
Abstract
In Gansbaai (South Africa), at Dyer Island Nature Reserve, a large White shark population is present and can be observed due to the support of local ecotourism operators authorised to reach the field observation sites. Between 2009 and 2019, it was possible to create a database including information about each individual observed. In total, 423 white sharks were sighted during 462 direct observation hours from the boat, that included 220 hours from the diving “cage”. The mean sighting rate was 0.91 (range 0.18–1.53) sharks per hour and sighting rates dramatically declined in the last three years of the study period. Ninety-nine unique Photo-Ids of the dorsal fin were collected and only five re-sightings occurred, which indicate a transient behaviour for the Gansbaai White shark population. The sex ratio showed that females were always prevalent over males throughout the duration of the observations: the ratios were 1:2.2:0.8 for males, females, and unsexed sharks, respectively, and showed the prevalence of immature female individuals (immature: 51 males, 201 females, and 40 unsexed; adults: 49 males, 14 females, and 1 unsexed; undefined maturity: 5 males, 19 females, and 43 unsexed sharks). The predominance of immatures only applies to the females; there were as many immature males (51) as mature (49). The total length for all the individuals was between 150 cm and 500 cm (mean 308 cm, n = 423) with few young-of-the-year and adults recorded, indicating that Gansbaai Area is not a nursery area nor an adult aggregation site, but a seasonal feeding ground. The interannual sighting trend showed a consistent long-term increasing peak (ca. 4–5 years) and this could confirm that, in Gansbaai, the White shark frequency is not affected by ecotourism but, since 2017, a consistent loss of sightings was also due to recorded transient killer whales’ unusual fatal attacks.
Introduction
The White shark Carcharodon carcharias (Linnaeus 1758) is a protected species in many countries such as United States, South Africa, New Zealand, Australia, and Mexico (Fergusson et al. Citation2009) and this species has been categorised as Vulnerable since 1996 (Fergusson et al. Citation2009). This shark is relatively abundant in only the following eight “Great White hot spots” (Bass Citation1975; Ferreira & Ferreira Citation1996; Martin Citation2003; Becerril-Garcìa et al. Citation2020; Leone et al. Citation2020): central California (especially the Farallon Islands and Año Nuevo Island), central Chile (from Punta Angamos to Punta Lavapie), New England, the Mediterranean Sea, South Africa (especially the Indian Ocean side of the Western Cape and the Southern Cape regions), southern Australia, New Zealand, and Japan (from Sendai Bay to Kumano Bay). In South Africa, overall estimates by Cliff et al. (Citation1996) were 1279 sharks (95% confidence limits, 839–1843) from Richards Bay region in Kwazulu-Natal to Struis Bay in Western Cape; Ferreira and Ferreira (Citation1996) recorded 255 sharks in False Bay, Gansbaai, Struis Bay, Mossel Bay, and Algoa Bay; Towner et al. (Citation2013a) in Gansbaai identified 532 unique individuals and estimated the super-population size of 908; Andreotti et al. (Citation2016) estimated the South African super population at 438 sharks, but one year later Irion et al. (Citation2017) stated that, due to the complex stock structure of White sharks and the model assumptions made by Andreotti et al. (Citation2016), the conclusions drawn could not be supported by their methods and data. In any case, even if the estimates of the Gansbaai and South African population are currently being debated in the scientific literature, an important white shark population is present in the coastal South African area. Gansbaai is a world-recognised white shark aggregation site and it is the only location in the world where cage-diving trips operate daily, weather permitting (Towner Citation2012). The estimation of demographic parameters is essential in assessing long-term changes in vulnerable populations and determining the causes of population decline (Lee et al. Citation2014). It is thus important to understand the behavioural dynamics and turnover of individuals, sexes, and life-history stages within populations over time if shark populations are to be effectively managed (Hewitt et al. Citation2017). Photo identification is a long-term individual identification methodology for white sharks (Anderson et al. Citation2010, Citation2011) and a non-invasive method of mark and recapture. It provides information on white shark population size (Chapple et al. Citation2011; Towner et al. Citation2013a; Hewitt et al. Citation2017), sexual composition (Nasby-Lucas & Domeier Citation2012; Hewitt et al. Citation2017), longevity (Anderson et al. Citation2011), site fidelity (Bonfil et al. Citation2005; Anderson et al. Citation2011; Nasby-Lucas & Domeier Citation2012; Hewitt et al. Citation2017), and migratory behaviour (Bonfil et al. Citation2005). Photo identification must be very detailed, and the database must include data such as sex, length, parasites, and scars on the body to avoid potential mistakes (Towner et al. Citation2013a; Micarelli et al. Citation2015). White sharks frequently prey on Cape fur seals (Arctocephalus pusillus pusillus) at rookeries off southern Africa and particularly during autumn and winter, when juvenile seals leave to forage offshore for the first time, making them vulnerable to predation by white sharks (Martin et al. Citation2005; Hammerschlag et al. Citation2006; Kirkman et al. Citation2006; Towner et al. Citation2013b). False Bay, Gansbaai and Mossel Bay are white shark hotspots because the white sharks predate on the resident seals. Concerning shark population demographic data, Gansbaai seems to host a white shark population similar in size to that of Mossel Bay (SA), but in Mossel Bay a greater proportion of sub-adults and only occasional adults are present (Ferreira & Ferreira Citation1996; Ryklief et al. Citation2014). In Gansbaai, white sharks are predominantly sub-adults and adults that primarily feed on marine mammals (Estrada et al. Citation2006; Towner et al. Citation2013a). The greatest proportion of sub-adults and potentially mature sharks occurs in False Bay, South Africa (Kock & Johnson Citation2006). The main goal of this paper was to contribute to defining the role of Gansbaai area in the dynamics of South African white shark populations, investigating: A) inter-annual sighting trends, B) the sex-ratio, and C) the size structure and life-history composition.
Materials and methods
Study area
Observations and data collection were performed in the Nature Reserve of Dyer Island, located 7.5 km off the coast of Gansbaai (34° 41’ S; 19° 24’ E), South Africa. The Reserve includes Dyer Island and Geyser Rock (): the first is a low-profile island ca. 1.5 km long and 0.5 km wide and it is characterised by the presence of different sea bird colonies; the second is ca. 0.5 km long and 180 m wide, and it hosts a colony of Cape fur seals Arctocephalus pusillus pusillus (Schreber 1775). The reserve is located in an area called Agulhas Bioregion, which is the meeting point between the Benguela Current, which is the eastern boundary current of the subtropical vortex located in the south Atlantic Ocean and the Agulhas Current, which is the current forming the western limit of the Indian Ocean. In summer, intensified south-easterly trade winds result in upwelling, causing cold waters of Benguela origin to enter the bay (Jury Citation1985). Upwelling along the coast results in high biological productivity, which in turn supports large fish stocks, including pilchard, anchovy, and hake (Griffiths et al. Citation2010).
Figure 1. Gansbaai, Dyer Island, and Geyser Rock Nature Reserve
Data collection
Eleven scientific expeditions were conducted in the study area in all years between 2009 and 2019. The water temperatures recorded ranged from 13.5° to 18° Celsius, and underwater visibility from 2 to 5 m (measured to the nearest 0.5 m with a Secchi disc). Expeditions took place in South African autumn, between March and May. Observations occurred between 2009 and 2013 aboard “Barracuda” (Unlimited Shark Diving owner), a 12 m long boat and, between 2014 and 2019 aboard “Slashfin” (Marine Dynamics owner), a 14 m long boat. The rectangular floating cage was made of galvanised steel, housing three researchers at the time and was moored to the side of the boat. We encountered white sharks at two aggregation areas in Gansbaai (Geyser Rock and Joubertsdam) as reported by Towner et al. (Citation2013a). Sharks were attracted using olfactory stimulants (chum), following the methods described by Laroche et al. (Citation2007). The chum was a mixture of seawater, cod liver oil, fish blood, and pilchards, and, in addition, 2–3 kg slices of tuna were used as bait which was kept at the sea surface by floats (Sperone et al. Citation2010). Observations made from the boats lasted from 6 to 8 hours/day, whereas underwater observations lasted from 2 to 4 hours/day, for a total of ca. 42 hours per year. The sex of each shark was determined by the surface and by cage diving observations, and also with underwater video records of the pelvic fin area. The males were recorded if claspers were seen and the females if the lack of claspers was verified and their pelvic fin area was filmed. All other specimens were categorised as of unknown sex. The shark size was estimated to the nearest 0.5 m, as total length (TL), based on observations of the sharks when they passed in front of the measured diving cage fixed to one of the boat sides. In our study we estimated the size at sexual maturity according to Hewitt et al. (Citation2017) and this helped us to define the population structure of the area. Sharks data were collected from the same operators and dorsal fin photos were collected using three digital cameras: one CANON model EOS 550 with a SIGMA 70 lens, one CANON model 70D with 18–125 mm lenses, and one CANON model 700D with 55–250 mm lenses. Images were then analysed with Photoshop and Excel was used to archive photographs. Each collected photo was numbered and completed with the following information: date, sex, and size of the shark, presence of scars, and weather conditions. Identification of individuals was based on a larger pattern including not only the different notches of the dorsal fin, but also the following characters: caudal fin features, pelvic fin patterns, presence or absence of claspers, gill slashes and body patterns, presence of scars, and/or ectoparasites (Sperone et al. Citation2012). In 2017, a study was published by Hewitt et al. on the white shark population around False Bay (Cape Town) and similar data collecting methods were carried out by our research group.
Inter-annual sighting by sex
In order to analyse the inter-annual trend in sighting rates by sex over time, we used a log-linear model (a Poisson Generalized Linear Model) presented in Appendix 1, in which the outcome is expressed as a logarithm and the covariates are observable discrete variables. A Chi-squared test was also performed in order to check independency of maturity and sex.
Three stages for both males (M) and females (F) were considered according to Hewitt et al. (Citation2017)
MALE:
juvenile if the TL was ≤299 cm,
sub-adult if the TL was between 300 and 349 cm, and
mature if the TL was ≥350 cm.
FEMALE:
juvenile if TL was ≤399 cm,
sub-adult if the TL was between 400 and 449 cm, and
mature if the TL was ≥450 cm.
The young-of-the-year (YOY) measuring between 130 and 160 cm was included in juvenile stages, because during the eleven years of data collection only nine YOY specimens overall were observed, and few ecological information were available for this life-history stage.
Results
Annual trends in Gansbaai area
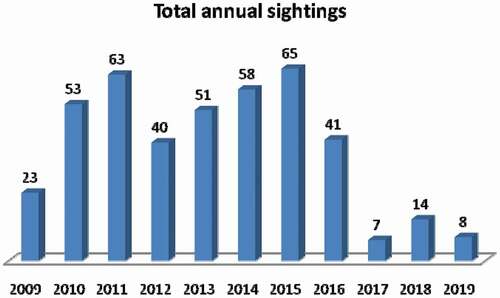
Overall, during the 11 years of sampling with a total of 462 observation hours and an average of 42 hours per year, 423 white sharks have been sighted in the following numbers: 105 males, 234 females, and 84 unsexed sharks; 99 unique dorsal fins were collected and five sharks were re-sighted, three in 2015 and only one in both 2016 and 2017. The highest and lowest annual sightings were 65 and seven white sharks in 2015 and 2017 respectively (). Although sightings varied between 2009 and 2016, with two peaks in 2011 (63 sightings) and 2015 (65 sightings), a consistently decreasing trend was observed from 2017 to 2019 ().
Table I. Observation hours, Sightings, Photo-Id sightings, and re-sightings of white sharks (Carcharodon carcharias) in Gansbaai (2009–2019)
Figure 2. Total annual sightings of white sharks (Carcharodon carcharias) in Gansbaai (2009–2019)
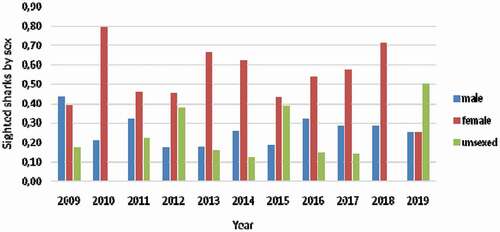
Focusing on the annual mean sighting of white sharks grouped by sex (), four main results were found: (1) the sighted female white sharks were always higher than the sighted males; (2) female sighting rates were higher than the mean (0.50) over time (6 years overall); (3) when female sighting rates were lower than the mean, for example, in 2012, 2015, and 2019, unsexed sighting rates were larger and they had probably been females; (4) male sighting was recorded during the 11 years of observations and rates were generally constant over time, contrary to female ones.
Figure 3. Mean annual sightings rates of white sharks (Carcharodon carcharias) by sex in Gansbaai (2009–2019)
Sex ratio, total length, and sexual maturity stage
The annual mean sex ratio over the eleven-year study period was: 1:2.2:0.8 for males, females, and unsexed, respectively. According to , given a total of 339 sighted and sized sharks (excluding the unsexed ones), the highest percentage of juvenile sharks was observed for females (81% for females and 14% for males, respectively). On the contrary, the males would seem to be mainly mature (22% for males vs. 7% for females). The Chi-squared test confirmed the dependence between sex and maturity (p-value = 0.001) and therefore it was statistically significant that males were mostly mature and females were immature.
Figure 4. Total length-frequency distribution (with corresponding data on sex) of white sharks (Carcharodon carcharias) in Gansbaai (2009–2019)
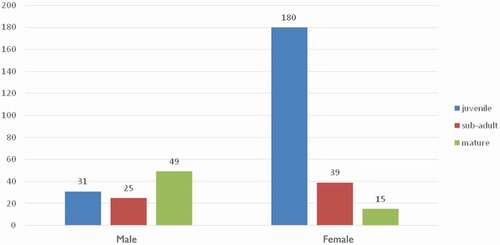
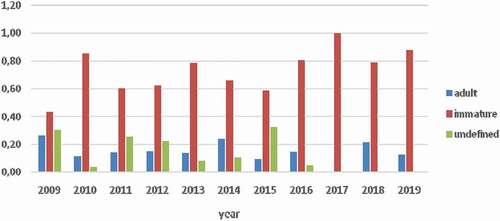
Finally, we considered the mean total length distribution (in percentages) of the sighted sharks by sex according to three distinct groups: (i) mature, (ii) immature, and (iii) undefined maturity. The results showed that most of the sharks observed over time were immature (). The results confirmed the findings found previously and the same ratios are noted by observing the adult sharks: on 64 sharks, 49 were males. For the group accounting for sharks of undefined maturity, a significant result was observed: of 67 sharks, 43 were unsexed.
Figure 5. Mean total length (TL) distribution (in %) of white sharks (Carcharodon carcharias) in Gansbaai (2009–2019)
Discussion
Sex ratio, sighting trend, and peak periodicity
During our research period the female sizes were always larger than the sighted males. The females sighting rates were higher than the mean (0.50) for most of the years, six of 11 years observations. When females sighting rates were lower than the mean, for example, in 2012, 2015, and 2019, unsexed sighting rates were relatively larger in size and thus they could have been females, also because a mature male (>350 cm) will have extremely large claspers which would be difficult to miss. It is interesting to observe that the male sighting rate was generally constant over the years in contrast to that of the female, slowly decreasing. The mean sex ratio was 1:2.2:0.8 for males, females, and unsexed. Ferreira and Ferreira (Citation1996) also reported that a degree of sex segregation occurred at both Dyer Island and Struis Bay, with females outnumbering males when collecting data during autumn-winter peak sightings season. In all the Dyer Island area studies, Ferreira and Ferreira (Citation1996), Johnson (Citation2001), Sperone et al. (Citation2010), and our research group confirmed that females outnumbered males. Towner et al. (Citation2013b) confirmed that during autumn (from March to May) and winter (from June to August) sub-adults, juveniles, and adults of both sexes were sighted at Dyer Island. Towner et al. (Citation2013b) indicated that May is the most important sighting period of the year, reporting that the white shark mean sightings per trip was 8.4. In other white sharks South African aggregation such as Seal Island in False Bay, a seasonal pattern is also exhibited, with peaks in relative abundance occurring during austral autumn and winter that coincide with a seasonal increase in abundance and availability of prey in the form of juvenile, predator-naïve Cape fur seals (Kock et al. Citation2013). Hewitt et al. (Citation2017) reported that the annual mean sex ratio over the nine-year study period was 1.0:1.0:2.3 for male: female: unsexed sharks, respectively, for Seal Island. No explanations are available to clarify the population differences between the two sites, Gansbaai and Seal Island. During the 11 years of data collection, periodic sightings peaks were observed, followed by a decrease in total sightings. The peak of sightings was observed in 2011 with 63 sightings and in 2015 with 65 sightings (). These events were also described by other authors along the coast of South Africa. Annual catch rates of white sharks C. carcharias in shark nets, set along the KwaZulu-Natal coast, varied considerably from 1966 to 1993 (Cliff et al. Citation1996) and those authors observed that a cyclical trend peaked with 4–6 years intervals. The similar peak year interval is not surprising, as most white sharks that are part of Gansbaai aggregations also move into KZN shark netted areas (Ocearch accessed, Citation2013; Towner et al. Citation2013b). Hewitt et al. (Citation2017) suggested that sighting rates peaked at five-year intervals for males (2005/2010) and six-year intervals for unsexed sharks (2006/2012), but there was no obvious cyclical pattern for females at Seal Island. It was established by various studies (Klimley et al. Citation1992; Skomal et al. Citation2012; Kock et al. Citation2013) that the best time during the year to observe white sharks coincides with the moment of maximum availability of prey, in particular the juveniles of Cape fur seals (Arctocephalus pusillus pusillus) (De Vos et al. Citation2015a, Citation2015b). Therefore, assuming a strong correlation between the presence of young Cape fur seals and the presence of sharks, it should be noted that no demographic collapses have been observed in the seal colony. Pup counts in South Africa suggest that there has been a little change in the overall population of these Cape fur seals since 1993 (Kirkman et al. Citation2007) and therefore the dependence of the minimum peaks of sightings from the availability of this type of prey can be excluded. Most of the white sharks sighted undergo a change of diet from piscivorous to marine mammals (Tricas & McCosker Citation1984) and it cannot be excluded that the presence of other prey (bony fish or other sharks) influences the presence of C. carcharias in the area. As evidence of the absence of correlation between the availability of young fur seals and the presence of sharks, the trend in the Kwazulu-Natal area is similar, despite the absence of colonies of seals (Cliff et al. Citation1996). However, it would take several decades of sightings to statistically confirm the existence of peaks. Bruce and Bradford (Citation2012) demonstrated extended residency periods of white sharks in an Australian study site where “chumming” and tourism activities had expanded. The 4–6 years peak hypothesis suggested by several authors for KZN, False Bay, and Gansbaai could be important concerning cage-diving ecotourism and white shark residence times in Gansbaai where chumming is used, but where the white shark frequency had not increased.
Total length and life history
Between 2009 and 2019, white sharks ranged from 130 to 500 cm TL but immatures, both for males and females, outnumbered mature specimens; the mode with a mean TL = 308 cm (σ 28.8 cm). At Dyer Island, an average length of 320 cm in TL was reported by Ferreira and Ferreira (Citation1996), 336 cm by Johnson (Citation2001), and 278 cm by Towner et al. (Citation2013b). Data collected by several research groups on the white shark population in Dyer Island between 1996 and 2019 would show a trend towards a decrease in the total length (Ferreira & Ferreira Citation1996; Johnson Citation2001; Sperone et al. Citation2010; Towner et al. Citation2013a). In False Bay the mode was 325 cm TL (Hewitt et al. Citation2017), while in Mossel Bay there was a large proportion of sharks (78%) ranging between 175 and 324 cm (Ryklief et al. Citation2014) (). Concerning YOY (<160 cm TL) in False Bay they were absent and only 1% of sharks were considered young in their first two years of life (YOY + 1) (<199 cm TL), while 25% were small juveniles (200–299 cm TL) (Hewitt et al. Citation2017). Different from False bay, nine YOY during the eleven years of study were observed, less than 2% of the total. This suggests that Gansbaai is not a nursery or pup area. Ferreira and Ferreira (Citation1996) suggested that the relatively few white sharks ˂ 200 cm and ˃ 400 cm TL may be the result of white shark hunting over the previous three decades along the South African coastline. Alternatively, Hewitt et al. (Citation2017) supposed that hunting was not the primary reason for the scarcity of particularly large white sharks in South Africa, because sufficient time had passed since protection for large numbers of individuals of both sexes to have reached adult size, assuming that hunting had negatively affected the adult stock before protection. The several possible questions that arise from the available data suggest that the ecology of the life history of white sharks is not yet clear for these South African populations, as there is a total lack of data on the link between the status of fish population prey and white sharks, and their influence on their life cycles. The low number of re-sightings that we recorded in Gansbaai, five in 11 years corresponding to 1.2% of the population, showed that the white shark population attending this area has a transient behaviour. In False Bay, 215 (71%) individuals were sighted in a single year only, also indicating a transient behaviour, and 88 (29%) sharks were re-sighted in at least one other year (Hewitt et al. Citation2017).
Table II. White Sharks mean TL in aggregation areas of South Africa; * in Mossel Bay 78% measured between 175 and 324 cm
Dramatic loss of white shark sightings since 2017
A dramatic decline in white shark sightings was observed in Gansbaai since 2017. The cause could be linked to the presence of a pair of transient killer whales (Orcinus orca) and this hypothesis is supported by the following evidence. Before 2015, killer whales in False Bay had been only observed preying on marine mammals, in particular on Common dolphins (Delphinus delphis) and whales such as humpbacks (Megaptera novaeangliae; reviewed by Best et al. Citation2010). No records of shark predation by killer whales exist historically for False Bay, and neither researchers nor shark-based tour operators reported marked changes in the presence or behaviour of sharks associated with the more frequent visits by killer whales in the last decade. It is indicative that groups of killer whales co-existed with resident shark species, most likely through niche partitioning (Engelbrecht et al. Citation2019). The arrival of two new individual killer whales in False Bay occurred in January 2015. Both individuals have collapsed dorsal fins, but in opposite directions, making them easily identifiable (Engelbrecht et al. Citation2019). Between 2015 and 2017, the couple of orcas was sighted at a number of different locations along the west and south coasts of South Africa, including areas such as Struis Bay and Gansbaai, and their presence at these two sites was followed by the discovery of multiple shark carcasses just between February and June 2017; in these instances, the prey were white sharks (Engelbrecht et al. Citation2019). Necropsies on the white shark carcasses in Gansbaai showed that they bore similar wounds to those observed on the seven-gill sharks in False Bay and, in all cases, the shark livers had been removed (A. Towner, public communication, Times Live, https://www.timeslive.co.za/news/sci-tech/ 2018–04-16-the-return-of-the-great-white-sharks/). The evidence from the carcasses at Millers Point (South Africa) and Gansbaai, in which only the liver of the shark was consumed while the rest of the carcass was discarded, is indicative of highly selective feeding habits by these killer whales. However, the gaps in the understanding of killer whale occurrence, behaviour, and movement patterns in South African waters make it difficult to predict why there has been an increase in their presence in coastal areas in recent years, or whether these incidents of shark predation will become more frequent in the future (Engelbrecht et al. Citation2019). Jorgensen et al. (Citation2019) stated that in multiple instances, brief visits from killer whales displaced white sharks from southeast Farallon Island (off San Francisco, California, United States), disrupting shark feeding behaviour for extended periods at this aggregation site. Although infrequent, killer whale attacks on white sharks do occur. An adult killer whale killed and partially ingested a 3–4 m white shark (C. carcharias) near southeast Farallon Island (Pyle et al. Citation1999). Although attacks are rare, Fertl et al. (Citation1996) speculated that elasmobranchs are likely underestimated in the diet of certain killer whale populations because of the difficulty of detecting underwater predation events and this assertion is supported by further studies (Visser et al. Citation2000; Visser Citation2005; Ford et al. Citation2011). The attacks in South Africa were linked to the pair of transient killer whales observed between 2015 and 2019 which have a different ecotype from that of the killer whale population residing along the South African coast and they are responsible for the loss of sightings, due to displacing the white sharks from Gansbaai. Therefore, ongoing research is needed to obtain both: a better understanding of the drivers behind killer whale presence along the South African coast, the impact on the long-term behaviour, the movement patterns of apex shark species such as seven-gills and white sharks and whether there are other cofactors responsible for this sudden and dramatic decline in white shark sightings. However, there are also other pressures that could act on Gansbaai’s white sharks as well, but the major threat could be represented by “biological resource use” and, more specifically, “fishing and harvesting aquatic resources” (Pacoureau et al. Citation2021). Recently, White et al. (Citation2019), stated that many species of sharks are threatened by overexploitation, but the degree of overlap between industrial fisheries and pelagic fishes remains poorly understood. Regarding white sharks, juveniles may be vulnerable as bycatch to various fisheries such as longlining, trawling, seine-netting and recreational angling; moreover, new developing fisheries that overlap with white shark migration routes should also be evaluated (Huveneers et al. Citation2018; Malara et al. Citation2021). The increased frequency and incidence of these activities may also have affected the white shark population of Gansbaai. Also, many indirect threats such as, above all, long-term exposure to pollutants should be considered. As top predators, white sharks are, in fact, likely to be exposed to high levels of heavy metals and organic pollutants, although the impact on their health and longevity is not completely determined (Bevacqua et al. Citation2021; Consales & Marsili Citation2021). For the population of white sharks of Gansbaai, Marsili et al. (Citation2016) have found levels of organochlorines and polycyclic aromatic hydrocarbons higher than those found in the literature. In the same study, the pattern of DDT and its metabolites showed high percentages of not only pp’DDE but also op’ DDE and this indicates a recent and continue DDT input in the environment. Finally, it cannot be excluded that such a drastic reduction in the presence of white sharks in Gansbaai may also be related to the effects of climate change. Although it is speculated that, compared to other elasmobranch species, the white shark populations are less vulnerable to direct effects of climate change (Chin et al. Citation2010; Hazen et al. Citation2012) specimens using coastal areas in shallow waters, like in Gansbaai, could likely be more exposed to and affected by climate change. Recent studies (Rouault et al. Citation2009) have shown that surface waters along South Africa’s subtropical east coast are warming significantly and this has been linked to warming and strengthening of the Agulhas current. In contrast, sections of the country’s south and west coast are cooling seasonally as winds that favour upwelling increase. In response to warming waters, changes in the distribution and abundance of tropical and temperate species have already been recorded (James et al. Citation2013). Evidence of influence of climate change on juvenile white sharks’ distribution has been observed also for a similar period (2014–16) in North Pacific by Tanaka et al. (Citation2021). In particular, the growing presence of juvenile sharks above 34° N in Central California suggested that climate change may be revising basic aspects of the established spatial population structure for the local white shark population. Moreover, this spatial shift is significant as it creates potential conflicts with commercial fisheries, protected species conservation, and public safety concerns. Therefore, the drastic decline in white shark sightings in Gansbaai could be the result of a number of interlinked factors.
Conclusions
In Gansbaai, between 2009 and 2019, the large numbers observed of few young-of-the-year sharks and few adults suggest that this area is an important seasonal feeding ground among individual sharks rather than an adult aggregation site, or a pupping or nursery area. The low number of re-sightings, only five in 11 years of 423 sharks sighted, confirms the transient behaviour of white shark population occurring in Gansbaai. The white shark sighting trend with periodic peaks followed by a slow decrease in total sightings, seems to show a cyclical trend with 4–6 years intervals between two peaks. The lack of correlation between the size of the Cape fur seals population and the variations in sightings of white sharks over the years suggest that other events are responsible, perhaps related to the diversity of the white shark diet which also includes fish and other elasmobranchs, or environmental factors, whose contribution must be researched. The peak hypothesis confirms that in Gansbaai, where cage diving with chumming is present, the white shark frequency is not increased by daily ecotourism, in contrast with the Australian study site where chumming and tourism activities expanded their resident periods. The dramatic loss of sighting from 2017 was linked also but not only, to the presence of a couple of transient killer whales with an ecotype including elasmobranchs in their diet. They were responsible for a number of fatal attacks and the subsequent displacement of white sharks from Gansbaai and other white sharks from South African aggregation sites, as Seal Island in False Bay. Other cofactors such as oceanic overfishing, pollution and climate change could probably also play an important role.
Acknowledgements
This research was made possible with the financial support of Sharks Studies Center – Scientific Institute, University of Calabria and University of Siena. We are grateful to the CSS team members that carried out 11 expeditions (Sperone, Tripepi, Giglio, Iosa, Pecchia, Rabboni, Giglioli, Da Ros, Manca, Leone, Camigliano, Chiarabelli, Mele, Visentin) for their indirect financial support of this research, and thanks are also due to Marine Dynamics (Hennie Otto, Wilfred Chivel) and Shark Diving Unlimited (Michael Rutzen) for their logistical assistance and all other field assistance with data collection. All research protocols were approved by the University of Calabria and Siena.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Anderson CJR, Lobo NDV, Roth JD, Waterman JM. 2010. Computer-Aided photo-identification system with an application to polar bears based on whisker spot patterns. Journal of Mammalogy 91(6):1350–1359. DOI:10.1644/09-MAMM-A-425.1.
- Anderson SD, Chapple TK, Jorgensen SJ, Klimley AP, Block BA. 2011. Long-term individual identification and site fidelity of white sharks, Carcharodon carcharias, off California using dorsal fins. Marine Biology 158(6):1233–1237. DOI:10.1007/s00227-011-1643-5.
- Andreotti S, Rutzen M, Van Der Walt S, Von Der Heyden S, Henriques R, Meÿer M, Oosthuizen H, Matthee CA. 2016. An integrated mark-recapture and genetic approach to estimate the population size of white sharks in South Africa. Marine Ecology Progress Series 552:241–253. DOI: 10.3354/meps11744.
- Bass AJ. 1975. Sharks of the east coast of southern Africa. IV. The families Odontaspididae, Scapanorhynchidae, Isuridae, Cetorhinidae, Alopiidae, Orectolobidae and Rhiniodontidae. Investigation Report Oceanographic Research Institute 39:1–102.
- Becerril-Garcìa EE, Hoyos-Padilla EM, Micarelli P, Galvàn-Magaña F, Sperone E. 2020. Behavioural responses of white sharks to specific baits during cage diving ecotourism. Scientific Reports 10(1):1–11. DOI:10.1038/s41598-020-67947-x.
- Best PB, Meÿer MA, Lockyer C. 2010. Killer whales in South African waters-a review of their biology. African Journal of Marine Science 32(2):171–186. DOI:10.2989/1814232X.2010.501544.
- Bevacqua L, Reinero FR, Becerril-García EE, Elorriaga-Verplancken FR, Juaristi-Videgaray D, Micarelli P, Galván-Magaña F, Curiel-Godoy P, Giglio G, Tripepi S, Barca D, Sperone E. 2021. Trace elements and isotopes analyses on historical samples of white sharks from the Mediterranean Sea. The European Zoological Journal 88(1):132–141. DOI:10.1080/24750263.2020.1853265.
- Bonfil R, Meÿer MA, Scholl MC, Johnson R, O’Brien S, Oosthuizen H, Swanson S, Kotze D, Paterson M. 2005. Transoceanic migration, spatial dynamics, and population linkages of white sharks. Science 310(5745):100–103. DOI:10.1126/science.1114898.
- Bruce BD, Bradford RW. 2012. Habitat use and spatial dynamics of juvenile white sharks, Carcharodon carcharias, in eastern Australia. In: Domeier ML, editor. Global perspectives on the biology and life history of the white shark. CRC Press. pp. 225–253.
- Chapple TC, Jorgensen SJ, Anderson SD, Kanive PE, Klimley AP, Botsford LW, Block BA. 2011. A first estimate of white shark, Carcharodon carcharias, abundance off Central California. Biology Letters 7(4):581–583. DOI:10.1098/rsbl.2011.0124.
- Chin A, Kyne PM, Walker TI, McAuley RB. 2010. An integrated risk assessment for climate change: Analysing the vulnerability of sharks and rays on Australia’s Great Barrier Reef. Global Change Biology 16(7):1936–1953. DOI:10.1111/j.1365-2486.2009.02128.x.
- Cliff G, Van Der Elst RP, Govender A, Witthuhn TK, Bullen EM. 1996. First estimates of mortality and population size of white sharks on the South African coast. In: Klimley AP, Ainley DG, editors. Great white sharks: The biology of carcharodon carcharias. San Diego: Academic Press. pp. 393–400.
- Consales G, Marsili L. 2021. Assessment of the conservation status of Chondrichthyans: Underestimation of the pollution threat. The European Zoological Journal 88(1):165–180. DOI:10.1080/24750263.2020.1858981.
- De Vos A, O’Riain MJ, Meÿer MA, Kotze PG, Kock AA. 2015a. Behavior of Cape fur seals (Arctocephalus pusillus pusillus) in response to spatial variation in white shark (Carcharodon carcharias) predation risk. Marine Mammals Science 31(3):1234–1251. DOI:10.1111/mms.12215.
- De Vos A, O’Riain MJ, Meÿer MA, Kotze PG, Kock AA. 2015b. Behaviour of Cape fur seals (Arctocephalus pusillus pusillus) in relation to temporal variation in predation risk by white shark (Carcharodon carcharias) around a seal rookery in False Bay, South Africa. Marine Mammals Science 31(3):1118–1131. DOI:10.1111/mms.12208.
- Engelbrecht TM, Kock AA, O’Riain MJ. 2019. Running scared: When predators become prey. Ecosphere 10(1):e02531. DOI:10.1002/ecs2.2531.
- Estrada JA, Rice AN, Natanson LJ, Skomal GB. 2006. Use of isotopic analysis of vertebrae in reconstructing ontogenetic feeding ecology in white sharks. Ecology 87(4):829–834. DOI:10.1890/0012-9658(2006)87[829:UOIAOV]2.0.CO;2.
- Fergusson I, Compagno LJV, Marks M. 2009. Carcharodon carcharias. The IUCN Red List of Threatened Species 2009:e.T3855A10133872.
- Ferreira CA, Ferreira TP. 1996. Population dynamics of white sharks in South Africa. In: Klimley AP, Ainley DG, editors. Great white sharks: The biology of Carcharodon carcharias. San Diego: Academic Press. pp. 381–391.
- Fertl D, Acevedo-Gutierrez A, Darby FL. 1996. A report of killer whales (Orcinus orca) feeding on a carcharhinid shark in Costa Rica. Marine Mammal Science 12(4):606–611. DOI:10.1111/j.1748-7692.1996.tb00075.x.
- Ford JKB, Ellis GM, Matkin CO, Wetklo MH, Barrett-Lennard LG, Withler RE. 2011. Shark predation and tooth wear in a population of northeastern Pacific killer whales. Aquatic Biology 11(3):213–224. DOI:10.3354/ab00307.
- Griffiths CL, Robinson TB, Lange L, Mead A. 2010. Marine biodiversity in South Africa: An evaluation of current states of knowledge. PloS One 5(8):e12008. DOI:10.1371/journal.pone.0012008.
- Hammerschlag N, Martin RA, Fallows C. 2006. Effects of environmental conditions on predator-prey interactions between white sharks (Carcharodon carcharias) and Cape fur seals (Arctocephalus pusillus pusillus) at Seal Island, South Africa. Environmental Biology of Fishes 76(2):341–350. DOI:10.1007/s10641-006-9038-z.
- Hazen EL, Maxwell SM, Bailey H, Bograd SJ, Hamann M, Gaspar P, Godley BJ, Shillinger GL. 2012. Ontogeny in marine tagging and tracking science: Technologies and data gaps. Marine Ecology Progress Series 457:221–240. DOI: 10.3354/meps09857.
- Hewitt AM, Kock AA, Booth AJ, Griffiths CL. 2017. Trends in sightings and population structure of white sharks, Carcharodon carcharias, at Seal Island, False Bay, South Africa, and the emigration of subadult female sharks approaching maturity. Environmental Biology of Fishes 101(1):39–54. DOI:10.1007/s10641-017-0679-x.
- Huveneers C, Apps K, Becerril-García EE, Bruce B, Butcher PA, Carlisle AB, Chapple TK, Christiansen HM, Cliff G, Curtis TH, Daly-Engel TS, Dewar H, Dicken ML, Domeier ML, Duffy CAJ, Ford R, Francis MP, French GCA, Galván-Magaña F, García-Rodríguez E, Gennari E, Graham B, Hayden B, Hoyos-Padilla EM, Hussey NE, Jewell OJD, Kock AA, Lowe CG, Lyons K, Meyer L, Oelofse G, Oñate-González EC, Oosthuizen H, O’Sullivan JB, Ramm K, Skomal G, Sloan S, Smale MJ, Sosa-Nishizaki O, Sperone E, Tamburin E, Towner AV, Wcisel MA, Weng KC, Werry JM. 2018. Future research directions on the “elusive” white shark. Frontiers in Marine Science 5:455. DOI: 10.3389/fmars.2018.00455.
- Irion DT, Noble LR, Kock AA, Gennari E, Dicken ML, Hewitt AM, Towner AV, Booth AJ, Smale MJ, Cliff G. 2017. Pessimistic assessment of white shark population status in South Africa: Comment on Andreotti et al. (2016). Marine Ecology Progress Series 577:251–255. DOI: 10.3354/meps12283.
- James NC, Van Niekerk L, Whitfield AK, Potts WM, Götz A, Paterson AW. 2013. Effects of climate change on South African estuaries and associated fish species. Climate Research 57(3):233–248. DOI:10.3354/cr01178.
- Johnson RL. 2001. The behavioural ecology of the white shark (Carcharodon carcharias) at Dyer Island. University of Pretoria. pp. 176.
- Jorgensen SJ, Anderson SD, Ferretti F, Tietz JR, Chapple T, Kanive P, Bradley RW, Moxley JH, Block BA. 2019. Killer whales redistribute white shark foraging pressure on seals. Scientific Reports 9(1):6153. DOI:10.1038/s41598-019-39356-2.
- Jury MR. 1985. Case studies of alongshore variations in wind-driven upwelling in the southern Benguela region. In: Shannon L, editor. South African ocean colour and upwelling experiment. Cape Town: Sea Fisheries Research Institute. pp. 29–46.
- Kirkman SP, Oosthuizen WH, Meÿer MA. 2006. The seal population of Seal Island, False Bay. In: Nel DC, Peschak TP, editors. Finding a balance: White shark conservation and recreational safety in the inshore waters of Cape Town, South Africa. Proceedings of a specialist workshop. WWF South Africa Report Series – 2006/Marine/001 Annexure 1. pp. 83–94.
- Kirkman SP, Oosthuizen WH, Meÿer MA, Kotze PGH, Roux JP, Underhill LG. 2007. Making sense of censuses and dealing with missing data: Trends in pup counts of Cape fur seal Arctocephalus pusillus pusillus for the period 1972–2004. African Journal of Marine Science 29(2):161–176. DOI:10.2989/AJMS.2007.29.2.2.185.
- Klimley AP, Anderson SD, Pyle P, Henderson RP. 1992. Spatiotemporal patterns of white shark (Carcharodon carcharias) predation at the South Farallon Islands, California. Copeia 3(3):680–690. DOI:10.2307/1446143.
- Kock A, O’Riain MJ, Mauff K, Meÿer M, Kotze D, Griffiths C. 2013. Residency, habitat use and sexual segregation of white sharks, Carcharodon carcharias in False Bay, South Africa. PloS One 8(1):e55048. DOI:10.1371/journal.pone.0055048.
- Kock AA, Johnson RL. 2006. White shark abundance: Not a causative factor in numbers of shark bite incidences. In: Nel DC, Peschak TP, editors. Finding a balance: White shark conservation and recreational safety in inshore waters of Cape Town, South Africa: Proceedings of a specialist workshop. WWF South Africa report series—2006/Marine/ 001. pp. 1–19.
- Laroche RK, Kock AA, Dill LM, Oosthuizen WH. 2007. Effects of provisioning ecotourism activity on the behaviour of white sharks Carcharodon carcharias. Marine Ecology Progress Series 338:199–209. DOI: 10.3354/meps338199.
- Lee KA, Huveneers C, Gimenez O, Peddemors V, Harcourt RG. 2014. To catch or to sight? A comparison of demographic parameter estimates obtained from mark-recapture and mark-resight models. Biodiversity and Conservation 23(11):2781–2800. DOI:10.1007/s10531-014-0748-9.
- Leone A, Puncher GN, Ferretti F, Sperone E, Tripepi S, Micarelli P, Gambarelli A, Sarà M, Arculeo M, Doria G, Garibaldi F, Bressi N, Dall’Asta A, Minelli D, Cilli E, Vanni S, Serena F, Diaz-Aimes P, Baele G, Cariani A, Tinti F. 2020. Pliocene colonization of the Mediterranean by Great White Shark inferred from fossil records, historical jaws, phylogeographic and divergence time analyses. Journal of Biogeography 47(5):1119–1129. DOI:10.1111/jbi.13794.
- Malara D, Battaglia P, Consoli P, Arcadi E, Longo F, Stipa MG, Pagano L, Greco S, Andaloro F, Romeo T. 2021. When opportunistic predators interact with swordfish harpoon fishing activities: Shark depredation over catches in the Strait of Messina (central Mediterranean Sea). The European Zoological Journal 88(1):226–236. DOI:10.1080/24750263.2021.1879284.
- Marsili L, Coppola D, Giannetti M, Casini S, Fossi MC, Van Wyk JH, Sperone E, Tripepi S, Micarelli P, Rizzuto S. 2016. Skin biopsies as a sensitive non-lethal technique for the ecotoxicological studies of great white shark (Carcharodon carcharias) sampled in South Africa. Expert Opinion on Environmental Biology 4:1. DOI: 10.4172/2325-9655.1000126.
- Martin RA. 2003. Field guide to the great white shark. In: Reefquest centre for shark research. British Columbia (Canada): Vancouver. p. 185.
- Martin RA, Hammerschlag N, Collier RS, Fallows C. 2005. Predatory behaviour of white sharks (Carcharodon carcharias) at Seal Island, South Africa. Journal of Marine Biological Association of the United Kingdom 85(5):1121–1136. DOI:10.1017/S002531540501218X.
- Micarelli P, Sperone E, Giglio G, Pecchia J, Romano C, Scuderi A, Vespasiani L, Mele F. 2015. Dorsal fin photoidentification: Tool for long term studies of White shark (Carcharodon carcharias) behaviour. Marina Mediterranea 22 (1) – Atti XLVI Congresso SIBM. Roma, 11–12 giugno 2015.
- Nasby-Lucas N, Domeier ML. 2012. Use of photo identification to describe a white shark aggregation at Guadalupe Island, Mexico. In: Domeier ML, editor. Global perspectives on the biology and life history of the white shark. CRC Press. pp. 381–392.
- Ocearch Shark Tracker. 2013. Available: sharks-ocearch.vertite.com. Accessed May 2013 8.
- Pacoureau N, Rigby CL, Kyne PM, Sherley RB, Winker H, Carlson JK, Fordham SV, Barreto R, Fernando D, Francis MP, Jabado RW, Herman KB, Liu KM, Marshall AD, Pollom RA, Romanov EV, Simpfendorfer CA, Yin JS, Kindsvater HK, Dulvy NK. 2021. Half a century of global decline in oceanic sharks and rays. Nature 589(7843):567–571. DOI:10.1038/s41586-020-03173-9.
- Pyle P, Schramm MJ, Keiper C, Anderson SD. 1999. Predation on a white shark (Carcharodon carcharias) by a killer whale (Orcinus orca) and a possible case of competitive displacement. Marine Mammal Science 15(2):563–568. DOI:10.1111/j.1748-7692.1999.tb00822.x.
- Rouault M, Penven P, Pohl B. 2009. Warming in the Agulhas Current system since the 1980’s. Geophysical Research Letters 36(12):L12602. DOI:10.1029/2009GL037987.
- Ryklief R, Pistorius PA, Johnson R. 2014. Spatial and seasonal patterns in sighting rate and life-history composition of the white shark Carcharodon carcharias at Mossel Bay, South Africa. African Journal of Marine Science 36(4):449–453. DOI:10.2989/1814232X.2014.967296.
- Skomal GB, Chrisholm J, Correira SJ. 2012. Implications of increasing pinniped interactions on the diet and abundance of white sharks off the coast of Massachusetts. In: Domeier ML, editor. Global perspectives on the biology and life history of the white shark. CRC Press. pp. 405–417.
- Sperone E, Micarelli P, Andreotti S, Brandmayr P, Bernabò I, Brunelli E, Tripepi S. 2012. Surface behaviour of bait-attracted white sharks at Dyer Island (South Africa). Marine Biology Research 8(10):982–991. DOI:10.1080/17451000.2012.708043.
- Sperone E, Micarelli P, Andreotti S, Spinetti S, Andreani A, Serena F, Brunelli E, Tripepi S. 2010. Social interactions among bait-attracted white sharks at Dyer Island (South Africa). Marine Biology Research 6(4):408–414. DOI:10.1080/17451000903078648.
- Tanaka KR, Van Houtan KS, Mailander E, Dias BS, Galginaitis C, O’Sullivan J, Lowe CG, Jorgensen J. 2021. North Pacific warming shifts the juvenile range of a marine apex predator. Scientific Reports 11(1):3373. DOI:10.1038/s41598-021-82424-9.
- Towner AV. 2012. Great white sharks Carcharodon carcharias in Gansbaai, South Africa: Environmental influences and changes over time, 2007–2011. University of Cape Town (UCT).
- Towner AV, Underhill LG, Jewell OJ, Smale MJ. 2013b. Environmental influences on the abundance and sexual composition of white sharks Carcharodon carcharias in Gansbaai, South Africa. PloS One 8(8):e71197. DOI:10.1371/journal.pone.0071197.
- Towner AV, Wcisel MA, Reisinger RR, Edwards D, Jewell OJD. 2013a. Gauging the threat: The first population estimate for white sharks in South Africa using photo identification and automated software. PloS One 8(6):e66035. DOI:10.1371/journal.pone.0066035.g001.
- Tricas TC, McCosker JE (1984). Predatory behaviour of the white shark (Carcharodon carcharias), with notes on its biology. Proceedings of the California Academy of Science 43:221–238.
- Visser INJ. 2005. First observations of feeding on thresher (Alopias vulpinus) and hammerhead (Sphyrna zygaena) sharks by Killer Whales (Orcinus orca), which specialise on elasmobranchs as prey. Aquatic Mammals 31(1):83–88. DOI:10.1578/AM.31.1.2005.83.
- Visser INJ, Berghan J, Van Meurs R, Fertl D. 2000. Killer whale (Orcinus orca) predation on a shortfin mako shark (Isurus oxyrinchus) in New Zealand waters. Aquatic Mammals 26(3):229–231.
- White TD, Ferretti F, Kroodsma DA, Hazen EL, Carlisle AB, Scales KL, Bograd SJ, Block BA. 2019. Predicted hotspots of overlap between highly migratory fishes and industrial fishing fleets in the northeast Pacific. Science Advances 5(3):eaau3761. DOI:10.1126/sciadv.aau3761.
Appendix 1
Table of the annual sightings by sex and the outcome of the log-linear model about the inter-annual trend in sighting rates by sex over the years.