
Abstract
The complete chloroplast genome of Farinopsis salesoviana is reported and characterized in this study. The chloroplast genome of F. salesoviana was a circular form of 154,901 bp in length. The genome presented a typical quadripartite structure consisting of a pair of inverted repeats (IRa and IRb) of 25,991 bp separated by a large single copy (LSC) region of 84,847 bp and a small single copy (SSC) region of 18,072 bp. The genome contained a set of 129 genes, including 84 protein-coding genes, 37 tRNA genes, and eight rRNA genes. Phylogenetic analysis indicated that F. salesoviana was sister to species of Alchemilla in current sampling.
Farinopsis salesoviana (Steph.) Chrtek et Soják (synonyms: Comarum salesovianum [Steph.] Asch. et Gr., Potentilla salesoviana Steph.; Rosaceae Juss.: Potentilleae Sweet), is native to Pakistan and Asia-Temperate according to Plants of the World online maintained by the Royal Botanic Gardens, Kew, UK (http://powo.science.kew.org). Farinopsis salesoviana has been used as traditional medicine (Ligaa et al. Citation2006; Khan et al. Citation2011). Its aerial parts have antibacterially active phenolic lipid derivatives (Odontuya et al. Citation2015). Herein, the complete chloroplast (cp) genome of F. salesoviana was reported, which is useful for further studies on its taxonomy, population genetics, and medical use.
Fresh leaves of F. salesoviana were collected from Qilian County, Qinghai Province, China. Voucher specimen (no. Li QQ ML1) was deposited in the herbarium of Inner Mongolia Normal University (NMTC). Total genomic DNA was extracted using the method of Doyle and Doyle (Citation1987). Short-insert library (insert size, 300 bp) was prepared and then sequenced using an Illumina HiSeq 2000 Sequencing System in Novogene (Nanjing, China). Illumina paired-end sequencing generated 36,331,508-bp raw reads after adapters removed. The raw reads were used to assemble the cp genome in NOVOPlasty (Dierckxsens et al. Citation2017) with ribulose-1, 5-bisphosphate carboxylase/oxygenase (rbcL) gene from F. salesoviana (GenBank accession no. KY420034) as the seed. Chloroplast genome annotation was performed using GeSeq (Tillich et al. Citation2017) with cp genomes of Fragaria chiloensis (L.) Mill. (NC_019601) and Dasiphora fruticosa (L.) Rydb. (NC_036423) from GenBank as references. Draft annotation generated using GeSeq was imported into Geneious Prime (Kearse et al. Citation2012) for further manual adjustment. The annotated complete cp genome of F. salesoviana was deposited in GenBank with accession no. MT017928. The cp genome of F. salesoviana was a circular form with a size of 154,901 bp in length and had a typical quadripartite structure consisting of a pair of inverted repeats (IRa and IRb: 25,991 bp) separated by a large single-copy (LSC: 84,847 bp) and a small single-copy (SSC: 18,072 bp) regions. The overall GC content was 37.1%. The cp genome contained a set of 129 genes, including 84 protein-coding genes, 37 tRNA genes, and eight rRNA genes.
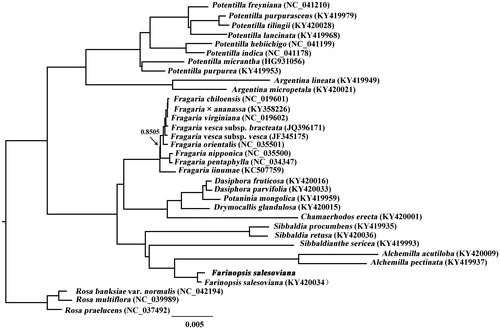
To ascertain the phylogenetic position of F. salesoviana, the cp genome sequences of 33 Potentilleae and Rosa taxa were downloaded from GenBank. All the cp genome sequences were aligned with MAFFT version 7.450 (Katoh and Standley Citation2013) and trimmed properly using trimAL version 1.4 (Capella-Gutiérrez et al. Citation2009). The GTR + I + G model was selected as a best-fit model using PartitionFinder2 (Lanfear et al. Citation2016). Bayesian inference (BI) was conducted using the MrBayes version 3.2.2 (Ronquist et al. Citation2012). Four independent Markov chain Monte Carlo (MCMC) algorithm were run for 6,000,000 generations, starting from random trees and sampling trees every 100 generations, with the first 25% discarded as burn-in. A 50% majority-rule consensus tree was finally produced. Phylogenetic tree showed that F. salesoviana was sister to species of Alchemilla L. in current sampling (), which is congruent with Zhang et al. (Citation2017).
Figure 1. Phylogenetic tree resulting from a Bayesian analysis of the cp genome sequences from 31 Potentilleae taxa plus three Rosa taxa as outgroups. Branch lengths correspond to the genetic distances (substitutions per site). Value along branch represents Bayesian posterior probability (only PP < 1.0 is shown).
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
Related Research Data
References
- Capella-Gutiérrez S, Silla-Martínez JM, Gabaldón T, 2009. trimAl: a tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics. 25(15):1972–1973.
- Dierckxsens N, Mardulyn P, Smits G. 2017. NOVOPlasty: de novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 45(4):e18.
- Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small amounts of fresh leaf tissue. Phytochem Bull. 19 (1):11–15.
- Katoh K, Standley DM. 2013. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 30(4):772–780.
- Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, et al. 2012. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 28(12):1647–1649.
- Khan B, Abdukadir A, Qureshi R, Mustafa G. 2011. Medicinal uses of plants by the inhabitants of Khunjerab National Park, Gilgit, Pakistan. Pak J Bot. 43(5):2301–2310.
- Lanfear R, Frandsen PB, Wright AM, Senfeld T, Calcott B. 2016. PartitionFinder 2: new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol Biol Evol. 34(3):772–773.
- Ligaa U, Davaasuren B, Ninjin N. 2006. Medicinal plants of Mongolia used in western and eastern medicine. Ulaanbaatar (Mongolia): JKL Printing, p. 218–219.
- Odontuya G, Banzragchgarav O, Murata T, Batkhuu J, Sasaki K, Yoshizaki F. 2015. Antibacterially active phenolic lipid derivatives from Comarum salesovianum (Steph.) Aschers. et Gr. Phytochem Lett. 13:360–364.
- Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck J P. 2012. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 61(3):539–542.
- Tillich M, Lehwark P, Pellizzer T, Ulbricht-Jones ES, Fischer A, Bock R, Greiner S. 2017. GeSeq–versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 45(W1):W6–W11.
- Zhang SD, Jin JJ, Chen SY, Chase MW, Soltis DE, Li HT, Yang JB, Li DZ, Yi TS. 2017. Diversification of Rosaceae since the Late Cretaceous based on plastid phylogenomics. New Phytol. 214(3):1355–1367.