
Abstract
Song plays a vital role in communication in songbirds, primarily for territorial defense and mate attraction. Larger animals, having larger vocal organs, produce low sound frequencies more efficiently. Accordingly, the frequency of vocalizations is often negatively correlated with body size across species, and also among individuals of many species, including several non-songbirds. However, little is known about whether song frequency reveals information about body size among males. We tested for the predicted positive relationship between body size and song traits in male dusky warblers (Phylloscopus fuscatus). In many animal species, some vocalizations are difficult to produce and can therefore indicate quality of signalers. Male dusky warblers produce songs with trilled sequences that are limited in their timing and frequency structure by physiological constraints on vocal performance. We investigated whether there was a relationship between body size and trilled vocalizations in the dusky warbler. We recorded songs of free-living male dusky warblers at dawn, and captured birds with mist nets for morphometric measurements (tarsus length, as an indicator of body size). We conducted correlation analyses between tarsus length and a composite measure of songs. In dusky warblers, body size was not significantly related to the average frequency of their overall songs, low frequency, bandwidth frequency, average note number of songs, or average song duration. In contrast, the maximum frequency of song was found to correlate positively with tarsus length. For trilled vocalizations, we found most trill traits were not significantly related to body size, and only the maximum frequency of trills was positively related to body size. Additionally, the maximal value of frequency bandwidth decreased with increasing trill rates. Overall, these results suggest that in male dusky warblers, the maximum frequency of the entire song is a reliable indicator of body size.
Introduction
Bird song is one of the most widely used means of communication in both oscine and suboscine birds. Songbirds use acoustic signals to elicit courtship and defend territories from rival conspecifics (Catchpole Slater 2008). Therefore, birdsong, as a sexually selected multidimensional signal, can reveal singer information to some extent. It is well known that advertising songs can indicate body size (Hall et al. Citation2013; Price & Crawford Citation2013). In many studies, all song characteristics reveal signaler quality through traits such as repertoire size (Soma & Garamszegi Citation2011; Hesler et al. Citation2012; Kagawa & Soma Citation2013), song timing (Poesel et al. Citation2006), song amplitude (Brumm Citation2009; Schuchmann & Siemers Citation2010), song frequency, song bandwidth or song rate (Potvin Citation2013; Linhart & Fuchs Citation2015; Mason & Burns Citation2015).
However, in other studies, many song characteristics failed to predict the body size of songbirds (Brumm Citation2009; Geberzahn et al. Citation2009; Miyashita et al. Citation2016). In these studies, song bandwidths, song rate, average frequency, song amplitude and repertoire size of the total song showed no relationship to body size. Therefore, females are unlikely to assess the size of male birds solely from individual parts of the song, but rather use the total song in their assessment of males. Perhaps some fine-scaled variation in sound production can reflect body size.
Songs of dusky warblers have been studied extensively. Males usually reduce the intensity of singing once they are mated with a female. Mated dusky warblers sing as much before as they do after pairing (Forstmeier & Balsby Citation2002). The reason that mated dusky warblers continue to sing is to assure within-pair paternity and gain the opportunity for extra-pair copulations. In another study fine-scaled variation in sound amplitude was correlated with both male success in obtaining extra-pair copulations and male longevity (Forstmeier et al. Citation2002). This indicates that females judge male quality by subtle differences in their performance during the production of song notes, rather than by the quantity or versatility of the song. However, the question remains whether songs themselves or fine-scaled variation within songs correlate with body size.
The dusky warbler produces songs with trilled sequences (Forstmeier & Balsby Citation2002; Ivanitskii et al. Citation2012). Ivanitskii et al. (Citation2012) found that 90% of the songs of dusky warblers contained only one trill. In dusky warblers, trilled sequences can be a part of a song, or a complete song in themselves, and they are limited in their timing and frequency structure by the physical constraints of vocal performance. We tested whether trill traits are signals of body size in male dusky warbler.
Podos (Citation2001) suggested that trill rate and frequency bandwidth could be used by birds in mate recognition. Sexually selected signals are reliable in revealing singer quality. These factors led us to focus our attention on trilled sequences within dusky warbler songs. In this study, we first investigated the relationship between trill rate and frequency bandwidth. This was done to assess whether this relationship in dusky warblers was similar to that observed in other species. Second, we investigated the hypothesis that trill traits are related to body size, and that larger males within a species sing songs of higher quality. Higher quality songs have larger bandwidth and faster pitch. Finally, we tested whether trill traits indicate body size.
Materials and methods
Study species
The dusky warbler is a single-brooded, long-distance migratory passerine with a cryptic, sexually monomorphic plumage lacking any obvious visual sexual decoration (Forstmeier et al. Citation2002). We conducted an investigation of a population of individual male dusky warblers in Saihanba Forest Farm, Hebei Province (42°02′–42°36′N, 116°51′–117°39′E) during July 2015. The dusky warbler lives at high population densities, in bushlands of varying coverage and height, on the banks of the Luan River. During the breeding season males do not guard fertile females, but singing does intensify when their partners are fertile (Forstmeier & Balsby Citation2002). They spend most of the daytime singing. Dusky warblers sing two distinct kinds of song: S- and V-song. There are differences between S- and V-song: (1) V-song is, on average, produced at a higher rate per minute compared to S-song; (2) in V-song, pauses between strophes may contain series of short calls, and these calls were present in 50% of 107 V-song recordings, but were never heard in S-song; (3) V-song strophes, on average, are longer than S-song strophes, and contain more and a more variable number of syllables; (4) V-song regularly shows syllable-type switches within strophes, that do not occur in S-song; and (5) V-song comprises large individual repertoires (Forstmeier & Balsby Citation2002). Because different song categories have different structures and functions, it seems important to account for song category in the analysis. For this reason, we only analyzed V-songs.
Song analyses
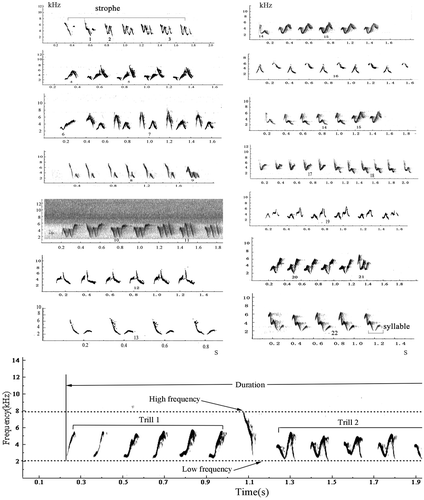
We recorded male songs between 5:00 a.m. and 10:00 a.m., at a distance not exceeding 10 m around the singing bird in good weather conditions (no wind, no rain). We used a TASCAM DR-680 recorder and a Sennheiser ME67 microphone with a Rycote Softie windshield. Songs were recorded at a 44.1 kHz sampling rate and 16-bit depth. Spectrograms were generated using Raven Pro sound analysis software (version 1.4; Cornell Laboratory of Ornithology, Ithaca, NY). Within Raven, we used a Hann spectrogram window, a DFT size of 512 samples, a hop size of 3.4 ms, a sampling frequency of 44.1 kHz, and a time resolution of 11.6 ms. Measurements were taken by visually cross-referencing the spectrogram and waveform displays to determine when vocalizations began and ended. Raw measurements included the start and end time as well as the low, high and maximum frequency of each trill and song. Maximum frequency was the frequency at which maximum power occurs in the song. The number of the notes in a trill was visually counted in Raven (). For analysis, (1) we defined a trill as a series of notes or note groups repeated in succession, and trill rate as the number of notes per second in a trill; (2) frequency bandwidth was calculated as the difference between the upper and lower frequency, and duration was the difference between start time and end time (); and (3) trill quality was calculated as the product of trill bandwidth and trill rate.
Figure 1. Spectrograms of 10 continuous strophes of variable song-type (V-song) in dusky warblers, and the parameters measured in this study. Numbers reflect the classification of different syllable types.
Body size measurements
To investigate the relationship between body size and song pitch, 33 males were recorded and measured, between 15 and 20 July 2015. We recorded males for 10 minutes and then lured them into a mist net with playback. We measured tarsus length (± 0.1 mm) 3 times at each capture, and used the average length as an indicator of body size. We used tarsus length as our measure of body size for the following three reasons: (1) it is observed to predict male quality and female choice in some birds (Kempenaers et al. Citation1992; Møller & Ninni Citation1998; Hall et al. Citation2013); (2) it has an observed relationship with call frequency (Price et al. Citation2006; Brumm Citation2009; Geberzahn et al. Citation2009; Linhart & Fuchs Citation2015); and (3) it is constant in adults.
Statistical analyses
We used quantile regression to explore how changes in syllable repetition rate affected the ninetieth percentile of the frequency bandwidth distribution. We used quantile regression rather than standard linear regression because we were mainly interested in the effects of song features at the tail of the frequency distributions. This analysis was conducted using the quantreg package in R software. For this, 1313 trilled sequences from 33 male dusky warblers were analyzed, with an average of 40 trills per individual. We calculated average values of all songs for each individual, and used Pearson’s correlation to test for a relationship between tarsus length and song traits. Multiple comparisons were required to consider the probability of false positives, so we applied Benjamini–Hochberg procedure to control the false discovery error rate and to correct P values.
Results
Relationship between trill rate and frequency bandwidth
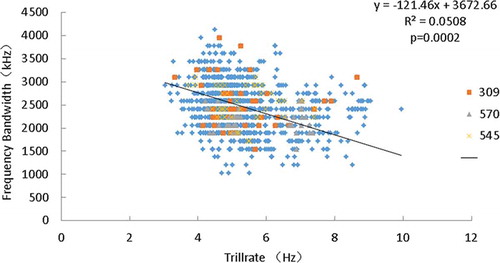
Quantile regression analysis showed that frequency bandwidth correlated negatively with trill rate (). Quantile regression for the sample was statistically significant at p < 0.05, and had a negative slope (y = −121.46x + 3672.66, R2 = 0.05, P = 0.00020).
Figure 2. Relationship between trill rate and frequency bandwidth in male dusky warblers. In this figure, the three different types represent songs of three different individuals.
Sound traits and body size
We conducted correlation analyses between tarsus length and a composite measure of songs (). Tarsus length showed no significant relationship with low frequency of songs (Pearson correlation: r = −0.188, P = 0.30, P critical = 0.033), bandwidth frequency of songs (Pearson correlation: r = −0.164, P = 0.36, P critical = 0.038), average note number of songs (Pearson correlation: r = 0.245, P = 0.17, P critical = 0.025), and average song duration (Pearson correlation: r = 0.206, P = 0.25, P critical = 0.029). The only significant relationship of tarsus length was with song maximum frequency, which showed a positive correlation (Pearson correlation: r = 0.524, P = 0.002, P critical = 0.004).
Table I. Relationships between average vocalization traits and tarsus length in male dusky warblers. Parameters with double asterisks were significant after the Benjamini–Hochberg adjustment.
Data in show that tarsus length also was positively related to average maximum frequency of trills (Pearson correlation: r = 0.471, P = 0.006, P critical = 0.008). In this study, for all other trilled vocalization features, we found no significant correlations with tarsus length (Pearson correlation test; low frequency of trills: r =−0.411, P = 0.017, P critical = 0.016; average note number of trills: r = 0.415, P = 0.016, P critical = 0.012; average duration of trills: r = 0.386, P = 0.027, P critical = 0.021; trill rate: r = −0.044, P = 0.81; frequency bandwidth: r = −0.117, P = 0.52; trill quality: r = −0.107, P = 0.56).
Discussion
Maximum song frequency was the best predictor of body size, not only in the trill part of the song, but also in the measurement of the entire song. Therefore, we conclude that maximum frequency of whole songs was the best indicator of body size in male warblers. However, an inverse relationship between body size and vocalization pitch has been described in a number of species (Smith & Harper Citation2004; Koetz et al. Citation2007; Potvin Citation2013; Linhart & Fuchs Citation2015; Mason & Burns Citation2015). In these studies, low frequency was the best predictor of body size, which is contrary to our results. The maximum frequency of entire songs was the best predictor of body size in male dusky warblers, likely because the maximum frequency of songs was closer to the upper limit of its potential frequency range. Therefore, this species may use maximum frequency as a relevant sexual signal, and female dusky warblers may prefer males with larger body size that sing at a higher maximum frequency.
Forstmeier & Balsby (Citation2002) revealed that males sing to guard territory and to announce their quality for copulation. Male dusky warbler songs were classified into two distinct types: S-song and V-song. S-song was a territorial signal to mark claimed territory. V-song appeared to attract females for copulation. However, it was not known whether V-song quality was correlated with body size, or how song was correlated with male body size. In another study, Forstmeier et al. (Citation2002) showed that a male’s ability to maintain a high sound amplitude during singing could reflect individual quality, and that amplitude in songs was correlated with male success in obtaining extra-pair copulations. However, it was not known whether the size of extra-pair fathers was bigger than the male they cuckolded.
For the two song types, we studied only the V-song, because V-song or advertising song was used to attract females for copulation. Therefore, this song type is more likely to be correlated with body size. Ivanitskii et al. (Citation2012) made a detailed comparison of syntax in the advertising song of dusky and Radde’s (Ph. schwarzi) warblers, and he found that in Southern Siberia 90% of songs produced by male dusky warblers contained only one trill. That was also the case in our study, because most of the songs of this species contained only one trill, and the trill could be a single song. However, many songs consisted of two parts: a trilled part and a non-trilled part. We expect that trill parts (perhaps difficult to produce) are more likely to reflect body size. Here we have shown that the trilled part was closely related to body size. For the parameters of advertising song of dusky warblers, our study differed from that of Ivanitskii et al. (Citation2012). In our study, advertising songs had a duration of 0.13–3.30 s and a frequency range of 1.22–9.53 kHz, while in the study of Ivanitskii et al. (Citation2012) advertising songs had a duration of 0.7–1.5 s and a frequency range of 1.2–7.6 kHz. Differences in sample size may lead to the difference in song duration and frequency range between the two studies. Ivanitskii et al. (Citation2012) analyzed 24 songs of eight males, while we analyzed 1313 songs of 33 males. The range of time characteristics and frequency characteristics was larger than 11.6 ms and 44.1 Hz. Thus we consider that spectrograph measurement error will have little impact on our conclusions.
Frequency bandwidth was negatively correlated with trill rate. These findings are in agreement with previous studies. There are physiological limits during trilled vocalization production. For example, in hand clapping, hands can be clapped either quickly or loudly, but beyond a certain point one may not produce claps both quickly and loudly. The ability to perform physically challenging songs predicts a male’s quality. Perhaps in the songs of male dusky warblers, trilled parts are more difficult to produce than other variables that are closely related to body size. We found the maximum frequency of trills was positively correlated with body size, and this means that maximum frequency of trills could also be a reliable indicator of body size.
Trill rate, bandwidth and their product were not correlated with tarsus length. This result seems to be contrary to the “trill performance” hypothesis (at least if size reflects quality). One possible explanation is that the sample size was insufficient for detecting a relationship. Perhaps upon enlarging the sample size, there will be an opposite result. Another explanation may be that trill characteristics were used in territorial defense. Thus, advertising songs would not show a relationship with body size, while in territorial songs trill characteristics may be related to body size. Additionally, perhaps trill characteristics reflect body mass or male age. For example, Sprau et al. (Citation2013) found that trill rate, bandwidth, and their product did not significantly predict body size, but predicted age in male nightingales Luscinia megarhynchos. Some studies discovered that frequency bandwidth or trill rate was negatively related to body mass (Tubaro & Mahler Citation1998; Michael et al. Citation2012). Lastly, bird song is comprised of multiple components, each of which can be limited by specific constraints. Although body size has been reported to be an important factor in songbird contests, it is still unknown whether tarsus length – used as an indicator of body size in this study – is an appropriate indicator of such constraints. Different studies have used different indicators to describe body size, such as weight, wing length, beak size, or a combination of these body size measures (Searcy Citation1979; Koivula et al. Citation1993; Jonart et al. Citation2007). We hypothesize that different methods of measurement may lead to different results. Additionally, large individuals do not ensure high song quality. Songs of the same individual are not concentrated in one area, and this also suggests that large individuals only signal that they have the ability to sing high quality songs, and not that they will sing songs of high quality all the time.
Ethical standards
Our research was approved by Hebei provincial wildlife protection, which is the highest wildlife administrative institution in Hebei province. For our study, we captured birds in mist nets and then recorded morphometric measurements (tarsus length). No birds were injured and, after measuring body traits, birds were released as soon as possible. Our study did not involve anesthesia, euthanasia or any kind of animal sacrifice. The safety and well-being of the birds was the primary concern at all times.
Acknowledgements
We thank our field assistants for their work on the project, and Dr. Canwei Xia for comments on the manuscript.
Additional information
Notes on contributors
J. P. Liu
The experiments were conceived and designed by Liu Jianping and Hou Jianhua, and performed by Liu Jianping, Zhang Zhenqun, Gao Lijie, Gu Dehai, Ma Laikun and Li Junjie. The data were analyzed by Liu Jianping, Zhang Zhenqun, Gu Dehai, Ma Laikun and Wang Jiaojiao. Liu Jianping wrote the manuscript.
L. K. Ma
The experiments were conceived and designed by Liu Jianping and Hou Jianhua, and performed by Liu Jianping, Zhang Zhenqun, Gao Lijie, Gu Dehai, Ma Laikun and Li Junjie. The data were analyzed by Liu Jianping, Zhang Zhenqun, Gu Dehai, Ma Laikun and Wang Jiaojiao. Liu Jianping wrote the manuscript.
Z. Q. Zhang
The experiments were conceived and designed by Liu Jianping and Hou Jianhua, and performed by Liu Jianping, Zhang Zhenqun, Gao Lijie, Gu Dehai, Ma Laikun and Li Junjie. The data were analyzed by Liu Jianping, Zhang Zhenqun, Gu Dehai, Ma Laikun and Wang Jiaojiao. Liu Jianping wrote the manuscript.
D. H. Gu
The experiments were conceived and designed by Liu Jianping and Hou Jianhua, and performed by Liu Jianping, Zhang Zhenqun, Gao Lijie, Gu Dehai, Ma Laikun and Li Junjie. The data were analyzed by Liu Jianping, Zhang Zhenqun, Gu Dehai, Ma Laikun and Wang Jiaojiao. Liu Jianping wrote the manuscript.
J. J. Wang
The experiments were conceived and designed by Liu Jianping and Hou Jianhua, and performed by Liu Jianping, Zhang Zhenqun, Gao Lijie, Gu Dehai, Ma Laikun and Li Junjie. The data were analyzed by Liu Jianping, Zhang Zhenqun, Gu Dehai, Ma Laikun and Wang Jiaojiao. Liu Jianping wrote the manuscript.
J. J. Li
The experiments were conceived and designed by Liu Jianping and Hou Jianhua, and performed by Liu Jianping, Zhang Zhenqun, Gao Lijie, Gu Dehai, Ma Laikun and Li Junjie. The data were analyzed by Liu Jianping, Zhang Zhenqun, Gu Dehai, Ma Laikun and Wang Jiaojiao. Liu Jianping wrote the manuscript.
L. J. Gao
The experiments were conceived and designed by Liu Jianping and Hou Jianhua, and performed by Liu Jianping, Zhang Zhenqun, Gao Lijie, Gu Dehai, Ma Laikun and Li Junjie. The data were analyzed by Liu Jianping, Zhang Zhenqun, Gu Dehai, Ma Laikun and Wang Jiaojiao. Liu Jianping wrote the manuscript.
J. H. Hou
The experiments were conceived and designed by Liu Jianping and Hou Jianhua, and performed by Liu Jianping, Zhang Zhenqun, Gao Lijie, Gu Dehai, Ma Laikun and Li Junjie. The data were analyzed by Liu Jianping, Zhang Zhenqun, Gu Dehai, Ma Laikun and Wang Jiaojiao. Liu Jianping wrote the manuscript.
References
- Brumm H. 2009. Song amplitude and body size in birds. Behavioral Ecology and Sociobiology 63:1157–1165. DOI:10.1007/s00265-009-0743-4.
- Catchpole CK, Slater PJB. 2008. Bird song: biological themes and variations. Cambridge, UK: Cambridge University Press.
- Forstmeier W, Balsby T. 2002. Why mated dusky warblers sing so much: territory guarding and male quality announcement. Behaviour 139:89–111. DOI:10.1163/15685390252902300.
- Forstmeier W, Kempenaers B, Meyer A, Leisler B. 2002. A novel song parameter correlates with extra-pair paternity and reflects male longevity. Proceedings of the Royal Society B-Biological Sciences 269:1479–1485. DOI:10.1098/rspb.2002.2039.
- Geberzahn N, Goymann W, Muck C, Cate CT. 2009. Females alter their song when challenged in a sex-role reversed bird species. Behavioral Ecology and Sociobiology 64:193–204. DOI:10.1007/s00265-009-0836-0.
- Hall ML, Kingma SA, Peters A, Johnson N. 2013. Male songbird indicates body size with low-pitched advertising songs. PLoSOne 8:5671. DOI:10.1371/journal.pone.0056717.
- Hesler N, Mundry R, Sacher T, Coppack T, Bairlein F, Dabelsteen T. 2012. Song repertoire size correlates with measures of body size in Eurasian blackbirds. Behaviour 149:645–665. DOI:10.1163/156853912X649920.
- Ivanitskii VV, Marova IM, Malykh IM. 2012. Between order and chaos: Contrasting syntax in the advertising song of Dusky (Phylloscopus fuscatus) and Radde’s (Ph. schwarzi) Warblers. Journal of Ornithology 153:337–346. DOI:10.1007/s10336-011-0748-4.
- Jonart LM, Hill GE, Badyaev AV. 2007. Fighting ability and motivation: Determinants of dominance and contest strategies in females of a passerine bird. Animal Behaviour 74:1675–1681. DOI:10.1016/j.anbehav.2007.03.012.
- Kagawa H, Soma M. 2013. Song performance and elaboration as potential indicators of male quality in Java sparrows. Behavioural Processes 99:138–144. DOI:10.1016/j.beproc.2013.07.012.
- Kempenaers B, Verheyen GR, Broeck MVD, Burke T, Broeckhoven CV, Dhondt A. 1992. Extra-pair paternity results from female preference for high-quality males in the blue tit. Nature 357:494–496. DOI:10.1038/357494a0.
- Koetz AH, Westcott DA, Congdon BC. 2007. Geographical variation in song frequency and structure: The effects of vicariant isolation, habitat type and body size. Animal Behaviour 74:1573–1583. DOI:10.1016/j.anbehav.2007.03.022.
- Koivula K, Lahti K, Orell M, Rytkönen S. 1993. Prior residency as a key determinant of social dominance in the willow tit (Parus montanus). Behavioral Ecology and Sociobiology 33:283–287. DOI:10.1007/BF02027126.
- Linhart P, Fuchs R. 2015. Song pitch indicates body size and correlates with males’ response to playback in a songbird. Animal Behaviour 103:91–98. DOI:10.1016/j.anbehav.2015.01.038.
- Mason NA, Burns KJ. 2015. The effect of habitat and body size on the evolution of vocal displays in Thraupidae (tanagers), the largest family of songbirds. Biological Journal of the Linnean Society 114:538–551. DOI:10.1111/bij.12455.
- Michael W, Sarah K, Silke K. 2012. Buzzwords in females ears? The use of buzz songs in the communication of Nightingales (Luscinia megarhynchos). PLoS One 7:e45057. DOI:10.1371/journal.pone.0045057.
- Miyashita A, Kizaki H, Sekimizu K, Kaito C, Fine ML. 2016. No effect of body size on the frequency of calling and courtship song in the two-spotted Cricket, Gryllus bimaculatus. PLoS One 11:e0146999. DOI:10.1371/journal.pone.0146999
- Møller AP, Ninni P. 1998. Sperm competition and sexual selection: A meta-analysis of paternity studies of birds. Behavioral Ecology and Sociobiology 43:345–358. DOI:10.1007/s002650050501.
- Podos J. 2001. Correlated evolution of morphology and vocal signal structure in Darwin’s finches. Nature 409:185–188. DOI:10.1038/35051570.
- Poesel A, Kunc HP, Foerster K, Johnsen A, Kempenaers B. 2006. Early birds are sexy: Male age, dawn song and extrapair paternity in blue tits Cyanistes (formerly Parus) caeruleus. Animal Behaviour 72:531–538. DOI:10.1016/j.anbehav.2005.10.022.
- Potvin DA. 2013. Larger body size on islands affects silvereye Zosterops lateralis song and call frequency. Journal of Avian Biology 44:221–225. DOI:10.1111/j.1600-048X.2012.05820.x.
- Price JJ, Crawford CL. 2013. Use and characteristics of two singing modes in pine warblers. The Wilson Journal of Ornithology 125:552–561. DOI:10.1676/13-006.1.
- Price JJ, Earnshaw SM, Webster MS. 2006. Montezuma oropendolas modify a component of song constrained by body size during vocal contests. Animal Behaviour 71:799–807. DOI:10.1016/j.anbehav.2005.05.025.
- Schuchmann M, Siemers BM, Somers M. 2010. Variability in echolocation call intensity in a community of horseshoe bats: A role for resource partitioning or communication? PLoS One 5:e12842. DOI:10.1371/journal.pone.0012842.
- Searcy WA. 1979. Morphological correlates of dominance in captive male red winged blackbirds. The Condor 81:417–420. DOI:10.2307/1366971.
- Smith M, Harper D. 2004. Animal signals. New York, NY: Oxford University Press.
- Soma M, Garamszegi LZ. 2011. Rethinking birdsong evolution: Meta-analysis of the relationship between song complexity and reproductive success. Behavioral Ecology 22:363–371. DOI:10.1093/beheco/arq219.
- Sprau P, Roth T, Amrhein V, Naguib M. 2013. The predictive value of trill performance in a large repertoire songbird, the nightingale Luscinia megarhynchos. Journal of Avian Biology 44:567–574. DOI:10.1111/jav.2013.44.issue-6.
- Tubaro PL, Mahler B. 1998. Acoustic frequencies and body mass in New World doves. The Condor 100:54–61. DOI:10.2307/1369896.