
ABSTRACT
Moriche palm (Mauritia flexuosa Linn f. [Arecaceae]) is a dominant species in the savanna margins of the Caribbean island of Trinidad. The savanna margins are frequently subjected to fires and the moriche palm, like many palm species, is adapted to cope with a degree of fire damage. These palms also frequently host large termite nests that tend to burn during fires, which causes further damage to the palm stems. This paper explores the possibility that the combination of termite nests and frequent fires can increase tree mortality. Two post-fire surveys were conducted to record, evidence of fire exposure, fire damage to termite nests and fire damage to palm stems. In both surveys, the majority of palms with termite nests suffered fire damage to their stems, while none of the termite-free palms showed damage. Furthermore, a comparison of nest size and damage to the bark showed stems with larger nests had significantly more damage than those with smaller or no nests. The results suggest that the presence of termite nests increases the extent of fire damage to palm trunks by providing a point of combustion that focuses flames on the already compromised stem tissue. Long-term studies are warranted to investigate the impact of repeated fires on wild arborescent palm populations.
Mauritia flexuosa Linn f. (Arecaceae) or moriche palm is common in the Neotropics, in swamps and temporally flooded habitats, and is considered a hyper-keystone species [Citation1]. It is also common in many hyper-seasonal savannas (e.g. Grand Sabana, Cerrado, Rupununi, and Sipaliwini), especially along the margins [Citation2–4]. On the island of Trinidad, off the north coast of the South American mainland, moriche palm can be found in great abundance on the savanna margins that border the marsh forest in the Aripo Savanna Environmentally Sensitive Area (ASESA) [Citation5–8]. The ASESA (10°35´30´´ N, 61°12´0´´ W) is an edaphic white sand savanna-forest system that is located at the base of Trinidad’s northern range mountains at approximately 45 meters above sea level [Citation9]. The ASESA consists of 7 open savannas contained within a system of marsh forests; the individual savannas are more than 0.02 km2 in area and are dominated by grass-sedge vegetation such as Lagenocarpus rigidus and Paspalum pulchellum [Citation2]. Savannas 1 and 3 are amongst the largest (approximately 0.75 km2) and are the most susceptible to fires due to their close proximity to human settlements; the sole source of fire ignition in the ASESA ().
Figure 1. Location and position of transect lines for survey 1 and survey 2 in the Aripo Savannas environmentally sensitive area (ASESA), Trinidad
In the ASESA, this palm is often found in association with arboreal nests of the termite Nasutitermes ephratae (Holmgren) [Citation2] ()), as occurs in other sites [Citation10–12]. Generally, these nests are not considered to cause significant damage to host palms because Nasutitermes termites are polyletic consumers of cellulose that prefer to feed on dead wood [Citation11]. Nonetheless, Nasutitermes termites are likely to cause some damage to the palms because they have been observed to consume both living and dead tissue from fruits, stems, and roots [Citation11–14]. This paper describes a more severe form of damage that occurs when the presence of arboreal nests is combined with the occurrence of wildfire.
Figure 2. Images of non-fire-exposed and fire-exposed palm stems after the first fire in March 2014 along Transect Line 1(a-d), and damages to fire-exposed stems after the second fire along Transect Line 2 in 2017 (e-h). (a) non-fire-exposed Mauritia flexuosa stem with Nasutitermes ephratae nest. (b) fire-exposed M. flexuosa stem without N. ephratae nest. (c. & d) fire-exposed M. flexuosa stems showing severe damage that is associated with N. ephratae nest. (e-h) repeated burning of M. flexuosa palms appears to cause further damage, including the death of some palms

Wildfires have long been a part of tropical savanna systems [Citation15–17], and the ASESA is prone to anthropogenic fires that occur during the dry season at least once every 5–7 years [Citation5,Citation18,Citation19]. The effect of fires on the Aripo Savannas is still not fully understood [Citation19–21], but studies have suggested that the vast majority of mature M. flexuosa palms have been exposed to fire in the fire-prone western margins and island margins (i.e. the periphery of small palm stands that are surrounded by open savanna) [Citation5]. These margins are more fire-prone because the prevailing trade winds drive flames in that direction (). Wildfires can kill palm seedlings, however, adult M. flexuosa are considered to be fire-tolerant [Citation5] because they are protected by a rhytidome (dead outer hard bark [Citation22]) and have monocotyledonous vascular bundles with numerous water channels scattered throughout the stem [Citation23]. These anatomical structures (i.e. the rhytidome and scattered vascular bundles) make palms less vulnerable to surface damage [Citation22–24]. Additionally, the outer portion of the bark produces suberin (a complex polyester biopolymer) that prevents water loss [Citation25]. Typically, the bark of M. flexuosa does not burn away during a fire event [Citation26], instead after several months the scorched bark “flakes” away leaving the inside of the stem unaffected ()). However, the presence of termite nests appears to alter this course of events, as it results in more substantial damage to bark tissue during the fire ().
To better understand the extent of this phenomenon, fire occurrences in the ASESA were monitored from January 2014 to January 2018 by liaising with onsite forestry officers and conducting targeted site visits whenever a fire occurred (three fire events in total; occurring in two savannas). In January 2014, an intense fire that lasted for several days burned most of Savanna 1. Soon after the fire, we conducted a post-fire survey (Transect Line 1; ) within the fire-prone western and island margins of Savanna 1. In Transect Line 1, palm stems were observed over a 500 meter transect covering the extent of the fire along the savanna margin and encompassing a total of 138 palms. The presence of burnt or unburnt termite nests was recorded, as were visual estimates of fire occurrence (i.e. blacked bark and burnt understory) and fire damage (i.e. broken bark and broken cortex tissue). Burnt nests were classified according to signs of charring (i.e. partial to entire), whereas unburnt nests were classified as showing no sign of charring (i.e. having no evidence of exposure to fire).
In March 2014, a fire occurred in Savanna 3, and a post-fire survey was conducted (Transect Line 2). This fire affected a larger area and palms were surveyed over a 1,000 meter Transect Line along the savanna margin (Transect Line 2; ). As outlined above, the presence of burnt and unburnt termite nests (a total of 80 palms) was recorded along with visual estimates of fire occurrence and fire damage.
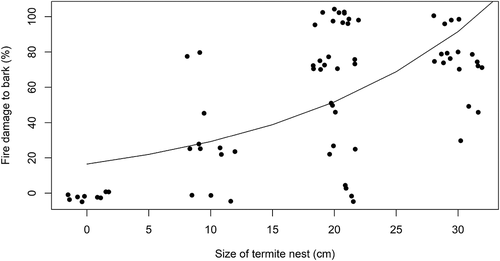
For palms that exhibited evidence of the fire on Transect Line 2, an additional assessment was made of the extent of fire damage to M. flexuosa bark by observing the circumference of the stem from ground level to 2.5 m on all sides and recording the proportion of the palm stem that was damaged (i.e. rhytidome burnt away and cortex exposed) to the nearest 25%. The size of each termite nest within this zone was also estimated as the maximum Feret diameter to the nearest 10 cm (i.e. 0–10, 10–20, 20–30 and >30). Sixty-nine palms in Transect Line 2 showed evidence of fire and had termite nests within the target zone, for these palms the association between the size of the termite nests (predictor variable) and the damage to the bark surface (response variable) was tested using a generalised linear model with a quasi-poisson distribution that was fitted using a maximum likelihood function in Rstudio (version 1.2.1335). In June 2017, another fire occurred in the same part of Savanna 3, after which Transect Line 2 was revisited, and further observations were made.
Along Transect Line 1, of the 138 adult palms, 95% hosted N. ephratae nests. Of these, 92% displayed evidence of fire, and 60% exhibit evidence of fire damage to the bark (i.e. rhytidome burnt away and cortex exposed). The seven palms without nests all exhibited evidence of fire, but none were damaged. During Transect Line 1 observations, it was noted that fire damage to the bark was concentrated in the areas where termite nests were attached to the trunks, particularly when nests were located low down on the trunks (i.e. < 2.5 m) and were exposed to savanna fires (). Typically, nests were extensively burnt during fires, and a considerable portion of the trunk was burnt as well. It appears that the nests provide a combustion point and focus the flames on the already compromised stem tissue. This observation was assessed more closely in the subsequent survey (i.e. Transect Line 2).
In Transect Line 2, 81% of the 80 adult palms hosted termite nests. Of these, 94% exhibited evidence of fire, while 80% exhibited damage to the bark, all of which also showed damage to the termite nests. Of the 15 palms with no nests, 12 exhibited evidence of fire, but none exhibited damage to the bark. An analysis of the association between damage to nest and damage to bark is depicted in . There was a significant positive association between the damage to termite nests and damage to palm bark (t = 6.1, df = 68:67, P < .001), which suggests that the presence of the nests increases the extent of fire damage to the palm trunks. A resurvey of these palms (along Transect Line 2) in June 2017 following a second fire found that most of the trees (96%) were still alive despite the damage to their bark, however, some palms were seen to have suffered fatal damage presumably due to repeated burning (–h)).
Figure 3. Association between the size of termite nests (i.e. the maximum Feret diameter: 0–10, 10–20, 20–30 or >30 cm) and the extent of damage to Mauritia flexuosa stems (% damage to rhytidome and cortex) following a wildfire. The fitted line is derived from a generalised linear model with a quasi-poisson distribution fitted using a maximum likelihood function in Rstudio (version 1.2.1335). Only palms showing evidence of fire are included, palms with termite nest above 2.5 m are also excluded. Points are jittered for clarity, n = 69
Savanna palms are generally considered to be resilient to wildfires, but the effect of wildfire’s on savanna palms is understudied, and recurrent fires can have negative impacts [Citation27,Citation28]. Previous work in the ASESA has estimated the rate of death in adult palms to be 0.7% per year for areas not impacted by fires and 3.0% per year for fire-impacted areas [Citation5]. Given the prevalence of termite nests near the base of M. flexuosa palms in this habitat, their role in fire damage is likely to be a significant driver of mortality in this species over time. Termites are generally considered to have a positive influence in fire-prone areas because they clear away dead material, which reduces the impact of future fires. However, termites may have a less benign impact on palms. Long term studies should be undertaken to monitor the mortality rate of M. flexuosa adults subjected to repeated burning, especially when termite nests are present.
Acknowledgments
We wish to thank Edmund Charles for providing field guidance and useful anecdotal information. We would also like to thank Christopher Starr and Scott Zona for commenting on earlier drafts of this manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Van Der Hoek Y, SolasSA, Peñuela MC. The palm Mauritia flexuosa, a keystone plant resource on multiple fronts. Biodivers Conserv. 2019;28(3):539–551.
- Beard JS. The Savanna vegetation of northern tropical America. Ecol Monogr. 1953;23(2):149–215.
- Montoya E, Rull V, Stansell ND, et al. Forest–Savanna–morichal dynamics in relation to fire and human occupation in the Southern Gran Sabana (SE Venezuela) during the last millennia. Quatern Res. 2011;76(3):335–344. .
- Mittermeier JC, Zyskowski K, Stowe ES, et al. Avifauna of the Sipaliwini savanna (Suriname) with insights into its biogeographic affinities. Bull Peabody Mus Nat Hist. 2010;51(1):97–123.
- Arneaud LL, Farrell AD, Oatham MP. Marked reproductive plasticity in response to contrasting fire regimes in a neotropical palm. Trop Ecol. 2017;58(4):693–703.
- John-Bejai C, Farrell AD, Cooper FM, et al. Contrasting physiological responses to excess heat and irradiance in two tropical savanna sedges. AoB Plants. 2013;5:5.
- Hosein A, Narang DS, Rostant L, et al. The abundance of red-bellied macaws (Orthopsittaca manilata) and orange-winged parrots (Amazona amazonica) in relation to fruiting moriche palms (Mauritia flexuosa) at the Aripo Savannas, Trinidad. Revista Brasileira de Ornitologia-Braz J Ornithol. 2017;25(1):40–46.
- Federman S, Hyseni C, Clement W, et al. Habitat fragmentation and the genetic structure of the Amazonian palm Mauritia flexuosa Lf (Arecaceae) on the Island of Trinidad. Conserv Genet. 2013;15(2):355–362.
- Arneaud LL, Duncan EJ. Preliminary checklist of endomycorrhizal fungi associated with Mauritia flexuosa Lf (Arecaceae, Calamoideae) in Trinidad, WI. living world. J Trinidad Tobago Field Naturalists’ Club. 2019;(1):1–6.
- Jiménez JJ, Decaëns T, Lavelle P. Nutrient spatial variability in biogenic structures of nasutitermes (Termitinae; Isoptera) in a gallery forest of the Colombian ‘Llanos’. Soil Biol Biochem. 2006;38(5):1132–1138.
- Howard FW. Borers of palms. CABI Publishing, Wallingford, GB; 2001.
- Johansson B. Stay below water!-A strategy to avoid seed predators: seed survival and germination of Mauritia flexuosa in Southeastern Peru. University of Linköping, 581–583 Linköping, Sweden; 2009.
- Junk WJ. The central Amazon floodplain: ecology of a pulsing system. In: Martius C, editor. The termites. Springer-Verlag Berlin Heideberg; 1997:69–108.
- Arneaud LL. Do leaf-cutter ants affect Mauritia flexuosa population structure? PALMS. 2018;62(1):35–36.
- Rull V, Montoya E. Mauritia flexuosa palm swamp communities: natural or human-made? A palynological study of the Gran Sabana region (Northern South America) within a neotropical context. Quat Sci Rev. 2014;99(1):17–33.
- Hoffmann WA, Geiger EL, Gotsch SG, et al. Ecological thresholds at the savanna‐forest boundary: how plant traits, resources and fire govern the distribution of tropical biomes. Ecol Lett. 2012;15(7):759–768.
- Nghalipo E, Joubert D, Throop H, et al. The effect of fire history on soil nutrients and soil organic carbon in a semi-arid savanna woodland, central Namibia. Afr J Range Forage Sci. 2019;36(1):9–16.
- EMA (Environmental Management Authority). Managing together: a summary of the management plans for the Aripo Savannas environmentally sensitive area. Port of Spain: Ministry of Housing, Land and Marine Affairs; 2008.
- Bisramsingh A, Oatham M, editors. Fire in the Aripo Savannas environmentally sensitive area: causes and consequences 2011 25 November, 2010. Port of Spain, Trinidad and Tobago. St. Augustine, Trinidad and Tobago: The Department of Life Science, UWI; 2011.
- E.M.A (Evironmental Management Authority). Managing together: a summary of the management plans for the Aripo Savannas environmentally sensitive area. Port of Spain: Ministry of Housing, Land and Marine Affairs; 2008.
- Schwab SI. Floral and faunal composition, phenology, and fire in the Aripo Savannas scientific reserve, Trinidad, West Indies [Unpublished Master Thesis]. University of Wisconsin-Stevens Point; 1988.
- Beck CB. Periderm, rhytidome, and the nature of bark. An introduction to plant structure and development: plant anatomy for the twenty-first century. 2nd ed. Cambridge University Press, Cambridge, England, UK; 2010. p. 459.
- Tomlinson PB. The uniqueness of palms. Bot J Linn Soc. 2006;151(1):5–14.
- Hodel DR. Biology of palms and implications for management in the landscape. HortTechnology. 2009;19(4):676–681.
- Rosell JA, Olson ME, Anfodillo T, et al. Exploring the bark thickness–stem diameter relationship: clues from lianas, successive cambia, monocots and gymnosperms. New Phytol. 2017;215(2):569–581. .
- Arneaud LL. Sustainability measures needed to preserve ‘tree of life’- researcher. Kaieteur News; 2018. Available from: www.kaieteurnewsonline.com/2018/01/22/sustainability-measures-needed-to-preserve-tree-of-life-researcher/.
- Mandle L, Ticktin T, Zuidema PA. Resilience of palm populations to disturbance is determined by interactive effects of fire, herbivory and harvest. J Ecol. 2015;103(4):1032–1043.
- Montúfar R, Anthelme F, Pintaud J-C, et al. Disturbance and resilience in tropical American palm populations and communities. Bot Rev. 2011;77(4):426–461.