y por el dedo V del pie ligeramente más pequeño que el dedo III. Aunque su ecotipo [Citation15], corresponde a las ranas espinosas de los Andes Orientales, consideramos que es improbable que esté asociado a tres de los cuatro linajes parafiléticos propuestos para este ecotipo, y sugerimos una estrecha relación con el grupo de especies de P. myersi. La implementación de futuros análisis moleculares para esta especie, podrán apoyar o descartar nuestra hipótesis sobre la relación de P. albujai con el grupo P. myersi.
Pristimantis almendariz [Citation39] (Figura 7(A))
Especie endémica de Ecuador, recientemente descrita de los bosques piemontanos de la Cordillera de Kutukú, entre 1400 a 1820 [Citation39]. En la SARU se registraron individuos provenientes del río Sardinayacu, correspondiendo la segunda localidad para la especie y ampliando su rango latitudinal de distribución. Preliminarmente ha sido asignado al grupo de especies de Pristimantis ridens [Citation24,39]. En la localidad del río Sardinayacu, P. almendariz es un habitante del bosque piemontano entre los 1400 y 1500 m de altitud. Prefiere el estrato bajo del bosque entre los 0.5 y 1.5 m. En Ecuador se lo considera una especie sin datos para evaluar su amenaza [Citation6], mientras que a nivel global la UICN [Citation8] no la ha evaluado.
Material de referencia: DHMECN 12363–66.
Pristimantis altamnis [Citation40] (Figura 7(B))
Especie endémica de Ecuador, circunscrita a los bosques tropicales de la Amazonía entre 400 a 1000 [Citation2,40]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies, aunque Elmer and Canatella [Citation40] la identifican cercanamente relacionada con el complejo de especies de P. ockendeni. En la localidad de Domono, P. altamnis es un habitante del bosque piemontano a 1150 m de altitud, prefiere el estrato bajo del bosque entre los 0.3 y 0.7 m. En Ecuador es categorizada como una especie Vulnerable [Citation6] y globalmente [Citation8] es considerada como en Riesgo menor.
Material de referencia: DHMECN 12214.
Pristimantis bicantus [Citation41] (Figura 7(C))
Especie endémica de Ecuador, conocida de dos localidades en los bosques nublados de la Estación Biológica Yanayacu y la cuenca alta del río Pastaza, entre 2000 a 2300 [Citation41,42]. Guayasamin and Funk [Citation41] y Padial et al. [Citation24] lo asignan al grupo de especies Pristimantis myersi. En la localidad de Sardinayacu y Guabisai, P. bicantus es un habitante del bosque montano entre 1750 y 2400 m de altitud, prefiriendo el estrato bajo donde permanecen activos durante el día y en el crepúsculo. En diciembre del 2010 dos hembras grávidas fueron registradas, contenían entre 22 y 25 ovocitos de color crema. En Ecuador [Citation6] y a nivel global [Citation8] la especie no ha sido evaluada.
Material de referencia: DHMECN 12162–4; MEPN 14452–6, 14458.
Pristimantis conspicillatus complex [Citation42] (Figura 7(D))
Especie que se distribuye en la vertiente oriental de los Andes de Colombia, Ecuador, Perú y Brasil [Citation2,43], en Ecuador habita en todas las provincias de la Amazonía, entre 690 y 2050 m. [Citation2] Padial et al. [Citation24] lo asignan al grupo de especies Pristimantis conspicillatus. Reyes-Puig et al. [Citation42] comentaron su estatus taxonómico determinando que las poblaciones en la región son un complejo de especies del grupo conspicillatus. En la localidad Domono y río Sardinayacu, P. conspicillatus es un habitante del bosque piemontano prístino y áreas alteradas como cultivos y bordes, a 1150 y 1400 m de altitud. Habita la vegetación herbácea entre 0.3 y 0.8 m de altura, perchando sobre hojas o ramas pequeñas. En Ecuador [Citation6] y a nivel global [Citation8] es categorizada como una especie en Preocupación Menor.
Material de referencia: MEPN 14675; DHMECN 12264-69.
Pristimantis churuwiai sp nov
Nombre propuesto en español: Cutín de Churuwia
Proposed common name in English: Chruruwia’s Rainfrog
Holotipo (Figuras 8–10)
Hembra adulta DHMECN 12242, colectada por DBR, JBM, Hernán Orellana y Jenny Curay el 18 de septiembre del 2015 en el río Sardinayacu (2º04′45.3″S; 78º09′37.3″O, 1400 m), Parque Nacional Sangay, parroquia Sinaí, cantón Morona, provincia de Morona Santiago, República del Ecuador.
Paratopotipos
Hembra adulta DHMECN 12243, con los mismos datos del holotipo.
Paratipos
Hembras adultas: DHMECN 12244, colectada por JBM, Hernán Orellana y Germán Tenecota el 16 de junio del 2014 en Sardinayacu (2°04′20.5″S, 78°12′52.4″W, 1800 m), Parque Nacional Sangay, parroquia Sinaí, cantón Morona; MEPN 14497, colectada por JBM el 25 de febrero del 2011; y MEPN 15040–1, 15047, colectadas por JBM y Glenda Pozo el 26 de junio del 2012 en Yapit (2°43′14.3″S, 78°05′17.0″W, 1581 m), Bosque Protector Kutukú-Shaimi, parroquia Shimpis, cantón Logroño, provincia de Morona Santiago, Ecuador.
Etimología
El epíteto específico hace referencia al guía naturalista del Parque Nacional Sangay Marcelo Sharup más conocido como Churuwia, en reconocimiento por su continuo esfuerzo por la conservación de los bosques del área de amortiguamiento del PNS.
Diagnóstico
Una especie de Pristimantis caracterizada por: (1) Piel del dorso finamente granular; vientre areolado; pliegues dorsolaterales pequeños y discontinuos; pliegue discoidal y torácico presente; (2) tímpano presente; (3) hocico corto, en vista dorsal subacuminado, angular de perfil; (4) un tubérculo subcónico sobre el párpado superior, rodeado por varios redondeados pequeños; con un tubérculo internarinal subcónico; distancia inter-orbital ligeramente más amplia que el párpado; (5) procesos vomerinos odontóforos presentes, de contorno triangular con 7-8 dientes; (6) hendiduras vocales y almohadillas nupciales desconocido; (7) dedo manual I más corto que el II, discos truncados y con almohadillas dilatadas; (8) dedos manuales con rebordes cutáneos laterales; (9) tubérculos ulnares bajos y pequeños; (10) talón con un tubérculo pequeño cónico; borde externo del tarso con pequeños tubérculos redondeados, pliegue tarsal interno presente; (11) tubérculo metatarsal interno oval, de tres a cuatro veces más grande que el tubérculo externo triangular; tubérculos supernumerarios plantares presentes bajos; (12) dedos pediales con estrechos rebordes cutáneos; membrana interdigital ausente; dedo V más largo que el III; dedo V alcanza al tubérculo subarticular del dedo IV; discos del pie ligeramente más pequeños que los de las manos; (13) color dorsal en preservado gris cenizo, ingles y superficies ocultas negro con manchas blancas, manchas blancas en las superficies ocultas de las extremidades, manchas escapulares negras, barras en los flancos de coloración gris cenizo y mancha rodeando el tubérculo internarinal de color gris oscura (negro en vida); (14) adultos de tamaño mediano, LRC en machos desconocido, hembras 27.1–32.4 mm ( 30 ± 1.7, n = 7).
Comparaciones
Pristimantis churuwiai sp. nov., se distingue de otras especies de Pristimantis de las estribaciones orientales de los Andes y amazonia baja de Ecuador por la presencia de tímpano, tubérculo cónico en el párpado superior, tubérculo subcónico internarinal, distintivas coloraciones crema amarillento en axilas, ingles y superficies de los muslos y esclera azul en vida. La única especie dentro del área de distribución con la que Pristimantis churuwiai sp. nov., podría ser confundido es P. quaquaversus [Citation44]. No obstante, Pristimantis churuwiai presenta un tímpano visible (oculto en P. quaquaversus), tubérculo cónico pequeño en talón (cónico grande), de 7–8 dientes alineados longitudinalmente (4–5 dientes alineados transversalmente), tubérculo subcónico internarinal presente (ausente), esclera de color azul en vida (crema amarillento), manchas distintivas crema amarillento en superficies ocultas (crema rosáceo).
Otras especies con las que P. churuwiai podría ser confundido son P. nigrogriceus de la ladera Oriental de los Andes [Citation45] y P. miktos [Citation46] de las tierras bajas de la Amazonía. Pristimantis churuwiai sp. nov., tiene en la ingle y superficie superior de los muslos una coloración negro con manchas blancas, e iris amarillo metálico en vida (ingle y superficie de los muslos negro con manchas amarillo brillante, e iris rojo en P. nigrogriceus), borde externo del tarso con tubérculos bajos o ausentes (tubérculos cónicos medianos en P. nigrogriceus). Pristimantis churuwiai no presenta crestas dermales en la región escapular (marca en forma de “W” o “V” presente en P. miktos), párpado superior con un tubérculo redondeado pequeño (ausente en P. miktos), talón con tubérculo cónico delgado (sin tubérculo en P. miktos).
Descripción del holotipo
Cabeza más pequeña que el cuerpo, 44% de LRC, longitud de la cabeza ligeramente mayor que la anchura, 107%; hocico corto, subacuminado en vista dorsal y angular de perfil; longitud horizontal de la órbita mayor que la distancia órbita-narina, 136%; narinas dirigidas dorsolateralmente; canto rostral redondeado; región loreal angular; labios ligeramente engrosados; un tubérculo subcónico en párpado superior y varios tubérculos pequeños redondeados; un tubérculo internarinal subcónico; crestas craneales ausentes; fino pliegue supratimpánico que cubre la mitad superior del tímpano; varios pequeños tubérculos postrictales presentes; membrana timpánica presente; anillo timpánico visible dorsalmente, redondeado, su longitud es el 28.8% de la longitud horizontal del ojo; coanas pequeñas y ovaladas, no adheridas al piso palatal; procesos dentígeros del vómer medianos y triangulares de contorno, con 7-8 dientes alineados longitudinalmente; lengua ligeramente más larga que ancha, con el borde posterior bilobulado, el 50% anterior está adherida al piso de la boca.
Piel del dorso finamente granular; pliegues dorsolaterales pequeños, discontinuos que se extienden en la parte anterodorsal del cuerpo; pliegue discoidal y torácico presente; superficie ventral aereolada; cubierta cloacal pequeña; tubérculos cloacales presentes, pequeños y dispersos. Tubérculos escapulares presentes; tubérculos pequeños y esparcidos en la región ulnar y el codo. Superficie palmar pequeña; tubérculo palmar acorazonado y ligeramente elevado (Figura (A)), tubérculo tenar mediano y ovalado, aproximadamente 1/2 del ancho del tubérculo palmar; tubérculos supernumerarios presentes agrandados, ligeramente elevados y dispersos; tubérculos subarticulares grandes redondeados y visibles de perfil; dedos manuales con rebordes cutáneos laterales, sin membranas interdigitales; dedo I más corto que el dedo II; disco del dedo I ligeramente expandido, discos de los dedos II–IV expandidos y redondeados (Figura (A)); todos los dedos manuales con cojinetes ventrales definidos por hendiduras circunmarginales. Extremidades posteriores delgadas, longitud de la tibia 58.7% de la longitud rostro-cloaca; talón con un tubérculo cónico pequeño y delgado; tubérculos pequeños cónicos y dispersos a lo largo del borde externo del tarso; tubérculo metatarsal interno mediano, elevado y alargado, aproximadamente cinco veces más grande que el tubérculo metatarsal externo triangular; tubérculos supernumerarios plantares presentes, ligeramente elevados; tubérculos subarticulares redondeados y elevados; dedos pediales con finos rebordes cutáneos laterales; discos de los dedos pediales de similar tamaño que los discos de los dedos manuales, los discos están definidos por hendiduras circunmarginales; longitud relativa de los dedos del pie: I < II < III < V < IV; dedo III del pie más pequeño que el dedo V; dedo V del pie llega al borde anterior del penúltimo tubérculo subarticular del dedo IV (Figura (B)).
Figura 8. Pristimantis churuwiai sp. nov. (DHMECN 12242). (A) Vista palmar de la mano; (B) vista plantar del pie. Longitud del pie 14 mm.

Figura 9. Pristimantis churuwiai sp. nov., en preservado. (A) Vista dorsal; (B) vista ventral del holotipo DHMECN 12242, hembra adulta, LRC: 29.8 mm.

Figura 10. Pristimantis churuwiai sp. nov., en vida. Vista dorsal y ventral del holotipo DHMECN 12242, hembra adulta, LRC: 29.8 mm.

Figura 11. (A) Pristimantis delius, ♀, LRC 28.4 mm, DHMECN 12225; (B) P. aff. devillei, ♀, LRC 31 mm, DHMECN 12308; (C) P. diadematus, ♀, LRC 27.3 mm, DHMECN 12275; (D) P. gagliardoi, ♀, LRC 26.3 mm, DHMECN 12336; (E) P. galdi, ♂, LRC 17.1 mm: MEPN 14360; (F) P. ganonotus, ♂, LRC 14 mm: MEPN 14000.

Figura 12. (A) Pristimantis aff. gualacenio, ♀, LRC 27.8, DHMECN 11272; (B) P. incomptus, ♀, LRC 26.9, MEPN 14652; (C) P. inusitatus, ♀, DHMECN 12391; (D) P. katoptroides, ♂, LRC 28 mm, DHMECN 12169; (E) P. lacrimosus, ♀, LRC 22.6 mm, DHMECN 12165; (F) P. lanthanites, ♀, LRC 34.7 mm, DHMECN 12385.

Figura 13. (A) Pristimantis latericius ♂, LRC 24.9 mm, DHMECN 12202; (B) P. nigrogroseus, ♀, LRC 20.4 mm, MEPN 14399; (C) P. grp orcesi, ♀, QCAZ 56437; (D) P. aff. pecki, ♂, LRC 17.8 mm: MEPN 14431; (E) P. petersi, ♂, LRC 19.8 mm, DHMECN 12228; (F) P. aff. phoxocephalus, ♂, LRC 26.2 mm, DHMECN 13158.

Figura 14. (A) Pristimantis prolatus, ♂, LRC 17.7 mm, DHMECN 12301; (B) P. aff. prolatus, ♀, LRC 19.5 mm: DHMECN 12372; (C) P. proserpens, ♀, LRC 18.4, DHMECN 12221; (D) P. quaquaversus, ♀, LRC 22.1 mm, DHMECN 12196; (E) P. roni ♂, LRC 27.5 mm, DHMECN 12131; (F) P. rubicundus, ♀, LRC, DHMECN 12192; (G) P. tinguichaca, ♀, LRC 31.1 mm, DHMECN 12293; (H) P. trachiplepharis, ♂, LRC 18.1, DHMECN 12397; (I) P. ventrimarmoratus ♀, LRC 18.8 mm, DHMECN 12386.

Coloración del holotipo en preservado (Figura 9)
Dorso gris cenizo, con barras y manchas gris oscuras hacia la región posterior y zona media del dorso; manchas negras redondeadas dispuestas en la zona media, región escapular, interorbital y nasal. Párpados superiores negruzcos, con tubérculos supraoculares gris claros de aspecto blanquecino (tipo gris luz). Región rostral de la misma coloración del dorso; tímpano monocromático (negro–gris claro). Superficie dorsal de las extremidades anteriores gris claras, ligeramente manchadas a nivel del brazo, gris oscuras y negruzcas en el resto de los miembros anteriores, exceptuando los dedos manuales I–II que son gris claros de aspecto blanquecino; extremidades posteriores del mismo patrón de coloración del dorso, con manchas y barras negruzcas y gris cenizo dispuesto a lo largo de los miembros, dedos pediales con un patrón similar a los dedos manuales (tres primeros dedos gris claros y el resto oscuros), (Figura 9(A)). Vientre vainilla oscuro con tonos grises hacia la parte del pecho y región gular, manchas negras prominentes y pequeñas maculaciones marrones dispuestas a lo largo de la superficie ventral. Superficie ventral de las extremidades anteriores, con un similar patrón de coloración que en su superficie dorsal; extremidades posteriores ligeramente cremas, grisáceas a nivel del muslo, con manchas marrón y gris claras a lo largo de los miembros posteriores (Figura 9(B)). Extremos distales de las superficies palmar y plantar de los dedos más claras en relación a sus superficies dorsales.
Coloración del holotipo en vida (Figura 10)
Dorso marrón claro (tipo marrón desierto), con tonalidades oscuras hacia los flancos, región medial e inguinal, difuminados de manera intercalada a manera de bandas o barras; manchas negruzcas redondeadas dispuestas en la región media del dorso, región escapular, interorbital y nasal. Cabeza de la misma coloración del dorso, canto rostral con banda negruzca que va hasta 1/3 de su extensión. Región rostral de diferentes tonalidades entre marrón oscuro a nivel de las narinas, marrón claro con leves contrastes dorados y verdosos a nivel loreal y vainilla a nivel rostrolateral; manchas marrón oscuras y negruzcas extendidas desde las narinas hasta la región infratimpánica. Tímpano policromático entre tonalidades negras, marrones, cremas y ligeramente grises. Superficie dorsal de las extremidades anteriores y posteriores del mismo patrón de coloraciones del dorso, con barras marrón oscuras, profusas y conspicuas en las extremidades posteriores. Miembros manuales y pediales con el mismo patrón de coloración del dorso y extremidades (marrón desierto-marrón oscuro), exceptuando los extremos distales de los dedos manuales que son de tonos vainillas ligeramente amarillentos. Vientre vainilla oscuro con tonalidades más claras hacia los flancos y mandíbula, manchas marrones amorfas distribuidas a lo largo de la superficie ventral, siendo conspicuas y negras a nivel posterior (superficie ventrolateral). Superficie ventral de las extremidades anteriores de la misma coloración del vientre, extremidades posteriores marrón, ligeramente cobrizo hacia el borde interno de los muslos, con pequeñas maculaciones negras y redondeadas; tercio distal de los muslos y resto de los miembros posteriores de la misma coloración del vientre. Superficie plantar más oscura (marrón oscuro) que la palmar (vainilla claro), en especial el extremo distal de los dedos. Iris amarillo metálico, con ligeros tonos verdosos hacia la región supraocular, indio hacia la infraocular, con reticulaciones cobrizas extendidas hacia el radio, parte media y región infraocular; esclerótica azulada (Figura 10).
Medidas del holotipo (mm). Longitud rostro-cloaca = 29.8; distancia órbita-narina = 3.8; longitud cefálica = 13.2; ancho cefálico = 12.3; distancia interorbital = 3.7; distancia internarinal = 4.5; longitud de la tibia = 17.5; longitud del pie = 14; longitud de la mano = 8.7; diámetro del tímpano = 1.5; longitud horizontal del ojo = 5.2; ancho del párpado = 3.5.
Variación
La variabilidad de las medidas de la serie tipo se presenta en la Tabla . El paratipo DHMECN 12244 presenta una banda interorbital de color crema.
Tabla 2. Medidas morfométricas (en mm) de Pristimantis sp. nov. Mínima, máxima, promedio y desviación estándar en paréntesis.
Distribución y ecología
La especie nueva es conocida de dos localidades; en Sardinayacu dentro del Parque Nacional Sangay y en Yapit en el Bosque Protector Kutukú–Shaimi, provincia de Morona Santiago, Ecuador, en una amplitud altitudinal entre los 1400 a 1800 msnm (Figuras and (A)). El piso zoogeográfico donde habita Pristimantis churuwiai pertenece al Piso Subtropical [Citation37] el ecosistema corresponde al Bosque siempreverde piemontano del sur de la Cordillera Oriental de los Andes [Citation38]. El hábitat se caracteriza por árboles con troncos y ramas nudosas donde albergan gran cantidad de orquídeas y bromelias; la altura de la vegetación emergente alcanza hasta 30 m de altura, dominado por árboles de romerillo (Prumnopitys montana) y palma real (Dictyocaryum lamarckianum); el sotobosque presenta dominancia de herbáceas de las familias Araceae y Marantaceae. Pristimantis churuwiai fue colectado en las primeras horas de la noche (19:00–21:00), al borde de pequeños cuerpos de agua y en el interior del bosque, perchados sobre hojas entre los 0.8 y 1.8 m sobre el suelo. Fue colectado en simpatría con Pristimantis almendariz, P. katoptroides, P. aff. petersi, P. roni y P. rubicundus.
Comentarios taxonómicos
Los caracteres morfológicos utilizados para separar a Pristimantis churuwiai de P. quaquaversus son congruentes para definirlos como linajes evolutivos distintos. No obstante, su alta similitud morfológica nos permite sospechar una fuerte relación filogenética en las dos especies. Los estudios filogenéticos moleculares disponibles, sugieren una estrecha relación de Pristimantis quaquaversus con el grupo de especies de P. frater [Citation47] por lo cual nosotros asignamos a P. churuwiai a éste grupo hasta que futuras evidencias moleculares confirmen o descarten nuestra hipótesis. Mientras tanto, nosotros lo asignamos a este grupo por las siguientes características morfológicas: tamaño corporal pequeño (<32.5 mm), con cuerpo moderadamente robusto, cabeza relativamente ancha, hocico subacuminado y extremidades moderadamente largas; dedo I de la mano más pequeño que el II; dedo V del pie más largo que el III y extendido hasta el margen distal del tubérculo subarticular del dedo IV; dígitos expandidos, cresta craneal ausente, vomerinos presentes.
Pristimantis aff. cremnobates (Figura 7(E))
Una rana similar a Pristimantis cremnobates, caracterizada por su coloración dorsal café pálido con marcas cafés oscuras, vientre café grisáceo, ingles con manchas amarillas, iris blanquecino con reticulaciones ocres; la condición del I dedo manual de similar tamaño que el II lo difiere de P. cremnobates el cual presenta el dedo I más corto que el II [Citation45]. Fue registrado en vegetación arbustiva dentro de bosque maduro entre 1360 y 1800 m en Danú y Sardinayacu. En diciembre del 2010 una hembra grávida fue registrada, contenía 14 ovocitos de color crema. Es necesario una revisión del complejo que resuelva su estatus.
Material de referencia: MEPN 14370, 14538, 14451.
Pristimantis aff. curtipes (Figura 7(F))
Especie similar a Pristimantis curtipes, caracterizada por su coloración dorsal café grisáceo con manchas irregulares más oscuras, vientre blanquecino, iris ocre oscuro; la condición del tímpano visible y los tubérculos plantares supernumerarios escasos lo diferencia de P. curtipes el cual exhibe un tímpano oculto bajo la piel y tubérculos supernumerarios numerosos [Citation48]. Fue registrado en vegetación herbácea de ecotono entre 0 y 0.9 m sobre el suelo dentro del páramo arbustivo a 3500 m en Cisnian.
Material de referencia: DHMECN 12253–57.
Pristimantis delius [Citation49] (Figura 11(A))
Especie que se distribuye en los bosques tropicales y piemontanos de Ecuador y Perú [Citation5,49], en Ecuador habita en escasas localidades de las provincias de Sucumbios, Orellana y Pastaza entre 183 y 1200 [Citation2]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. En la localidad de Domono, P. delius es un habitante del bosque piemontano a 1150 m de altitud. Habita en la vegetación arbustiva del bosque entre 0.8 y 1.7 m sobre el suelo. En Ecuador [Citation6] no ha sido evaluada su amenaza, mientras que a nivel global [Citation8] es considerada una especie con Datos Deficientes.
Material de referencia: DHMECN 12225.
Pristimantis aff. devillei (Figura 11(B))
Especie similar a Pristimantis devillei, caracterizada por presentar un pliegue dorsolateral conspicuo, una coloración dorsal café grisácea, vientre blanquecino e iris rojizo; la condición del tímpano pequeño y los tubérculos plantares supernumerarios numerosos lo diferencia de P. devillei el cual exhibe un tímpano prominente y ausencia de tubérculos plantares supernumerarios [Citation45]. Ésta especie fue registrada en el Cerro Sambalán entre los 3200 y 3500 m, perchado sobre hojas entre 0.3 y 0.6 m sobre el suelo durante la noche.
Material de referencia: DHMECN 12303–25.
Pristimantis diadematus (Jiménez de la Espada, 1875) (Figura 11(C))
Especie ampliamente distribuida en los bosques tropicales y piemontanos de Ecuador, Perú y Brasil [Citation5], en Ecuador habita en todas las provincias de la Amazonía entre 100 a 1200 [Citation2,50]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. En la localidad de Domono, P. diadematus es un habitante del bosque piemontano a 1150 m de altitud. Habita en la vegetación arbustiva del bosque, entre 0.7 y 1.4 m sobre el suelo. En Ecuador [Citation6] y a nivel global [Citation8] lo categorizan como una especie en Preocupación Menor.
Material de referencia: DHMECN 12275.
Pristimantis gagliardoi [Citation51] (Figura 11(D))
Especie endémica de Ecuador, restringida a los bosques montanos de Cañar y Morona Santiago entre 2800 a 3000 [Citation10,51]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. En la localidad de Tinguichaca, P. gagliardoi es un habitante del bosque piemontano entre 2700 a 2900 m de altitud, donde habita en la vegetación baja del bosque entre 0.3 y 1.70 m sobre el suelo; es la especie más abundantes en el área. En Ecuador [Citation52] es considerada una especie En Peligro, mientras que a nivel global [Citation8] no ha sido evaluada.
Material de referencia: DHMECN 12326–45.
Pristimantis galdi (Jiménez de la Espada, 1870) (Figura 11(E))
Especie que se distribuye en las estribaciones orientales de Colombia, Perú y Ecuador [Citation5,43], en Ecuador habita en todas las provincias de la Amazonía desde Sucumbíos hasta Zamora-Chinchipe, entre 1000 a 2900 m de elevación [Citation2,42,45,50]. En la localidad de Sardinayacu y Guabisai, P. galdi es un habitante del bosque piemontano y montano entre 1800 y 2300 m de altitud. Habita la vegetación arbustiva y herbácea, perchando sobre hojas, entre musgos y ocasionalmente en bromelias; generalmente están activos en el día y en el crepúsculo; los machos vocalizan esporádicamente durante el día o en la noche; es una especie común. En diciembre del 2010 tres hembras grávidas fueron registradas, contenían entre 15 y 28 ovocitos de color amarillentos. En Ecuador [Citation6] lo categorizan como una especie en Preocupación Menor, mientras que a nivel global [Citation8] se lo cita como Casi Amenazada.
Material de referencia: DHMECN 12177–12190; MEPN 14360, 14365, 14367, 14369.
Pristimantis ganonotus [Citation53] (Figura 11(F))
Especie endémica de Ecuador, que se distribuye en los bosques piemontanos y montanos de las provincias de Napo, Pastaza y Morona Santiago, entre 1700 a 2000 m de elevación [Citation42,53]. En la localidad de Sardinayacu y Guabisai, P. ganonotus es un habitante del bosque piemontano entre 1800 y 1950 m de altitud. Habita la vegetación herbácea, perchando generalmente sobre hojas de las familias Araceae, Piperaceae y Marantaceae; suelen estar activos durante el día y en el crepúsculo; los machos presentan un amplio repertorio vocal y cantan durante las primeras horas del día [Citation54], es una especie poco común. En Ecuador [Citation6] la amenaza no ha sido evaluada y a nivel global [Citation8] es considerada una especie con Datos Deficientes.
Material de referencia: DHMECN 12270, 12167–8; MEPN 14000, 14401-2.
Pristimantis aff. gualacenio (Figura 12(A))
Corresponde a una especie similar a Pristimantis gualacenio [Citation55], caracterizada por su coloración dorsal café pálido, tubérculos cónicos en el párpado superior e iris blanquecino con una banda ancha longitudinal de color ocre; la presencia de pliegues dorsolaterales y superficies ocultas de las extremidades e ingles con tonos rojizos difieren de P. gualacenio el cual carece de pliegues dorsolaterales y presenta conspicuas manchas amarilla y negras en las ingles. [Citation55] Fue registrado en vegetación herbácea dentro y al borde de bosque entre 2800 y 3500 m en Tinguichaca, Sambalán y Cisnian.
Material de referencia: DHMECN 11927–51, 12258–59.
Pristimantis incomptus (Lynch & Duellman, 1980) (Figura 12(B))
Especie que se distribuye en las estribaciones orientales de los Andes de Ecuador y norte de Perú entre 1400 a 1910 m de elevación [Citation2,5,42,45]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. En la localidad de río Sardinayacu habita en el interior del bosque prístino entre 1350 y 1400 m de altitud, percha en la vegetación arbustiva y sobre bromelias, es una especie rara. En Ecuador [Citation6] es categorizada como una especie Casi Amenazada y a nivel global [Citation8] como una especie Vulnerable.
Material de referencia: MEPN 14652–3.
Pristimantis inusitatus (Lynch & Duellman, 1980) (Figura 12(C))
Especie endémica de la vertiente oriental de los Andes, habita en las provincias de Napo y Pastaza, entre 2000 y 2200 m. [Citation2,41,45] Padial et al. [Citation24] no lo asignan a ningún grupo de especies. Yánez-Muñoz et al. [Citation15] lo incluyen en el ecotipo de ranas verde-espinosas. Aquí citamos por primera vez la especie para la provincia de Morona Santiago, a 108 km de Abitagua localidad reportada como las más austral [Citation45]. En la localidad de Guabisai, P. inusitatus es un habitante del bosque montano prístino a 2400 m de altitud. Habita la vegetación arbustiva entre 2 m de altura, perchando sobre hojas o ramas pequeñas. En Ecuador [Citation6,52] se lo categorizan como una especie En Peligro y a nivel global [Citation8] se lo cita como una especie Vulnerable.
Material de referencia: DHMECN 12391–3.
Pristimantis katoptroides [Citation56] (Figura 12(D))
Especie que se distribuye en la vertiente oriental de los Andes de Ecuador y Perú [Citation56–59], en Ecuador habita desde la provincia de Sucumbios hasta Zamora-Chinchipe, entre 1000 y 1800 m [Citation42,56,59]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. Yánez-Muñoz et al. [Citation15] lo incluyen en el ecotipo de ranas verde-espinosas. En la localidad Danu y río Sardinayacu, P. katoptroides es un habitante del bosque piemontano prístino, entre 1360 y 1400 m de altitud. Habita la vegetación arbustiva entre 2 y 6 m de altura, perchando sobre hojas o ramas pequeñas, a menudo también suele ser observado sobre el musgo de los troncos de los árboles donde se mimetiza a la perfección; en septiembre de 2014 numerosos machos vocalizaban durante las primeras horas de la noche. En Ecuador [Citation6] y a nivel global [Citation8] lo categorizan como una especie En Peligro.
Material de referencia: DHMECN 12169–76.
Pristimantis lacrimosus (Jiménez de la Espada, 1875) (Figura 12(E))
Especie que se distribuye en las tierras bajas de Colombia, Ecuador, Perú y Brasil [Citation1], en Ecuador habita en las provincias de Sucumbíos hasta Morona Santiago [Citation2,45,60]. Padial et al. [Citation24] lo asignan al grupo de especies de Pristimantis lacrimosus. En la localidad de Domono, P. lacrimosus es un habitante de borde de bosque y en áreas de cultivo a 1150 m de altitud, prefiriendo el estrato bajo en áreas abiertas entre los 0.5 y 0.9 m sobre el suelo; la mayor actividad vocal de los machos suele darse durante las primeras horas de la noche. En Ecuador [Citation6] y a nivel global [Citation8] lo categorizan como una especie con Preocupación Menor.
Material de referencia: DHMECN 12165–6.
Pristimantis lanthanites (Lynch, 1975) (Figura 12(F))
Especie que se distribuye en la vertiente oriental de los Andes de Colombia, Ecuador, Perú y Brasil [Citation2,43], en Ecuador habita en todas las provincias de la Amazonía, entre 200 y 1650 m [Citation2]. Padial et al. [Citation24] lo asignan al grupo de especies Pristimantis ridens. En la localidad Domono, P. lanthanites es un habitante del bosque piemontano prístino y áreas alteradas como cultivos y bordes de bosque, a 1150 m de altitud. Habita la vegetación herbácea entre 0.3 y 0.8 m de altura, perchando sobre hojas o ramas pequeñas. En Ecuador Ron et al. [Citation6] y a nivel global la UICN [Citation8] lo categorizan como una especie en Preocupación Menor.
Material de referencia: DHMECN 12385.
Pristimantis latericius [Citation14] (Figura 13(A))
Especie endémica de Ecuador, recientemente descrita de los bosques montanos del Parque Nacional Sangay a 2750 m de altitud [Citation14]. Batallas and Brito [Citation14] y Padial et al. [Citation24] lo asignan al grupo de especies de Pristimantis lacrimosus. En la localidad de Tinguichaca y Guabisai, P. latericius es un habitante del bosque montano entre los 2300 y 2700 m de altitud, prefiriendo el estrato bajo del bosque o áreas abiertas entre los 0.5 y 1.3 m sobre el suelo; la mayor actividad vocal de los machos suele darse durante las noches con lluvia ligera. En Ecuador [Citation6] se lo considera como una especie con Datos Insuficientes para evaluar su amenaza.
Material de referencia: DHMECN 12200–10.
Pristimantis nigrogriseus (Andersson, 1945) (Figura 13(B))
Especie endémica de Ecuador, restringida a la ladera oriental de los Andes entre 1000 a 2200 m de altitud, entre las provincias de Sucumbíos hasta Zamora Chinchipe [Citation2,45,59]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. En la localidad de Sardinayacu, P. nigrogriseus es un habitante del bosque montano a 1800 m. Prefiriendo el estrato arbustivo del bosque prístino entre 1.5 y 2 m sobre el suelo; en el área de estudio es una especie rara. En Ecuador [Citation6] lo categorizan como una especie Casi Amenazada y a nivel global [Citation8] como una especie Vulnerable.
Material de referencia: MEPN 14393–4, 14396–9.
Pristimantis grp. orcesi (Figura 13(C))
Es una distintiva especie de Pristimantis, caracterizada por su llamativa coloración dorsal verde con manchas ocres, superficie ventral y partes ocultas de las extremidades verdosas e iris turquesa; por sus características como cuerpo robusto, cabeza estrecha, extremidades moderadamente cortas y dedo V del pie mucho más largo que el III lo relacionamos con el grupo P. orcesi [Citation22], Fue registrado en vegetación arbustiva entre 1.5 y 2.5 m sobre el suelo, dentro de bosque maduro a 2200 m en las cercanías del volcán Sangay.
Material de referencia: QCAZ 56437–39.
Pristimantis aff. pecki (Figura 13(D))
Una rana terrestre asociada a Pristimantis pecki [Citation53], se caracteriza por presentar una coloración dorsal ocre con una mancha crema en la región cefálica, un pliegue dorsolateral extendido desde la escápula hasta el sacro, la presencia del tímpano pequeño y el I dedo manual ligeramente más pequeño que el II lo diferencia de P. pecki, el cual presenta un tímpano prominente y el I dedo manual mucho más pequeño que el II [Citation53]. Fue registrado en Sardinayacu a 1800 m, perchando sobre una hoja a 1.5 m sobre el suelo.
Material de referencia: MEPN 14392.
Pristimantis petersi [Citation45] (Figura 13(E))
Especie endémica de Ecuador; se distribuye en la ladera oriental de los Andes de 1400 a 1800 m de altitud, entre las provincias de Sucumbíos hasta Zamora Chinchipe [Citation2,45]. Padial et al. [Citation24] lo asignan al grupo de especies Pristimantis lacrimosus. En la localidad de Sardinayacu, es un habitante del bosque piemontano entre 1400 y 1800 m de altitud, prefiriendo el estrato arbustivo del bosque prístino entre 1.5 y 3 m sobre el suelo; especie de actividad nocturna; sus vocalizaciones son parecidas a un «tic» metálico que emite desde arbustos y árboles; sus coros carecen de orden específico, intensificando su actividad en lluvias ligeras, generando silencios prolongados en particular cuando canta otra especie [Citation61], en el área de estudio es una especie común. En Ecuador [Citation6,52] lo consideran una especie Vulnerable, mientras que a nivel global la especie no ha sido evaluada.
Material de referencia: DHMECN 12227-30.
Pristimantis aff. phoxocephalus (Figura 13(F))
Especie similar a Pristimantis phoxocephalus [Citation3], caracterizada por presentar un amplio saco subgular medio, una coloración dorsal café amarillento, vientre blanquecino, flancos amarillos con manchas negruzcas e iris cobrizo; la presencia de tubérculos en el talón y tarso, más una ingle rosácea con tonos amarillentos difieren de P. phoxocephalus, el cual carece de tubérculos en el talón y tarso y presenta una coloración blanca con reticulaciones marrones o negras en la ingle [Citation3]. Ésta especie fue registrada en Cisnian a 3300 m, perchado sobre hojas entre 0.3 y 0.7 m sobre el suelo.
Material de referencia: DHMECN 12308.
Pristimantis prolatus [Citation44] (Figura 14(A))
Especie endémica de la vertiente oriental de los Andes de Ecuador desde la provincia de Napo hasta Morona-Santiago, entre 1140 y 1700 m [Citation2,42,45]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. En la localidad río Sardinayacu se registró en el interior del bosque prístino a 1400 m de altitud; habita en la vegetación herbácea entre 0.6 y 1.2 m de altura. En Ecuador [Citation6,52] y a nivel global [Citation8] es considerada como una especie En Peligro.
Material de referencia: DHMECN 12301.
Pristimantis aff. prolatus (Figura 14(B))
Especie similar a Pristimantis prolatus [Citation45], caracterizada por presentar un pliegue dérmico en forma de W en la escápula, una coloración dorsal de café ocre a café oscuro, vientre blanquecino e iris amarillento; el pliegue dérmico en forma de W y varios tubérculos cónicos sobre el párpado lo diferencias de P. prolatus, mismo que presenta un pliegue dérmico en forma de H y un tubérculo punzante sobre el párpado [Citation45]. Ésta especie fue registrada en Danu y río Sardinayacu entre 1360 y 1450 m, perchado sobre hojas entre 0.1 y 0.3 m sobre el suelo.
Material de referencia: DHMECN 12372–79.
Pristimantis proserpens [Citation62] (Figura 14(C))
Es una rana endémica de las estribaciones orientales de Ecuador en las provincias de Morona Santiago y Zamora Chinchipe, entre 1710 y 2600 m [Citation45,50,62]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. En Guabisai es una especie que habita a 2600 m, en el interior de bromelias terrestres y arbóreas entre 0 y 2.5 m sobre el suelo; suele ser una especie poco común, prefiriendo áreas con abundante entrada de luz como las cumbres de montaña. El 28 marzo del 2015 una hembra con una puesta de 11 huevos, fue registrada en el interior de una bromelia terrestre (Tillansia sp). En Ecuador [Citation6,52] y a nivel global [Citation8] es considerada como una especie En Peligro.
Material de referencia: DHMECN 12218–24.
Pristimantis quaquaversus [Citation44] (Figura 14(D))
Especie ampliamente distribuida en la Amazonía desde las estribaciones bajas de los Andes en el norte de Ecuador, hacia el lado adyacente del norte de Perú entre 200 y 1750 m de elevación [Citation42,43]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. Ha sido registrada en la localidad de Domono a 1150 m, en el interior de bosque en regeneración y dentro del bosque prístino; percha en la vegetación herbácea entre los 0.5 y 0.8 m de altura. En Ecuador [Citation6] y a nivel global [Citation8] lo categorizan como una especie en Preocupación Menor.
Material de referencia: DHMECN 12196–99; MEPN 14493–4.
Pristimantis roni [Citation15] (Figura 14(E))
Especie endémica de Ecuador, recientemente descrita de los bosques piemontanos del Parque Nacional Sangay, a 1900 m de altitud [Citation15]. Yánez-Muñoz et al. [Citation15] no lo asignan a ningún grupo de especies, sin embargo, proponen que podría formar parte del grupo de especies Pristimantis devillei por la presencia de cresta craneal; mientras tanto lo incluyen en el ecotipo de ranas verde-espinosas. En la localidad de Sardinayacu, P. roni es un habitante del bosque piemontano entre 1750 y 1900 m de altitud. Prefiriendo el estrato bajo y arbustivo del bosque entre los 0.5 y 1.5 m sobre el suelo. El estado de conservación no ha sido evaluado.
Material de referencia: DHMECN 11311–27.
Pristimantis rubicundus (Jiménez de la Espada, 1875) (Figura 14(F))
Es una rana terrestre endémica de las estribaciones orientales de Ecuador desde la provincia de Napo hasta Morona Santiago, entre 1080 y 1560 m [Citation10,42,45]. Hedges et al. [Citation22] asignaron a esta especie dentro del subgénero Hypodiction y Padial et al. [Citation24] lo asignaron dentro del grupo Pristimantis ridens. En Danu y en el río Sardinayacu la especie se encuentra entre 1360 a 1400 m, es una especie que habita en la vegetación herbácea entre 0.3 y 0.7 m sobre el suelo; suelen ser abundantes al borde de pequeñas quebradas. En Ecuador [Citation6,52] y a nivel global [Citation8] lo categorizan como una especie En Peligro de extinción.
Material de referencia: DHMECN 12191–95, MEPN 14498.
Pristimantis sambalan sp nov
Nombre propuesto en español: Cutín de Sambalán
Proposed English names: Sambalan Rainfrog
Holotipo (Figuras 15–17)
Hembra adulta DHMECN 12251, colectada por JBM y Glenda Pozo el 18 de enero del 2015 en el Cerro Sambalán (2º12′22.1″S; 78º27′09.7″O, 2851 m), Parque Nacional Sangay, parroquia Zuñac, cantón Morona, provincia de Morona Santiago, República del Ecuador.
Paratopotipos
Hembras adultas DHMECN 12249, 12250 colectados por JBM y Glenda Pozo el 18 de enero del 2015, MEPN 14676 colectada por JBM y Segundo Tenemaza el 28 de noviembre del 2011; machos adultos DHMECN 12252 colectado por JBM y Glenda Pozo el 18 de enero del 2015, MEPN 14677 colectado por JBM y Segundo Tenemaza el 28 de noviembre del 2011. Colectados en la misma área del holotipo.
Etimología
El epíteto hace referencia al Cerro Sambalán, la localidad tipo, un sector del Parque Nacional Sangay.
Diagnóstico
Una especie de Pristimantis del grupo de especies P. myersi caracterizada por la combinación de los siguientes caracteres: (1) Piel del dorso tuberculado; vientre areolado; flancos con pústulas prominentes dispersas; fino pliegue dorsolateral formado por una hilera de pequeños tubérculos subcónicos elevados; pliegue discoidal presente; (2) tímpano presente; (3) hocico corto, en vista dorsal subacuminado, redondeado de perfil; (4) de uno a dos tubérculos cónicos y varios pequeños redondeados sobre el párpado superior; distancia inter-orbital ligeramente más amplia que el párpado; (5) procesos vomerinos con odontóforos pequeños y oblicuos de contorno; (6) macho sin hendiduras vocales y almohadillas nupciales; (7) dedo manual I más corto que el II, discos truncados y con almohadillas dilatadas; (8) dedos manuales con estrechos rebordes cutáneos; (9) tubérculos ulnares bajos y pequeños; (10) talón y borde externo del tarso con tubérculos cónicos pequeños; pliegue tarsal interno presente; (11) tubérculo metatarsal interno ovoide, tres veces más grande que el externo subcónico; (12) dedos pediales con estrechos rebordes cutáneos; membrana interdigital ausente; dedo V más largo que el III, no alcanza al tubérculo subarticular del dedo IV; discos del pie ligeramente más pequeños que los de las manos, tubérculos supernumerarios plantares pequeños; (13) dorsalmente marrón castor (en vida kobe siena), vientre marrón grisáceo finamente punteado de café oscuro (marrón beige con granulaciones de color café oscuro); ingle y superficies ocultas del muslo marrón grisáceo (rojo herrumbe); iris marrón claro con reticulaciones ocre y con línea media horizontal café rojiza; (14) adultos de tamaño pequeño, LRC en el macho 15.6 mm (n = 2), hembras 20.6–21.9 mm ( 21.3 ± 0.7, n = 4).
Comparaciones
Pristimantis sambalan sp. nov., es un miembro del grupo P. myersi (sensu Guayasamin et al. [Citation35], de las estribaciones orientales de Ecuador que inmediatamente se diferencia de sus congéneres por su distintiva textura de la piel fuertemente tuberculada y con pústulas muy bien definidas en los flancos.
La única especie de Pristimantis del Parque Nacional Sangay [Citation10] con la que podría ser confundido es P. bicantus [Citation41] sin embargo, la presencia de pústulas en los flancos, discos expandidos y la presencia de un tubérculo cónico en el párpado en P. sambalan lo diferencia con facilidad de P. bicantus (pústulas ausentes, discos reducidos y tubérculo subcónico en el párpado).
Pristimantis sambalan se distingue de otros congéneres del grupo P. myersi de la vertiente oriental de los Andes: P. festae, P. gladiator y P. marcoreyesi con superficies de las ingles con coloraciones llamativas, sin embargo éstas tres especies carecen de pústulas llamativas en los flancos.
Finalmente, Pristimantis sambalan que podrían ser confundido con siete especies de las estribaciones occidentales de los Andes de Ecuador, entre ellas: P. floridus, P. hectus, P. leoni, P. lucidosignatus, P. munozi y P. pyrromerus. Pristimantis floridus [Citation3] discos expandidos y superficies ocultas de las piernas de color rojo; sin embargo, P. sambalan se distingue por la presencia de pústulas en los flancos, finos pliegues dorsolaterales, dígitos con rebordes cutáneos y tubérculos supernumerarios presentes (ausentes en P. floridus). Pristimantis hectus [Citation3], presenta un peculiar tubérculo metatarsal en el borde interno del tarso y una coloración en las superficies ocultas de los muslos con puntos blancos (sin tubérculo peculiar y superficies ocultas de color rojizo en P. sambalan). Pristimantis leoni [Citation3], tiene los testículos de color negro y los discos digitales angostos (testículos blancos y discos digitales amplios en P. sambalan). Pristimantis lucidosignatus [Citation63], presenta una cresta escapular en forma de “W”, pliegue supratimpánico ausente (cresta escapular ausente y pliegue supratimpánico presente en P. sambalan). Pristimantis munozi [Citation64], presenta una coloración gris en la ingle con marcas blancas y vientre naranja (ingle rojizo y vientre translúcido en P. sambalan). Pristimantis pyrromerus [Citation3], tiene un pliegue occipital en forma de “W”, discos digitales angostos e iris verde crema (sin pliegue occipital, discos digitales amplios e iris cobrizo en P. sambalan).
Descripción del holotipo
Cabeza más pequeña que el cuerpo, 44% de LRC, cabeza ligeramente más larga que ancha, 120% del ancho; hocico corto, subacuminado en vista dorsal y redondeado de perfil; longitud horizontal del ojo mayor que la distancia ojo-narina, 152%; región interorbital ligeramente mayor que el ancho del párpado, 77.7%; narinas dirigidas lateralmente; canto rostral redondeado; región loreal ligeramente angular; labios ligeramente engrosados; dos tubérculos cónicos y varios pequeños redondeados sobre el párpado superior; crestas craneales ausentes; pliegue supratimpánico cubriendo el borde superior del tímpano; dos tubérculos postrictales presentes y agrandados; membrana timpánica presente; anillo timpánico visible redondeado, su longitud es el 42.8% de la longitud horizontal del ojo; coanas pequeñas redondeadas y no adheridas al piso palatal; procesos vomerianos medianos, oblícuos y con bordes irregulares, dientes diferenciables (3–4), redondeados y agrupados medialmente; lengua ligeramente más larga que ancha, con el borde posterior bilobulado, con el 50% anterior adherida al piso de la boca. Piel del dorso tuberculado; finos pliegues dorsolaterales presentes, formados por hileras de tubérculos subcónicos; fino pliegue dérmico escapular en forma de “W” formado por hileras de pequeños tubérculos subcónicos; flancos con pústulas dispersas; superficie ventral incluyendo la garganta areolada; pliegue discoidal presente; cubierta cloacal pequeña, tubérculos cloacales presentes y agrandados. Tubérculos escapulares presentes; dos tubérculos sacrales subcónicos; tubérculos pequeños y esparcidos en la región ulnar y el codo. Superficie palmar pequeña; tubérculo palmar redondeado y elevado (Figura (A)), tubérculo tenar grande y alargado, aproximadamente 1/2 del ancho del tubérculo palmar; tubérculos supernumerarios grandes, ligeramente elevados y dispersos; tubérculos subarticulares grandes redondeados; dedos manuales con finos rebordes cutáneos, sin membranas interdigitales; dedo I más corto que el dedo II; disco del dedo I ligeramente expandido, discos de los dedos II–IV expandidos y redondeados (Figura (A)); todos los dedos manuales con cojinetes ventrales definidos por hendiduras circunmarginales. Extremidades posteriores delgadas, longitud de la tibia 61% de la longitud rostro–cloaca; talón con dos tubérculos cónicos pequeños; tubérculos pequeños cónicos y dispersos a lo largo del borde externo del tarso; tubérculo metatarsal interno grande, elevado y alargado, aproximadamente dos veces más grande que el tubérculo metatarsal externo; tubérculos supernumerarios plantares pequeños y dispersos; tubérculos subarticulares redondeados; dedos pediales con débiles rebordes cutáneos; discos de los dedos del pie ligeramente más pequeños que los discos de los dedos manuales, los discos están definidos por hendiduras circunmarginales; longitud relativa de los dedos del pie: I < II < III < V < IV; dedo III del pie más pequeño que el dedo V, dedo V del pie no alcanza al tubérculo subarticular distal del dedo IV (Figura (B)).
Figura 15. Pristimantis sambalan sp. nov. (DHMECN 12251). (A) Vista palmar de la mano; (B) vista plantar del pie. Longitud del pie 11.6 mm.
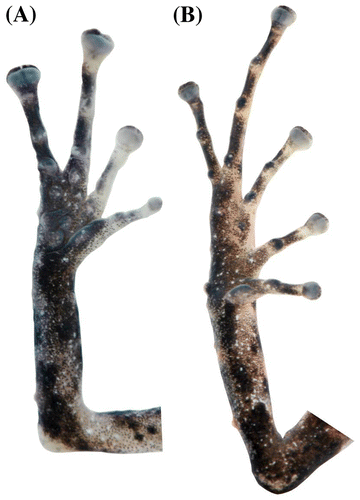
Coloración del holotipo en preservado
Dorso marrón castor, con tonos ocres y marrón claro hacia los flancos, gris pardo hacia la región cefálica; tubérculos marrón oscuros en la región dorsolateral, y manchas marrón entre oscuras y claras (deslucidas) en la región posterior, línea media, región supraescapular (forma de “W”), hasta la región interorbital, donde es más oscura (Figura (A)). Párpado superior marrón pálido, con tubérculo supraocular amarillento. Región rostral más clara que el dorso, con bandas marrones descoloridas. Superficie ventral marrón grisácea (beaver) con tonalidades más claras o cremas hacia el abdomen, y manchas redondas marrones, más notorias hacia la garganta, región gular y maxilar (Figura (B)). Extremidades anteriores marrón pálido, a nivel dorsal y ventral; posteriores de color salmón en los muslos, teniendo un color cobre pálido a nivel ventral; las piernas a nivel dorsal son de tonalidades marrones, mientras que a nivel ventral mantienen el patrón de coloración de los muslos; las bandas que cruzan la superficie dorsal de las extremidades son más opacas y poco notorias en las posteriores. Miembros manuales y pediales maculados, exceptuando el borde externo de la superficie dorsal de los miembros pediales, donde son de colores cobre pálidos. Triángulo cloacal café finamente delineado de blanco (Figura ).
Figura 16. Pristimantis sambalan sp. nov., en preservado. (A) Vista dorsal; (B) vista ventral del holotipo DHMECN 12251, hembra adulta, LRC: 21.5 mm.

Coloración en vida
Dorso kobe siena con tubérculos marrón oscuros que se extienden hasta la región posterior (Figura ), siendo más conspicuos en la región dorsolateral; manchas del mismo color que los tubérculos, dispuestos en la región posterior, línea media, región supraescapular (forma de “W”), hasta la región interorbital. Párpados superiores ligeramente cafés, con tubérculos supraoculares de la misma coloración del dorso. Área nasal marrón ocre, formando una especie de máscara o antifaz. Región rostral más clara que el dorso, con bandas marrón oscuras que se extienden desde el canto, siguiendo a lo largo de la superficie loreal, región infraocular; hasta el anillo timpánico. Vientre marrón beige, con tonalidades pálidas acromáticas tipo taupe hacia los flancos y rojizos a nivel posterior (Figura ), con dispersas manchas redondas marrones a nivel del abdomen y pecho. Garganta y región gular grisáceas, con manchas irregulares marrón oscura, más prominentes a nivel maxilar. Superficie dorsal de las extremidades anteriores y posteriores marrón toscana (a excepción del brazo que es de color marrón ocre), con bandas más oscuras que se extienden a lo largo de las mismas, hasta el extremo distal de las superficies dorsales de los miembros manuales y pediales. Superficie ventral de las extremidades anteriores con el mismo patrón de coloración que el vientre, con tonos marrones hacia los bordes; superficie ventral de las extremidades posteriores rojo herrumbre (rust) a nivel de los muslos, marrón claro en las piernas y tarsos, con manchas de forma irregular (menos notorias hacia los muslos). Iris marrón claro (marrón de madera), con tonalidades cobrizas a nivel preocular y reticulaciones ocre a nivel supraocular.
Figura 17. Pristimantis sambalan sp. nov., en vivo. Vista dorsal y ventral del holotipo DHMECN 12251, hembra adulta, LRC: 21.5 mm.

Medidas del holotipo (mm). Longitud rostro-cloaca = 21.5; ancho cefálico = 7.9; longitud cefálica = 9.5; longitud de la tibia = 13.2; longitud del pie = 11.6; longitud horizontal del ojo = 3.5; distancia órbita-narina = 2.3; distancia internarinal = 4; distancia interorbital = 2.7; diámetro del tímpano = 1.5; longitud de la mano = 7.3; ancho del párpado = 2.1.
Variación
La serie de medidas morfométricas de la serie tipo se presenta en la Tabla . Variaciones cromáticas observadas en la serie tipo incluyen: holotipo DHMECN 12251 con finos pliegues dorsolaterales pigmentados de crema, especímenes con DHMECN 12250 con banda interorbital crema, y DHMECN 12249 con mancha cefálica triangular de color crema. El ejemplar macho 12252 presentó una distintiva marca escapular café oscuro en forma de “W” y de “V” invertida en el sacro; éste espécimen presenta el vientre, pecho y garganta densamente pigmentado de café oscuro; a diferencia de las hembras donde la coloración ventral es predominantemente crema.
Distribución y ecología
La especie nueva es conocida únicamente de la localidad tipo ˝Cerro Sambalán˝, en el Parque Nacional Sangay, provincia de Morona Santiago, Ecuador, a una elevación de 2851 msnm. El piso zoogeográfico donde habita Pristimantis sambalan pertenece al Piso Templado [Citation37], el ecosistema corresponde al Bosque siempreverde montano del sur de la Cordillera Oriental de los Andes [Citation38], el cual se caracteriza por árboles con ramas nudosas, abundancia de zurales (Chusquea sp) y el sotobosque dominado por herbáceas de las familias Melastomataceae, Piperaceae Lamiaceae y Asteraceae. Pristimantis sambalan fue colectado en las primeras horas de la noche (19:00–21:00), al borde de una pequeña quebrada de 1.5 m de ancho, perchados sobre hojas entre 0.4 y 0.8 m sobre el suelo. El 28 de noviembre del 2011 una hembra (MEPN 14676) fue registrada con seis ovocitos de color crema. Fue colectado en simpatría con P. gagliardoi e Hyloscirtus psarolaimus.
Comentarios taxonómicos
Nosotros asignamos a Pristimantis sambalan al grupo de especies de P. myersi (sensu [Citation22,24,35] por presentar los siguientes caracteres: tamaño corporal pequeño (<21.9 mm), con cuerpo robusto, hocico corto, extremidades moderadamente largas, procesos vomerianos medianos y oblícuos; dedo I de la mano más pequeño que el II; dedo V del pie más largo que el III; dedo V del pie no alcanza al tubérculo subarticular distal del dedo IV; dedos angostos y largos con discos relativamente anchos. No obstante; advertimos que el grupo está conformado por un complejo de especies crípticas que dificultan establecer sus relaciones filogenéticas [Citation65]. Por lo cual sugerimos que en futuros estudios filogenéticos incorporen secuencias de ADN y análisis de vocalizaciones de este grupo de especies para poner a prueba nuestra hipótesis.
Pristimantis saturninoi sp nov
Nombre propuesto en español: Cutín del Padre Saturnino
Proposed English names: Priest Saturnino Rainfrog
Holotipo (Figuras 18, 20, 21(A))
Hembra adulta DHMECN 12232, colectada por JBM, Glenda Pozo y Reed Ojala-Barbour el 15 de enero del 2015 en el Cerro Sambalan (2º11′56.9″S; 78º26′50.2″O, 3337 m), Parque Nacional Sangay, parroquia Zuñac, cantón Morona, provincia de Morona Santiago, República del Ecuador.
Paratopotipos
Hembras adultas DHMECN 12235, 12240; machos adultos DHMECN 12231, 12233, 12234, 12237, 12238, 12239, 12247 colectados por JBM, Glenda Pozo y Reed Ojala-Barbour, el 18 de enero del 2015. Colectados junto con el holotipo.
Etimología
El nombre especifico saturninoi hace referencia al Padre Saturnino López, sacerdote Salesiano que por más de dos décadas ha gestionado y trabajado arduamente en numerosos proyectos de desarrollo social y productivo en Morona Santiago, principalmente en las comunidades de la parroquia Zuñac.
Diagnóstico
La nueva especie es asignada al género Pristimantis [Citation3,22,24]. Se diferencia de otras especies congenéricas por la combinación de los siguientes caracteres: (1) Piel del dorso tuberculado; vientre areolado, presencia de pliegue dorso lateral; pliegue discoidal presente; (2) membrana timpánica y anillo timpánico presentes; (3) hocico corto, en vista dorsal redondeado, ligeramente angular de perfil; (4) escasos tubérculos pequeños aplanados y redondeados sobre el párpado superior; distancia inter-orbital más amplia que el párpado; (5) procesos vomerinos con odontóforos diferenciables; (6) machos con hendiduras vocales y almohadillas nupciales; (7) dedo manual I ligeramente más corto que el II, discos redondeados y con almohadillas ligeramente más dilatadas en los dedos III–IV; (8) dedos manuales con estrechos rebordes cutáneos; (9) tubérculos ulnares bajos y pequeños; (10) talón con tubérculos pequeños y aplanados; borde externo del tarso con diminutos tubérculos redondeados, pliegue tarsal interno presente; (11) tubérculos palmares y plantares grandes y aplanados; (12) dedos pediales con estrechos rebordes cutáneos; membrana interdigital basal presente únicamente entre los dedos IV–V; dedo V ligeramente más largo que el III; discos del pie de tamaño similar a los de las manos; (13) en preservado, dorso negro opaco, vientre negro con granulaciones de color marrón a blanco a grisáceas; ingle y superficies ocultas del muslo y pantorrillas con manchas irregulares de color marrón a blanquecino; iris cobrizo en vida; (14) adultos de tamaño pequeño, LRC en machos 14.6–18.7 mm ( 16.8 ± 1.5, n = 7), hembras 19.5–21.5 mm (
20.2 ± 1.2, n = 3).
Comparaciones
La nueva especie se distingue de sus congéneres más similares del grupo Pristimantis Orestes [Citation22,66] en los siguientes caracteres de (caracteres de la especie nueva en paréntesis): P. orestes [Citation67] presenta coloración negra de la ingle y superficies ocultas de las piernas con grandes puntos blancos (ingle negra con manchas agrandadas e irregulares de color blanquecino a crema amarillento); de P. andinognomus [Citation68] el cual presenta la superficie anterior de los muslos y superficie oculta de las pantorrillas café claro con puntos diminutos de color café (superficie anterior de los muslos y pantorrillas de color negro y con manchas agrandadas e irregulares de color blanquecino a crema amarillento), y de Pristimantis bambu [Citation69] que presenta amplias marcas ámbar delineadas de negro sobre un fondo rojizo anaranjado en la ingle y superficie oculta de los muslos (ingle y superficie anterior de los muslos y pantorrillas de color negro y con manchas agrandadas e irregulares de color blanquecino a crema amarillento en P. saturninoi).
Pristimantis mazar [Citation70] presenta el vientre reticulado y líneas negras notorias en los flancos (vientre sin reticulaciones y flancos con bandas difusas en algunos individuos de P. saturninoi). Pristimantis simonbolivari [Citation71] tiene la superficie dorsal lisa y dígitos sin rebordes laterales (dorso tuberculado y rebordes laterales presentes en P. saturninoi).
Otras especies con las que podría confundirse incluye a Pristimantis proserpens, P. colodactylus y P. vidua; sin embargo P. proserpens [Citation62,67], exhibe una coloración dorsal y ventral café y la ingle sin patrón de puntos o manchas, (generalmente negro a café y con patrón de manchas en la ingle de P. saturninoi); P. colodactylus y P. vidua [Citation5,43,45,67] no presentan membrana timpánica (presente en P. saturninoi).
Pristimantis saturninoi difiere en sus caracteres craneales de sus especies más similares, P. orestes y P. vidua [Citation67] por: borde basal del esfenetmoide regular y en forma de media luna (recto e irregular en P. orestes e irregular en P. vidua). Vómer formados por una placa palatal delgada (placa palatal ancha en P. orestes y P. vidua). Las tres especies comparten similaridad en la forma del paresfenoide que presenta una forma de cruz invertida; sin embargo en P. saturninoi la región anterior del processus cultriforme es corto, ancho y con borde irregular (largo, delgado y con borde irregular en P. orestes y largo, delgado y con borde regular en P. vidua).
Descripción del holotipo
Cabeza más pequeña que el cuerpo, 40% de LRC, longitud de la cabeza ligeramente menor que la anchura, 98.8%; hocico corto, redondeado en vista dorsal, ligeramente angular de perfil; longitud horizontal del ojo mayor que la distancia ojo-narina, 160%; narinas dirigidas lateralmente; canto rostral recto; región loreal ligeramente angular; labios delgados; cabeza y el párpado superior con escasos tubérculos pequeños y aplanados; crestas craneales ausentes, parietales con surco medio, fontanelas occipitales profundas y evidentes; pliegue supratimpánico extendido desde la región anterior del ojo hasta el borde del primer tubérculo postrictal; borde superior del tímpano superpuesto por el pliegue; tres tubérculos postrictales presentes y varios de poco relieve; membrana timpánica presente; anillo timpánico visible, ovalado, su longitud es el 66.6% de la longitud horizontal del ojo; coanas pequeñas, redondeadas, ligeramente cubiertas en la parte superior por el piso palatal; procesos vomerianos pequeños, ligeramente redondeados, dientes diferenciables (4–5), redondeados y agrupados medialmente; lengua ligeramente más ancha que larga, con el borde posterior bilobulado, la mitad anterior está adherida al piso de la boca. Piel del dorso finamente tuberculada; pliegues dorsolaterales presentes; superficie ventral areolada; pliegue discoidal presente; cubierta cloacal pequeña; tubérculos cloacales ausentes. Tubérculos escapulares presentes; tubérculos pequeños no pronunciados esparcidos en la región ulnar, ausentes en el codo. Superficie palmar ligeramente amplia; tubérculo palmar redondeado y elevado (Figura (A)), tubérculo tenar grande y alargado, aproximadamente 1/2 del ancho del tubérculo palmar; tubérculos supernumerarios grandes, aplanados y dispersos; tubérculos subarticulares pequeños y aplanados; un surco profundo y angosto se extiende desde la base anterior del dedo manual I hasta la base posterior del tubérculo palmar; dedos manuales con rebordes cutáneos estrechos, sin membranas interdigitales; dedo I ligeramente más corto que el dedo II; disco del dedo I ligeramente expandido, discos de los dedos II–IV ligeramente expandidos y redondeados (Figura (A)); todos los dedos manuales con cojinetes ventrales definidos por hendiduras circunferenciales. Extremidades posteriores poco robustas, longitud de la tibia 35.8% de la longitud rostro-cloaca; talón con tubérculos pequeños; tubérculos pequeños no protuberantes y dispersos a lo largo del borde externo del tarso; tubérculo metatarsal interno agrandado, ligeramente elevado y ovalado, dos veces más grande que el tubérculo metatarsal externo; tubérculos supernumerarios plantares agrandados y aplanados; tubérculos subarticulares definidos, agrandados, redondeados y elevados; dedos pediales con rebordes cutáneos estrechos; discos de los dedos pediales I–II de similar tamaño que los discos de los dedos manuales, mientras que los discos pediales III–V son ligeramente de menor tamaño, los discos están definidos por hendiduras circunferenciales; longitud relativa de los dedos del pie: I < II < III < V < IV; dedo III del pie ligeramente más pequeño que el dedo V; dedo III del pie llega al borde anterior del segundo tubérculo subarticular del dedo IV (Figura (B)).
Figura 18. Pristimantis saturninoi sp. nov. (DHMECN 12232). (A) Vista palmar de la mano; (B) vista plantar del pie. Longitud del pie 8.8 mm.
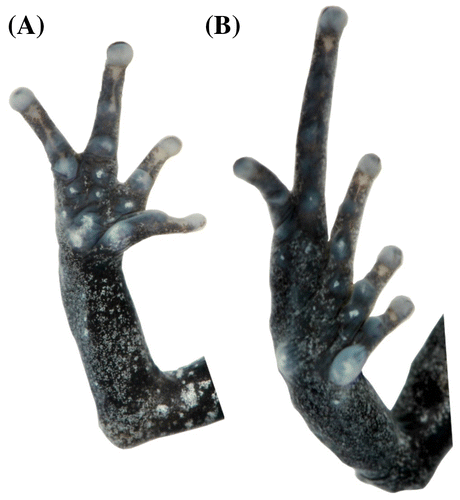
Caracteres craneales
A nivel dorsal el cráneo presenta una forma redondeada. El hueso nasal es delgado y cubre toda la parte anterior del esfenetmoide. Esfenetmoide poco robusto, con su región basal en forma de media luna (Figura (A)). Frontoparietal con bordes posteriores inclinados, fontanelas poco profundas pero evidentes. Otoccipital con cresta parótica evidente, cóndilos occipitales pequeños.
Figura 19. Vista dorsal y ventral del cráneo. (A) Pristimantis saturninoi DHMECN 12249, macho adulto; (B) P. orestes (redibujado de Lynch, 1979) y (C) P. vidua (redibujado de Lynch, 1979). Barra = 5 mm.
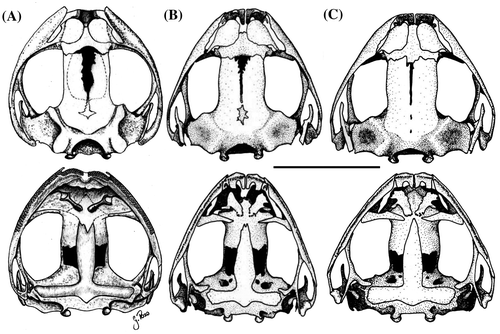
A nivel ventral presenta una forma redondeada. Maxila con dos filas de dientes, la primera fila presenta los dientes más evidentes; distalmente se articula por superposición al cuadradoyugal. Vómers formado por una placa palatal delgada, redondeado y separado medialmente por un espacio similar a su anchura; odontóforos diferenciables (4–5) y agrupados medialmente. Paresfenoide con forma de cruz invertida. En la región anterior, el processus cultriforme se articula con el esfenetmoide en su extremo distal, el cual es ancho y con borde irregular; los bordes laterales son irregulares y su parte media, que es aguda, no alcanza el foramen magnum. En la región posterior, el processus alaris tiene forma romboidal irregular que llega hasta el extremo lateral del proótico, recubriendo las cápsulas auditivas.
Coloración del holotipo en preservado
Dorso negro opaco (Figura (A)), con tonalidades gris cenizo hacia los flancos y cabeza, con finas maculaciones blanquecinas distribuidas a lo largo del cuerpo. Región rostral más clara que el dorso, con bandas marrones; tímpano amarillo vainilla, con área supratimpánica marrón oscura. Vientre negro, con tonalidades marrones hacia la garganta y la región gular (Figura (B)); granulaciones blanquecinas y grisáceas extendidas desde los brazos, hasta los muslos. Superficie dorsal de las extremidades anteriores con un mismo patrón de coloración que el cuerpo, con tonalidades marrón opacas, ligeramente cremas hacia el borde externo; extremidades posteriores del mismo patrón de coloración que las anteriores, con tonos blanquecinos hacia los tarsos. Superficie ventral de las extremidades anteriores opacas, entre tonalidades negras y marrones; extremidades posteriores con el mismo patrón de coloración que el vientre a nivel del muslo, y grisáceas hacia las piernas y tarsos. Tubérculos de la superficie palmar y plantar blanquecinos.
Figura 20. Pristimantis saturninoi sp. nov., en preservado. (A) Vista dorsal; (B) vista ventral del holotipo DHMECN 12232, hembra adulta, LRC: 21.5 mm.

Coloración del holotipo en vida
Dorso azabache (Figura (A)), con manchas ligeramente pardas, más claras y evidentes hacia la región dorsolateral. Región cefálica de la misma coloración del dorso, exceptuando el área maxilar, donde presenta manchas marrón claras dispuestas a manera de barras; tímpano dicromático entre azabache y marrón bronceado (windsor tan). Párpado superior con manchas marrón claras, con ligeros tonos verduzcos hacia la región preorbital. Superficie dorsal de las extremidades anteriores y posteriores ocres oscuras, ligeramente marrón rojizo; dedos manuales y pediales levemente más claros. Vientre de la misma coloración del dorso (azabache), con granulaciones gris plata, que se extienden desde los brazos, siguiendo a lo largo de la superficie ventral del cuerpo hasta los muslos (Figura (A)). Ingle y superficie anterior de los muslos y pantorrillas de color negro y con manchas agrandadas e irregulares de color blanquecino a crema amarillento; superficie palmar y plantar de los miembros manuales y pediales más claros en relación a las extremidades, en especial en el extremo distal de los dedos. Iris verde bronce con tonalidades marrones y amarillas (bronce antiguo).
Figura 21. Pristimantis saturninoi sp. nov., en vivo. Vista dorsal y ventral. (A) Hembra adulta DHMECN 12232, LRC: 21.5 mm; (B) hembra adulta, DHMECN 12235, LRC: 19.5 mm; C macho adulto, DHMECN 12231, LCR: 18.7 mm.

Medidas del holotipo (mm). Longitud rostro-cloaca = 21.5; ancho cefálico = 8.4; longitud cefálica = 8.6; longitud de la tibia = 7.7; longitud del pie = 8.8; longitud horizontal del ojo = 2.4; distancia órbita-narina = 1.5; distancia internarinal = 2.4; distancia interorbital = 2.6; diámetro del tímpano = 1.6; longitud de la mano = 5.3; ancho del párpado = 1.8.
Variación
La variabilidad de las medidas morfométricas se presenta en la Tabla . Variación cromática observada en el paratipo hembra adulta DHMECN 12235 (Figura (B)), presentó una distintiva coloración café en los flancos y la superficie ventral de la mano y pie de color ladrillo.
Distribución y ecología
La especie nueva es conocida únicamente de la localidad tipo ˝Cerro Sambalán˝, en el Parque Nacional Sangay, provincia de Morona Santiago, Ecuador, a una elevación entre 3000 y 3300 msnm. El piso zoogeográfico donde habita Pristimantis saturninoi pertenece al Piso Templado [Citation37], el ecosistema corresponde al Bosque siempreverde montano alto del Sur de la Cordillera Oriental de los Andes [Citation38], el cual se caracteriza por árboles cuyas ramas nudosas albergan gran cantidad de orquídeas, musgos y líquenes; en general la altura de la vegetación emergente alcanza hasta 15 m de altura, donde dominan especies de las familias Clusiacae, Melastomataceae, Asteraceae y Araliaceae. Pristimantis saturninoi fue colectado en las primeras horas de la noche (19:00–22:00), perchados sobre pequeñas hojas y ramitas entre los 0.2 y 0.6 m sobre el suelo. En las cercanías del hábitat de Pristimantis saturninoi se registró a tres especies de cutines Pristimantis no identificadas.
Descripción de las llamadas
Se analizó una muestra de 94 notas de 53 cantos grabados a los ejemplares DHMECN 12231–12239. Se caracterizan por ser de frecuencia modulada, cuya frecuencia dominante es de = 3.03 ± 0.07 kHz; presentan parciales armónicos de los cuales el segundo tiene una frecuencia de
= 6 ± 0.16 kHz, el tercero una frecuencia de
= 8.97 ± 0.26 kHz, y el cuarto una frecuencia de
=11.98 ± 0.34. Los cantos tienen una duración de
= 356.88 ± 391.26 ms, con intervalos de
= 3002.92 ± 1082.65 ms, emitiendo
= 20.44 ± 8.45 cantos/minuto. Están conformados por
= 1.77 ± 0.99 notas, las mismas que tienen una duración de
= 48.62 ± 12.03 ms, con intervalos de
= 350.31 ± 39.91 ms, emitiendo
= 2.57 ± 2.57 notas/segundo (Figura ; Tabla ).
Figura 22. Oscilograma y espectrogramas del llamado de Pristimantis saturninoi sp. nov. Macho adulto, DHMECN 12231, LCR: 18.7 mm.
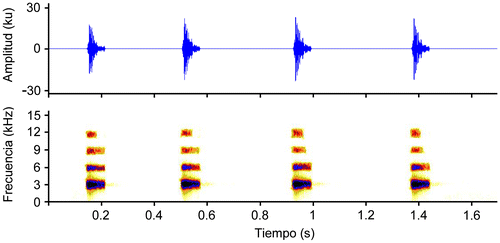
Tabla 3. Resumen de los análisis acústicos de los llamados de Pristimantis saturninoi sp. nov. La muestra analizada (n) corresponde a llamados/notas. Las abreviaciones utilizadas en los parámetros corresponden a: kHz = kilohertz, ms = milisegundos.
Comentarios taxonómicos
Nosotros no asignamos a Pristimantis saturninoi sp. nov., a ningún grupo de especies; aunque morfológicamente se relaciona con el grupo P. orestes (sensu Hedges et al. [Citation22] y Brito et al. [Citation66] por presentar los siguientes caracteres: tamaño corporal pequeño (<21.5 mm), hocico corto, cuerpo robusto, patas cortas y dedos relativamente cortos con discos reducidos. No obstante, Brito et al. [Citation66] mencionan que el morfotipo de P. orestes ha evolucionado independientemente en otro clado de Pristimantis como el grupo P. myersi, por lo tanto, futuros estudios filogenéticos que incorporen secuencias de ADN ayudarán a consolidar a P. saturninoi dentro de un grupo, evitando problemas de polifilia.
Pristimantis tinguichaca [Citation16] (Figura 14(G))
Especie endémica de Ecuador, recientemente descrita. [Citation16] En la localidad de Guabisai y Tinguichaca, P. tinguichaca es un habitante del bosque montano entre los 2300 y 2900 m de altitud; prefiriendo el estrato bajo del bosque, áreas abiertas o bordes de quebradas entre los 0.5 y 1.3 m sobre el suelo; la mayor actividad vocal de los machos suele darse durante las noches con lluvia ligera. El estado de conservación aún no ha sido evaluado
Material de referencia: DHMECN 12283–300.
Pristimantis trachyblepharis (Boulenger, 1918) (Figura 14(H))
Especie endémica de la cuenca alta del río Pastaza distribuida entre 950 y 1500 m de elevación [Citation2,42,45]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. En Danu y río Sardinayacu ha sido registrada al interior del bosque maduro entre 1350 y 1400 m de elevación. Percha en la vegetación herbácea en el interior del bosque prístino entre 0.2 y 0.6 m sobre el suelo. En Ecuador [Citation6] se lo considera como una especie en Preocupación Menor y a nivel global [Citation8] con Datos Deficientes.
Material de referencia: MEPN 14510–2, 14057–8.
Pristimantis ventrimarmoratus (Boulenger, 1912) (Figura 14(I))
Es una especie ampliamente distribuida en los bosques nublados de la vertiente amazónica de los Andes desde el norte de Ecuador, Perú y parte adyacente de Bolivia, se presume su presencia en los bosques al oeste de Brazil entre 0 y 1800 m de elevación [Citation2,5]. Padial et al. [Citation24] no lo asignan a ningún grupo de especies. En el río Sardinayacu ha sido localizado perchando en la vegetación herbácea, en el interior del bosque prístino entre 1350 y 1450 m de elevación. En Ecuador [Citation6] y a nivel global [Citation8] lo consideran como una especie con Preocupación Menor.
Material de referencia: DHMECN 12386–90, MEPN 14674.
Discusión
Los primeros estudios para documentar la diversidad de ranas terrestres Pristimantis de los bosques montanos de la Cuenca alta del río Santiago (subcuenca del Zamora y Paute), entre 1000 y 3500 m, fueron realizados por Lynch [Citation67] y Lynch and Duellman [Citation45] reportando un total de 17 especies. Sumando las 37 especies de la SARU aquí reportadas e integrando especies de publicaciones recientes [Citation51], estimamos la presencia de 48 taxa de Pristimantis para la Cuenca alta del Santiago (ver Apéndice 2). Se observa que ésta diversidad iguala en número a lo reportado para las estribaciones occidentales del Distrito Metropolitano de Quito [Citation36], además de ser superior a las 43 especies reportadas en la Cordillera Oriental del norte de Perú y a las 33 especies de la Cordillera Occidental del mismo país [Citation5]. Solamente la Cordillera del Cóndor [Citation50], una región aislada de los Andes (suroriente de Ecuador y noroccidente de Perú) posee tal diversidad de ranas Pristimantis que despunta a la Cuenca alta del Santiago, en una gradiente desde los 800–2860 m, alberga a 55 especies. No obstante, en la Cuenca Alta del Santiago existen varias áreas con vacíos de información (e. g. cabeceras de los ríos Negro, Paute, Tutanangosa, Abanico y Sangay) que con futuros muestreos podrían incrementar el número de especies.
En la subcuenca Alta del río Upano las especies de Pristimantis se reúnen en cinco grupos de especies (sensu Padial et al. [Citation24]): (1) Pristimantis ridens (P. almendariz, P. lanthanites y P. rubicundus); (2) Pristimantis myersi (P. albujai, P. bicantus y P. sambalan); (3) Pristimantis lacrimosus (P. lacrimosus, P. latericius, y P. aff. petersi); (4) Pristimantis conspicillatus que acopia una especie con su mismo nombre; y (5) Pristimantis frater (P. churuwiai y P. quaquaversus); mientras 17 especies que representan el 43%, carecen de pertenencia a algún grupo (P. altamnis, P. delius, P. diadematus, P. gagliardoi, P. incomptus, P. inusitatus, P. katoptroides, P. nigrogriseus, P. prolatus, P. proserpens, P. quaquaversus, P. roni, P. orestes, P. saturninoi, P. trachiplepharis y P. ventrimarmoratus).
La importancia biogeográfica de la SARU se demuestra con el registro de 37 especies de ranas Pristimantis, representando el 46% (80 spp) reportadas para la Cuenca Alta de los ríos Napo, Pastaza y Santiago; 26 especies son endémicas, que representa el 51% de endemismo de la vertiente oriental [Citation7].
En conclusión, nuestros resultados proporcionan el primer esfuerzo para representar los patrones de diversidad de las ranas Pristimantis en éstos ecosistemas poco conocidos y altamente diversos. La información presentada en nuestro trabajo es esencial para futuros estudios biogeográficos, ecológicos o evolutivos, además de facilitar datos de las fajas altitudinales con más especies amenazadas, que pudieran dar las pautas para emprender programas de conservación de las especies amenazadas de extinción.
Contribución de los autores
JB y DB concibieron el estudio. JB y MYM realizaron la descripción de las especies nuevas. DB realizo la descripción de la coloración y el análisis de las vocalizaciones. MYM realizó los análisis de los patrones de diversidad. JB realizo el compendio de las especies y escribió el primer borrador del manuscrito. MYM y DB realizaron revisión crítica del manuscrito. Todos los autores aprobaron la versión final del manuscrito.
Agradecimientos
Los funcionarios del Parque Nacional Sangay zona baja, en especial V. León brindaron las facilidades necesarias para llevar a cabo esta investigación. A los guardaparques E. Chacha, G. Tenecota, E. Marín, T. Shacay y M. Sharup que colaboraron con la logística para el ingreso a los diferentes sitios. A nuestros colegas y amigos F. Anaguano, P. Bejarano, R. Ojala-Barbour, J. Curay, R. Vargas, H. Orellana y G. Pozo por su incondicional ayuda en el trabajo de campo. A G. Pozo por la elaboración de la Figura . Nuestro reconocimiento a L. Rivadeneira (Domono), C. Tenecota y L. Piña (Nueva Alianza), M. Chuqui (Guabisai) por su ayuda y gentileza al hospedarnos en sus hogares. A. Almendáriz (EPN) y C. Reyes-Puig (MECN) colaboraron con las facilidades para el trabajo de laboratorio. Al Ministerio del Ambiente de Morona Santiago quien otorgó el permiso de investigación N°. 05-2014-I-B-DPMS/MAE.
Conflicto de intereses
Los autores no presentan conflicto de intereses.
Financiamiento
El estudio fue realizado con el apoyo financiero del Museo Ecuatoriano de Ciencias Naturales, la Fundación Naturaleza Kakaram y con fondos personales de JB y DB.
Associate Editor: Juan M. Guayasamin
Related Research Data
Referencias
- Frost DR. Amphibian species of the world: an online reference [Internet]. Version 6.0. New York: American Museum of Natural History; 2016 [cited 2017 Jan 18]. Available from: http://research.amnh.org/herpetology/amphibia/index.html
- Ron SR, Guayasamin JM, Yánez-Muñoz MH, et al. AmphibiaWebEcuador [Internet]. Version 2016.0. Quito: Museo de Zoología, Pontificia Universidad Católica del Ecuador; 2016 [cited 2016 Jan 18] Available from: http://zoologia.puce.edu.ec/Vertebrados/anfibios
- Lynch JD, Duellman WE. Frogs of the genus Eleutherodactylus in western Ecuador. Systematics, ecology, and biogeography. Special public. Univ Kansas Nat Hist Mus. 1997;23:1–236.
- Mendoza AM, Ospina OE, Cárdenas-Henao H, et al. A likelihood inference of historical biogeography in the world’s most diverse terrestrial vertebrate genus: Diversification of direct-developing frogs (Craugastoridae: Pristimantis) across the Neotropics. Mol Phylogenet Evol. 2015;85:50–58.
- Duellman ED, Lehr E. Terrestial breeding frogs (Strabomantidae) in Peru. Berlin: Ulrich Manthey; 2009.
- Ron SR, Guayasamin JM, Menéndez-Guerrero P. Biodiversity and conservation status of Ecuadorian amphibians. En: Heatwole H, Barrio-Amoros CL, Wilkinson HW, editors. Amphi Biol. 2011;9(2):129–170.
- Nori J, Lemes P, Urbina-Cardona N, et al. Amphibian conservation, land-use changes and protected areas: a global overview. Biol Cons. 2015;191:367–374.
- UICN (Unión Internacional para la Conservación de la Naturaleza). Categorías y criterios de la lista roja de la UICN, versión 3.1. Segunda edición. Gland, Suiza: UICN; 2012.
- Cerón C, Montalvo C. Aspectos Florísticos, Diversidad y Ecología del Parque Nacional Sangay, Ecuador. Cinchonia. 2006;7:1–10.
- Brito J, Almendáriz A. Anfibios y Reptiles del Parque Nacional Sangay. Rapid Color Guide. 2013;502:1–9.
- Brito J, Ojala-Barbour R. Mamíferos no voladores del Parque Nacional Sangay, Ecuador. Pap Avulsos Zool. 2016;56(5):45–61.
- Ojala-Barbour R, Pinto M, Brito J, et al. A new species of northern shrew-opossum (Paucituberculata: Caenolestidae) with a phylogegy of extant caenolestids. J Mammal. 2013;94(5):967–982.
- Harvey M, Almendáriz A, Brito J, et al. A new species of Noblella (Anura: Craugastoridae) from the Amazonian Slopes of the Ecuadorian Andes with Comments on Noblella lochites (Lynch). Zootaxa. 2013;3635(1):1–14.
- Batallas D, Brito J. Nueva especie de rana del género Pristimantis del grupo lacrimosus (Amphibia: Craugastoridae) del Parque Nacional Sangay, Ecuador. Pap Avulsos Zool. 2014;54:51–62.
- Yánez-Muñoz M, Bejarano-Muñoz P, Brito J, et al. Ranas terrestres de los Andes Surorientales de Ecuador II: Una nueva especie de Pristimantis verde espinosa de los bosques montanos del Parque Nacional Sangay (Anura: Craugastoridae). Av Cienc Ing. 2014;6(2):B63–B77.
- Brito J, Ojala-Barbour R, Batallas D, et al. A new species of Pristimantis (Amphibia: Strabomantidae) from the cloud forests of Sangay National Park, Ecuador. J Herpetol. 2016;50(2):337–344.
- Brito J, Tinoco N, Chávez D, et al. New species of arboreal rat of the genus Rhipidomys (Cricetidae, Sigmodontinae) from Sangay National Park, Ecuador. Neotropical Biodiversity. 2017;3(1):65−79.
- Larsen TH, editor. Core standardized methods for rapid biological field assessment. Arlington (VA): Conservation International; 2016.
- Dodd Jr CK, editor. Amphibian ecology and conservation. A handbook of techniques. Oxford: Oxford University Press; 2010.
- Heyer WR, Donnelly MA, McDiarmid RW, et al., editors. Measuring and monitoring biological diversity: standard methods for amphibians. Washington: Smithsonian Institution Press; 1994.
- Hammer Ø, Harper DAT, Ryan PD. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electron. 2001;4(1): 9pp. Available from: http://palaeo-electronica.org/2001_1/past/issue1_01.htm
- Hedges SB, Duellman WE, Heinicke MP. New world direct-developing frogs (Anura: Terrarana): molecular phylogeny, classification, biogeography and conservation. Zootaxa. 2008;1737:1–182.
- Pyron RA, Wiens JJ. A large-scale phylogeny of Amphibia including over 2,800 species, and a revised classification of extant frogs, salamanders, and caecilians. Mol Phylogenet Evol. 2011;61:543–583.
- Padial JM, Grant T, Frost DR. Molecular systematics of terraranas (Anura: Brachycephaloidea) with an assessment of the effects of alignment and optimality criteria. Zootaxa. 2014;3825(1):1–132.
- Rueda JV, Lynch JD. Una nueva especie de Eleutherodactylus (Amphibia: Leptodactylidae) para la cordillera oriental de Colombia. Lozania. 1983;42:1–6.
- Chen HM, Combs CA. An alternative anesthesia for amphibians: ventral application of benzocaine. Herpetol Rev. 1999;30:34.
- Simmons JE. Herpetological collecting and collection management. Herpetol Cir. 2002;31:1–153.
- Encycolorpedia. Lista extendida de colores. [Internet]; 2016; [cited 2016 May 10]. Available from: http://encycolorpedia.es.
- Charif RA, Waack AM, Strickman LM. Raven Pro 1.4 user’s manual. Ithaca (NY): Cornell Lab of Ornithology; 2010.
- Duellman WE, Pyles RA. Acoustic resource partitioning in Anuran communities. Copeia. 1983;639–649.
- Cocroft RB, Ryan MJ. Patterns of advertisement call evolution in toads and chorus frogs. Anim Behav. 1995;49:283–303.
- Angulo A. Fundamentos de bioacústica y aspectos prácticos de grabaciones y análisis de cantos. En: Angulo A, Rueda-Almondacid JV, Rodriguez-Mahecha JV, Marca E, editors. Técnicas de inventario y monitoreo para los anfibios de la Región Tropical Andina. Bogotá: Conservación Internacional; 2006. p. 93–134.
- Díaz LM, Cádiz LM. Guía descriptiva para la identificación de las llamadas de anuncio de las ranas cubanas del género Eleutherodactylus (Anura: Leptodactylidae). Herpetotropicos. 2007;3:100–122.
- Lynch JD, Burrowes PA. The frogs of the genus Eleutherodactylus (Family: Leptodactylidae) at the La Planada Reserve in southwestern Colombia with descriptions of eight new species. Occas Pap Mus Nat Hist Univ Kansas. 1990;136:1−31.
- Guayasamin JM, Krynak T, Krynak K. Phenotypic plasticity raises questions for taxonomically important traits: a remarkable new Andean rainfrog (Pristimantis) with the ability to change skin texture. Zool J Linn Soc. 2015;173:913–928.
- Yánez-Muñoz MH, Bejarano-Muñoz EP. Lista actualizada de ranas terrestres Pristimantis (Anura: Craugastoridae) en las Estribaciones Occidentales del Distrito Metropolitano de Quito, Andes de Ecuador. Bol Téc Serie Zool. 2013;11:125–150.
- Albuja L, Almendáriz A, Barriga R, et al. Fauna de Vertebrados del Ecuador. Escuela Politécnica Nacional. Quito: Editorial Arial 12; 2012.
- Galeas R, Guevara J, Medina-Torres B, et al., editors. Sistema de clasificación de los ecosistemas del Ecuador continental. Quito: Ministerio del Ambiente del Ecuador; 2013.
- Brito J, Pozo-Zamora G. Una nueva especie de rana terrestre del género Pristimantis (Amphibia: Craugastoridae), de la Cordillera de Kutukú, Ecuador. Pap Avulsos Zool. 2013;53(24):315–325.
- Elmer KR, Cannatella DC. Three new species of leaflitter frogs from the upper Amazon forests: cryptic diversity within Pristimantis “ockendeni” (Anura: Strabomantidae) in Ecuador. Zootaxa. 2008;1784:11−38.
- Guayasamin JM, Funk CW. The amphibian community at Yanayacu Biological Station, Ecuador, with a comparison of vertical microhabitat use among Pristimantis species and the description of a new species of the Pristimantis myersi group. Zootaxa. 2009;2220:41–66.
- Reyes-Puig M, Reyes-Puig J, Yánez-Muñoz MH. Ranas terrestres del género Pristimantis (Anura: Craugastoridae) de la Reserva Ecológica Río Zúñag, Tungurahua, Ecuador: Lista anotada y descripción de una especie nueva. Av Cienc Ing. 2013;5(2):B5–13.
- Duellman WE, Pramuk JB. Frogs of the Genus Eleutherodactylus (Anura: Leptodactylidae) in the Andes of Northern Peru. Sci Pap Nat Hist Mus Univ Kansas. 1999;13:1–78.
- Lynch JD. New species of frogs (Leptodactylidae: Eleutherodactylus from the Amazonian lowlands of Ecuador. Occas Pap Mus Nat His Univ Kansas. 1974;31:1–22.
- Lynch JD, Duellman WE. The Eleutherodactylus of the Amazonian slopes of the Ecuadorian Andes (Anura: Leptodactylidae). Misc Publ Univ Kansas Mus Nat Hist. 1980;69:1–86.
- Ortega-Andrade HM, Venegas PJ. A new synonym for Pristimantis luscombei (Duellman and Mendelson 1995) and the description of a new species of Pristimantis from the upper Amazon basin (Amphibia: Craugastoridae). Zootaxa. 2014;3895:031–057.
- Ortega-Andrade HM, Valencia JH. A new species of the Pristimantis frater group (Anura: Strabomantidae) from the eastern evergreen lowland forests of Ecuador. Herpetologica. 2012;68:244–255.
- Lynch JD. Leptodactylid frogs of the genus Eleutherodactylus in the Andes of Northern Ecuador and adjacent Colombia. Univ Kansas Mus Nat Misc Publ. 1981;72:1–46.
- Duellman WE, Mendelson JR. Amphibians and reptiles from northern department Loreto, Peru: Taxonomy and biogeography. Univ Kans Sci Bull. 1995;55:329–376.
- Almendáriz A, Simmons J, Brito J, et al. Overview of the herpetofauna of the unexplored Cordillera del Cóndor of Ecuador. Amphib Reptile Conserv. 2014;8(1):45–64(e82).
- Bustamante MR, Mendelson JR III. A new frog species (Strabomantidae: Pristimantis) from the high Andes of southeastern Ecuador. Zootaxa. 2008;1820:49–59.
- Coloma LA, Guayasamin JM, Menéndez-Guerrero P, editors. Lista Roja de Anfibios de Ecuador. AnfibiosWebEcuador. Base de datos electrónica disponible en http://www.anfibioswebecuador.ec/index.php?lr,10FundaciónOtonga. Quito, Ecuador; 2011–2015. [Cited 2017 Jan 19].
- Duellman WE, Lynch JD. Anuran amphibians from the Cordillera de Cutucú, Ecuado. Proc Acad Nat Sci Philadelphia. 1988;140(2):125–142.
- Batallas D, Brito J. Descripción de las vocalizaciones de Pristimantis ganonotus (Anura: Craugastoridae) de Ecuador. Cuadernos de Herpetología. 2016;30(1):31–34.
- Urgilés V, Sánchez-Nivicela J, Nieves C, et al. Ranas terrestres en los ecosistemas surandinos de Ecuador I: Dos nuevas especies de Pristimantis (Anura: Craugastoridae) de la ladera oriental. Av Cien Ing. 2014;6:B51–B59.
- Flores G. Two new species of Ecuadorian Eleutherodactylus (Leptodactylidae) of the E. Crucifer assembly. J Herpetol. 1988;22:34–41.
- Catenazzi A, Venegas P. Anfibios y reptiles. en: Pitman N, Ruelas E, Alvira D, et al, editors. Perú: Cerros de Kampankis. Rapid biological and social inventories report 24. Chicago: The Field Museum; 2012. p. 106–117.
- Cusi JC, Barboza AC, von May R. A new locality, range extension and record of Batrachochytrium dendrobatidis in the endangered terrestrial breeding frog Pristimantis katoptroides Flores, 1988 (Anura: Craugastoridae) in Peru. Check List. 2015;11(2):1608.
- Chaparro JC, Jiménez-Robles O, Brito J, et al. Anfibios y Reptiles de la cordillera del Cutucú, Ecuador. Con el apoyo de: Universidad Internacional Menéndez Pelayo (UIMP), Universidad Tecnológica Indoamerica (UTI), Consejo Superior de Investigaciones Científicas (CSIC), Máster en Biodiversidad en Áreas Tropicales y su Conservación (MBATC), TheTinker Foundation (TF); 2011.
- Guayasamin JM, Ron SR, Cisneros-Heredia DF, et al. A new species of frog of the Eleutherodactylus lacrimosus assemblage (Leptodactylidae) from the western Amazon Basin, with comments on the utility of canopy surveys in lowland rainforest. Herpetologica. 2006;62:191–202.
- Batallas D, Brito J. Análisis bioacústico de las vocalizaciones de seis especies de anuros de la laguna Cormorán, complejo lacustre de Sardinayacu, Parque Nacional Sangay, Ecuador. Rev Mex Biodivers. 2016;87:1292–1300.
- Brito J, Batallas D, Velalcázar D. Nueva especie de rana terrestre del género Pristimantis (Amphibia: Craugastoridae), meseta de la Cordillera del Cóndor. Pap Avulsos Zool. 2014;54(30):435–446.
- Rödder D, Schmitz A. Two new Pristimantis (Anura, Strabomantidae) belonging to the myersi group from the Andean slopes of Ecuador. Rev Suisse Zool. 2009;116:275–288.
- Rojas-Runjaic FJM, Delgado AJ, Guayasamin JM. A new rainfrog of the Pristimantis myersi Group (Amphibia, Craugastoridae) from Volcán Pichincha, Ecuador. Zootaxa. 2014;3780:36–50.
- Yánez-Muñoz M, Meza-Ramos P, Cisneros-Heredia DF, et al. Descripción de tres nuevas especies de ranas del género Pristimantis (Anura: Terrarana: Craugastoridae) de los bosque nublados del Distrito Metropolitano de Quito, Ecuador. Av Cienc Ing. 2010;2(3):B16–B27.
- Brito J, Almendáriz A, Ron SR, et al. Nueva especie de rana bromelícola del género Pristimantis (Amphibia: Craugastoridae), meseta de la Cordillera del Cóndor, Ecuador. Pap Avulsos Zool. En preparación.
- Lynch JD. Leptodactylid frogs of the genus Eleutherodactylus from the Andes of Southern Ecuador. Misc Publ Nat Hist Mus Univ Kansas. 1979;66:1–62.
- Lehr H, Coloma LA. A minute new Ecuadorian Andean frog (Anura: Strabomantidae, Pristimantis). Herpetologica. 2008;64(3):354–367.
- Arteaga-Navarro AF, Guayasamin JM. A new frog of the genus Pristimantis (Amphibia: Strabomantidae) from the high Andes of Southeastern Ecuador, discovered using morphological and molecular data. Zootaxa. 2011;2876:17–29.
- Guayasamin JM, Arteaga AF. A new species of the Pristimantis orestes group (Amphibia: Strabomantidae) from the high Andes of Ecuador, Reserva Mazar. Zootaxa. 2013;3616:345–346.
- Wiens JJ, Coloma LA. A new species of the Eleutherodactylus myersi (Anura: Leptodactylidae) assembly from Ecuador. J Herpetol. 1992;26:196–207.
- Reyes-Puig JP, Yánez-Muñoz MH. Una nueva especie de Pristimantis (Anura: Craugastoridae) del corredor ecológico Llangantes-Sangay, Andes de Ecuador. Pap Avulsos Zool. 2012;52(6):81–91.
- Navarrete MJ, Venegas PJ, Ron SR. Two new species of frogs of the genus Pristimantis from Llanganates National Park in Ecuador with comments on the regional diversity of Ecuadorian Pristimantis (Anura, Craugastoridae). ZooKeys. 2016;593:139–162.
- Reyes-Puig JP, Reyes-Puig C, Ramirez-Jaramillo SR, et al. Tres nuevas especies de ranas terrestres Pristimantis (Anura: Craugastoridae) de la cuenca alta del Río Pastaza, Ecuador. Av Cien Ing. 2014;6:B51–B62.
- Reyes-Puig JP, Reyes-Puig C, Pérez MB, et al. Dos nuevas especies de ranas Pristimantis (Craugastoridae) de la cordillera de los Sacha Llanganatis, vertiente oriental de los Andes de Ecuador. Av Cien Ing. 2015;7:B61–B74.
- Guayasamin JM, Almeida-Reinoso DP, Nogales-Sornosa F. Two new species of frogs (Leptodactylidae: Eleutherodactylus) from the high andes of northern Ecuador. Herpetological Monographs. 2004;18:127–141.
Apéndice 1.
Especímenes examinados, todos los especímenes son de Ecuador
Pristimantis andinognomus
Provincia Zamora Chinchipe: Reserva Tapichalaca (04°29′S, 79°08′W, elevación: 1800 a 3400m), DHMECN 1824-27, 1896, 2020, 2492, 2485-9, 2490, 2495, 2497, 2500, 2502-03 8107, 8135, 8207, 9667, 3041, 3776, 3781.
Pristimantis bicantus
Provincia de Napo: Microcuenca de los ríos Sardinas y Santa Rosa (0°22′32.92″S, 77°53′3.23″O, elevación: 2000m), DHMECN 9251-52; Baeza, Reserva Etnobotánica Cumandá (0°26′49.17″S, 77°52′56.53″O, elevación: 1920m), DHMECN 11891-92, 11897. Provincia de Tungurahua: Bosque Protector La Candelaria (01° 28′ 35.9′ S′ 78° 17 46.6 W, elevación, 1700-3800), DHMECN 4966-67, 4970; río Negro, DHMECN 5639.
Pristimantis colodactylus
Provincia de Loja: 13.5 km al este de Loja (3°58′3.79″S, 79° 3′20.72″O, elevación: 2800m): KU 142157 (Fotografías material tipo, AmphibiaWebEcuador).
Pristimantis floridus
Provincia de Imbabura: Reserva Alto Chocó (0°21′60.00″N, 78°28′60.00″O, elevación: 2500-2900m) DHMECN 2305, 2308, 2311, 2313, 2340-2349; Provincia de Pichincha: Reserva Maquipucuna, DHMECN 7249-7256 (0° 3′37.90″N, 78°34′45.10″O, elevación: 2600-2900m)
Pristimantis mazar
Provincia Azuay: Cuenca, Baños, Hacienda Yanasacha (2°55′5.41″S, 79° 5′55.71″O, elevación: 2200m): MEPN 15017.
Pristimantis miktos
Provincia Orellana: Parque Nacional Yasuní, DHMECN 3365-66 (0°58′36.02″S, 75°26′41.85″O, elvación: 270m); Provincia de Pastaza: Comunidad Bufeo (2°12′47.52″S, 76°49′2.95″O, elevación: 300m): DHMECN 4448.
Pristimantis nigrogriseus
Provincia de Morona Santiago: Sardinayacu, Parque Nacional Sangay (2◦04′20.59″S, 78◦12′52.40″O, elevación 1750m): MEPN 14393-4
Pristimantis orestes
Provincia de Loja: San Antonio de Cumbe: MEPN 3481; 11 km NE Urdaneta (3°31′48.00″S, 79° 7′25.10″O, elevación: 2970m): KU 141998 (Fotografías material tipo, AmphibiaWebEcuador).
Pristimantis proserpens
Provincia Zamora Chinchipe: Paquisha, Paquisha Alto (78°28′54.1″W; 03°54′05.1″S, elevación: 2300m): MEPN 11416, 11420, 11426, 11428˗9, 12352, 12361, 12372,12369, 12367, 12353, 12360, 13591, 14262, 14261, 14264, 15323–26: 1811 msnm.
Pristimantis quaquaversus
Provincia de Sucumbios: Río Verde, DHMECN 6355; Provincia Orellana: Kawimwro-OBE (1°8′17.76″S, 75°53′15.80″O, elevación202m), DHMECN 3116-17; Provincia de Tungurahua: Reserva río Zuñac (1°22′31.27″S, 78°9′43.49″O, ELEVACIÓN: 1400m) DHMECN 5602, 5154-5191; Provincia de Pastaza: Mera, Reserva Ecológica río Anzu (01° 24′32.4″S; 78°02′45.4″, elevación 1300 m) DHMECN 7040-43.
Pristimantis verecundus
Provincia de Pichincha: Lloa, vía Lloa-Mindo (0° 7′44.10″S, 78°45′15.70″O, elevación: 1390m) DHMECN 7399-7406, 7437-39; Nanegal, Curipogio (0° 7′52.01″N, 78°40′34.76″O, elevación: 1800) DHMECN 1927; Provincia de Imbabura, García Moreno, Junin (0°18′31.25″N, 78°39′52.96″O, elevación: 1700m) DHMECN 3532.