
Abstract
In species with biparental care, one parent may relieve him-/herself of the costs of parental care by deserting and leaving the mate to care for the offspring. Offspring desertion by a single parent, usually the female, occurs in many bird species, especially in Charadriiformes. The reasons for desertion are still being debated, and compelling evidence for the benefits and costs to either sex is not yet handed. The benefits of desertion for the deserter may include renesting with a new mate shortly after desertion. Therefore, desertion may enhance individual lifetime reproductive success. In Whiskered Tern Chlidonias hybrida almost all females desert, about half of females desert during chick-rearing period, the other half during the post-fledging period. Based on intensive fieldwork over a 4-year study, we investigated whether Whiskered Tern females remated and renested after desertion. We assumed that favourable conditions, such as an extended breeding season and food abundance resulting in a high breeding success would prompt most females to renest after desertion. In contrast to this expectation, however, only 5% of females that deserted during chick-rearing period (2 out of 41) remated and renested in the study area. About 50 days elapsed between the initiations of the first and second clutch by these two females. Females also participated in courtship behaviour after desertion (without renesting). We thus estimate that 15% (6 out of 41) of females that deserted during chick-rearing period were subsequently involved in some kind of breeding activity. Therefore, renesting does not seem to be a major direct benefit for deserting Whiskered Tern females. Females of this species probably benefit from desertion in other ways, but these have yet to be discovered.
Introduction
According to life history theory, animals are faced with trade-offs in the allocation of energy reserves to different functions (Williams Citation1966; Stearns Citation1992). During the reproductive season, the allocation of resources is a trade-off between survival and the current and future production of offspring (Clutton-Brock Citation1991). In birds with bi-parental care, each partner usually benefits from the effort invested by its mate and avoids some of the associated costs (e.g. Lessells Citation2012; Cockburn Citation2006). Therefore, offspring desertion is a typical reproductive strategy when one mate relinquishes parental care, leaving the other mate to care for the chicks (CitationSzékely et al. 1996; Parker et al. Citation2002; Lessells Citation2012). The obvious benefit for the deserter is that the considerable energy outlay on parental care is reduced and in consequence, enhancing survival and increasing the probability of returning to breed in subsequent seasons (Székely & Williams Citation1995; Tavecchia et al. Citation2002). Theoretical and experimental studies have shown that offspring desertion is often beneficial for the parents as it can increase lifetime reproductive success (CitationSzékely et al. 1996; Webb et al. Citation1999). Remating and renesting in the current breeding season as a benefit of desertion has been reported in many bird species (e.g. Beissinger Citation1986 Citation1987; Fujioka Citation1989; Székely & Williams Citation1995; Griggio & Pilastro Citation2007; Morton et al. Citation2010; Béziers & Roulin Citation2016). Other benefits of desertion include departing earlier on migration (e.g. Gratto-Trevor Citation1991; Currie et al. Citation2001; Nisbet Citation2002; Nisbet et al. Citation2011a Citation2011b) or earlier moulting (e.g. Urano Citation1992; Kloskowski Citation2003; Hemborg Citation2004; Ezaki Citation1988; Mumme Citation2018). It has been shown that natural desertion by one parent does not usually have an adverse effect on breeding success and that the one continuing parental care compensates for the absence of the deserter as long as breeding conditions are suitable (Beissinger Citation1987; Fujioka Citation1989; Osorno & Székely Citation2004; Ledwoń & Neubauer Citation2017; Harrod & Mumme Citation2021; but see Székely & Williams Citation1995).
Generally speaking, renesting may indeed increase the annual reproductive output of individuals, but the additional effort may have fitness consequences (reviewed by Moreno Citation1998; Wendeln et al. Citation2000; Verhulst & Nilsson Citation2008; Becker & Zhang Citation2010). The production of extra eggs and incubation are demanding processes in terms of energy, time and nutrition, and can impair the parents’ capacity to rear the brood all the way to fledging (Monaghan & Nager Citation1997; Monaghan et al. Citation1998; Moreno Citation1998). However, some studies have shown that relaying individuals are of higher quality than non-relaying ones and that the benefits, i.e. the production of young, outweigh costs such as reduced survival and/or future reproductive output (Wendeln et al. Citation2000; Becker & Zhang Citation2010).
The probability of renesting, both after desertion and clutch brood/failure, has been found to depend on a number of factors, e.g. laying date, partner availability, body condition and food availability (CitationSzékely et al. 1996). It has been suggested that early breeders have a greater probability of renesting because they have sufficient time to complete a second brood (Beissinger Citation1990). Furthermore, the rarer sex has more opportunities to find a new mate and, in consequence, to renest (CitationSzékely et al. 1996; Amat et al. Citation1999; Eberhart-Phillips et al. Citation2017; Citation2018; McDonald et al. Citation2023). In addition, females in poor body condition have limited renesting opportunities (Arnold et al. Citation2010; Becker & Zhang Citation2010). It has been suggested that an abundance of food which result in high fledging success encourages desertion and, consequently, renesting (Eldegard & Sonerud Citation2009; Zárybnická Citation2009).
The reasons for desertion are still being debated, and compelling evidence of the costs and benefits to either sex is required (Székely et al. Citation2007; Lessells Citation2012; Harrod & Mumme Citation2021). To understand why some individuals care for their young, whereas others do not, we need to know the relative benefits of desertion accruing to deserters (CitationSzékely et al., Citation1999). Offspring desertion by one parent, mainly females, occurs in many shorebirds Charadriiformes, particularly in Charadrii (Székely & Reynolds Citation1995; CitationSzékely et al. 1996; Gavin et al. Citation2007). The benefits gained from desertion can vary among species, for example, remating and renesting after desertion, and departing earlier on migration or moulting earlier; some authors do not report obvious benefits (CitationSzékely et al. 1996; Currie et al. Citation2001; Kosztolanyi et al. Citation2006; Gavin et al. Citation2007). In this study, we investigated whether females of Whiskered Tern Chlidonias hybrida (order Charadriiformes) renested after deserting their offspring. Among larids, Whiskered Tern has a unique system of parental care since almost all females of this species desert (Ledwoń & Neubauer Citation2017) and until now desertion has not been reported in this group of birds. In Whiskered Tern, females start to desert when the chicks are ~5 days old and no longer require intensive brooding, hence the chicks development of thermoregulatory capacity (Ledwoń & Neubauer Citation2017). The half (52%) of desertion events occur during the chick-rearing period, the other half during the post-fledging period. Two weeks after fledging, there are no nests with female care. Females desert throughout the breeding season even at the end of it, when there is not enough time for renesting. Female desertion probability does not depend on chicks number, hatching date or male provisioning rate. The female provisioning rate affects chances of nest desertion significantly – daily desertion rate is lower when females supplied more food. After female desertion, males increase the chick feeding rate, thereby at least partially compensating for the absent female. Desertions before fledging did not affect fledging success. Males care for the juveniles after they have fledged for at least several weeks. In the population we studied, Whiskered Tern females that deserted were expected to renest in the current breeding season because of the propitious conditions: an extended breeding season, an abundance of food, low predation pressure, stable hydrological conditions and a high breeding success (Ledwoń & Neubauer Citation2017). In particular, females that desert at the beginning of the breeding season are more likely to find a new mate and raise a second brood. In this study, therefore, which was based on intensive fieldwork over 4 years, we investigated whether females of Whiskered Tern are involved in any kind of breeding activity (courtship flights, remating or renesting) after desertion.
Material and methods
Study species
Whiskered Tern is a small (80–90 g), long-lived, colonially nesting bird (Gochfeld et al. Citation2020). It builds floating nests among aquatic plants. Its plumage is not sexually dimorphic, but males are larger than females (Ledwoń Citation2011). Apparent pre-breeding and breeding survival is estimated to be 0.54 and 0.80, respectively (Ledwoń et al. Citation2013). It is a semi-precocial, socially and genetically monogamous species with a high level of parental investment (Minias et al. Citation2014; Ledwoń & Neubauer Citation2017; Chambon et al. Citation2020; Paillisson & Chambon Citation2021; Ledwoń & Szczys Citation2022). The female lays usually a three-egg clutch that represents about one-third of her body mass, and both parents incubate the eggs and brood the chicks to an equal extent (Spina Citation1982; Paillisson et al. Citation2007; Ledwoń Citation2010). However, females make an approximately 25% smaller contribution than males to feeding the chicks (Ledwoń & Neubauer Citation2017). Whiskered Tern is a single prey loader, foraging on aquatic animals (fish, amphibians) and insects by plunge-diving or picking them off the surfaces of the water and plants (Gwiazda & Ledwoń Citation2015). Incubation lasts for 21 days and the chick-rearing period another 21 days or so; the exact duration of the post-fledging period is not known but it does last for at least 3 weeks (Ledwoń Citation2010; Ledwoń & Neubauer Citation2017; M. Ledwoń et al. unpublished data).
Study area
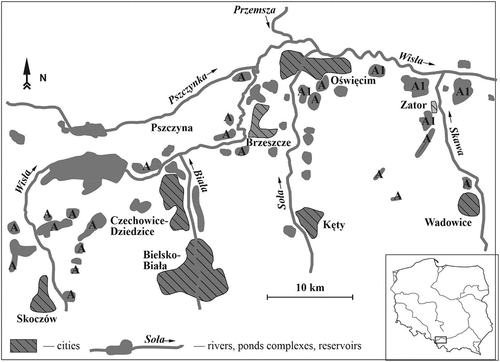
We carried out the fieldwork during 4 breeding seasons (2015–2018) in the Upper Vistula River Valley (approx. 1400 km2, 49°58ʹ21.7″ N 19°07ʹ12.5″ E, ), southern Poland. During these years, about 1000 pairs of Whiskered Tern bred there annually (M. Ledwoń et al. unpublished data), mainly on carp ponds. The ponds in the study area are grouped into over 70 complexes (total area approx. 6000 ha); the individual complexes are separated by several kilometres of land, where there are cities, villages, arable land and small deciduous forests. Each pond complex usually consists of a few to dozens of ponds. The area of each complex varies from 10 to 500 ha (mean 90 ha). The ponds are usually shallow with an average depth of about 1.3 m. The area of a single pond varies usually between 10 and 20 ha, the maximum is 90 ha.
Figure 1. Distribution of breeding sites of the Whiskered Tern Chlidonias hybrida in Upper Vistula River Valley, Southern Poland, 2015–2018. A – carp pond complexes with breeding colonies, A1 – carp pond complexes where breeding birds were marked.
Bird trapping and behavioural observations
The main aim of the fieldwork was to find out whether and how many females remated and renested after desertion, and also whether females participated in courtship displays following desertion. To do so, Whiskered Terns were trapped at nest and marked in the eastern part of the Upper Vistula River Valley () on the carp pond complexes at Spytkowice (50°00ʹ33.6″ N 19°29ʹ00.5″ E), Bugaj (49°58ʹ35.1″ N 19°25ʹ13.0” E), Przeręb (50°01ʹ05.8” N 19°24ʹ09.6” E), Stawy Monowskie (50°2ʹ7.48” N 19°20ʹ30.27” E) and Adolfin (50°00ʹ40.7” N 19°13ʹ16.2” E) hosting annually from 400 to 900 breeding pairs. The greatest distance between the colonies was 20 km. Adults on randomly chosen numbered nests were trapped using a roof trap, principally during the late incubation or downy chick stages (for details of the trapping methods see Ledwoń et al. Citation2015, Citation2016). Mainly early breeders – that laid clutches before 15 June – were trapped. During the 4 years (2015–2018), 162 females (20, 39, 42, 61 respectively) were marked with metal and colour alphanumeric ring. In all females, small areas of feathers were marked using hair dye so that we could recognize individual birds in flight from a distance. Sex of trapped birds was determined in the field based on measurements (CitationLedwoń Citation2011), additionally to confirm this, adults were also molecularly sexed using blood samples according to Goławski et al. (Citation2016) and Banach et al. (Citation2021).
To increase the effectiveness of the detection of renesting females that deserted, in 2016–2018, 64 females were additionally fitted with VHF tags (20, 23, 21, respectively; Biotrack Ltd.; PIP3 Tags Ag392 weighing 1.2 g and PIP3 Tags Ag393 weighing 1.4 g). Fifty-six tags were glued to clipped back feathers. In 2016, we attached eight tags to tail feathers with the aid of glue and string. The weight of the tags with equipment did not exceed 2% of a bird’s weight. In late July, when the birds moulted, the tags fell off. At least three females lost their tags soon after they had been attached – we observed individually marked birds without tags. The battery life span allowed for about 2 months of the transmitter’s operation. Each transmitter sent a signal on a specific frequency. Using a Yagi antenna (https://www.teltad.pl/) and a Yaesu FT-817 radio receiver we were able to detect the individual radio signal from a distance of about 3 km. After finding the direction from which a given signal was coming, the observer usually followed it to find and to observe female (for more details about radio tagging see Kenward Citation2001).
In 2015–2018 using spotting scopes and binoculars and two sets of Yagi antennas and receivers (2016–2018), observers looked for marked females in colonies to find renesting birds. Because we could not exclude that some VHF tags stopped working or fell off (see above) breeding birds were intensively viewed with optical equipment to detect renesting marked females. Observers focused mainly on newly established colonies formed in late June and July – these were visited 2–3 times per week during their formation. Because all studied females were individually marked by hair dye and the place of first breeding attempt was known (see below), in case of finding a renesting of deserter, we were able to check the condition of her first brood, i.e. whether the first male takes care of the brood or whether brood failed. Deserters were searched for in the colonies in the whole Upper Vistula River Valley. In 2015–2018 from 939 to 1269 pairs were breed there in 27–34 colonies located on 24 pond complexes (). In each year from about 15% to 30% of clutches were late – eggs were present in nests after 15 July, it means that at least some deserters have enough time for renesting.
All nests (n = 162) with marked females were monitored to obtain date of hatching and breeding success. We usually initiated this procedure in the first half of the incubation period. We erected plastic mesh fences (enclosures) around the nests with eggs to prevent the chicks from escaping until they were able to fly (see Ledwoń et al. (Citation2015) and Banach et al. (Citation2021) for a description of an enclosure). Such enclosures, set up during the second half of incubation, allow one to study breeding success and to make behavioural observations. We inspected the nests twice a week, noting the numbers of eggs and chicks in order to evaluate hatching and fledging success. The exact hatching date was deduced from observations during the nest monitoring visits: from the presence of freshly hatched chicks, readily identified as they were still wet after having just emerged from the egg; by the presence of the hatching star on the egg, which indicates that the chick will emerge from the egg within 24 h; and on the basis of chick wing length (Paillisson et al. Citation2008; Banach et al. Citation2021). The laying date was back-calculated (i.e. the laying date of the first egg of a clutch) on the assumption that incubation lasts 21 days (Betleja Citation2003).
To determine the rate of females that deserted during chick-rearing period (up to the 21st day of the chicks’ life) and the date of desertion, behavioural observations at the randomly chosen nest with marked birds were made (, detailed description of the methods, see Ledwoń & Neubauer Citation2017). Currently, the 4-year study covers the 2015 season which is also included in Ledwoń and Neubauer (Citation2017). We reanalysed the already published data (year 2015) and we added the original data obtained between 2016 and 2018. In this analysis, we included 91 nests observed during the chick-rearing period. We were not able to observe all 162 nets due to fact that some nests were hidden in rushes. We observed the group of nests from the pond shore from distances of several tens of metres to approx. 150 m using binoculars (10x42, 15 × 56) and spotting scopes (x30, x50), every 1–4 days in the morning, for an average of 190 min per observation session (range 60–350 min). As the focal colonies had been established mainly on floating leaves of Fringed Water Lily Nymphoides peltata and Water Chestnut Trapa natans, the nests were plainly visible. We defined desertion as having occurred when one parent did not attend the chicks on two consecutive days. Desertion was assumed to have taken place midway between the date on which the partner was seen for the last time and that of the next observation (Székely & Lessells Citation1993). In each case, one bird continued parental care after mate desertion (see also Ledwoń & Neubauer Citation2017). We did not note biparental desertion. Nests were observed during the chick-rearing period until the chicks fledged, i.e. usually ≤21 days after hatching. We did not study desertions after the chicks had fledged.
Table I. Desertion of Whiskered Tern Chlidonias hybrida females during the chick-rearing period, eastern part of the Upper Vistula River Valley.
Results
Desertion
Only females deserted their nests (), doing so during the chick-rearing period in 45% of the nests observed (41 out of 91). Each year from 31% to 52% of females deserted their brood during the chick-rearing period.
Renesting after desertion
Two confirmed cases of females remating and renesting after desertion were found (). In both cases, the renesting females had been early breeders, deserting 7 and 10 days after their chicks had hatched; they renested up to 2 km away from their previous breeding sites. One female laid eggs 17 days after desertion. In the case of other females, we were not able to determine the exact date of laying. In both cases, the males of the first broods were able to raise their chicks to fledging. The second brood of one deserting female failed, probably because of predation by Magpie Pica pica, whereas the fate of the other deserting female’s second brood could not be ascertained. From these observations, we can tentatively infer that after desertion during the chick-rearing period, at least 5% of females remated and renested in the Upper Vistula River Valley in the breeding season (2 out of 41 deserting females; , ).
Table II. Cases of Whiskered Tern Chlidonias hybrida females involved in some kind of breeding activity after desertion, eastern part of the Upper Vistula River Valley, 2015–2018.
In addition, we recorded 4 cases of marked females that participated in courtship flights or in courtship feeding after desertion (). In two of them, this courtship behaviour was observed just 3 and 5 days after desertion. At a rough estimate, 15% of females that deserted during the chick-rearing period (6 out of 41) were involved in some breeding activity – nesting, courtship flights and courtship feedings. These females were also early breeders.
Additionally, we recorded 3 females that deserted and renested after their first broods failed (). In all three cases, the males continued parental care after females’ desertion but the chicks died a few days after the desertion. One female renested with her first partner. As far as the other two females were concerned, we were unable to determine whether they had renested with the same mate because in both cases the males were unmarked. All these females laid clutches from 10 to 20 days after desertion and from 7 to 13 days after brood failure.
Table III. Whiskered Tern Chlidonias hybrida females that deserted and renested after brood failure, eastern part of the Upper Vistula River Valley, 2015–2018. *Female remated with the same male.
Discussion
We have shown that even though the population studied here has an extended breeding period, only c. 5% of females remated and renested after desertion in their current breeding area. By comparison, in related Kentish Plover Charadrius alexandrinus, Snowy Plover Charadrius nivosus, Dunlin Calidris alpina pacifica, in which the majority of females also desert during chick-rearing period, at least 27%, 37%, 8–33% of females renest after desertion in the current breeding area, respectively (Warriner et al. Citation1986; Székely & Williams Citation1995; Jamieson Citation2011). Renesting after offspring desertion was also noted in several other species, e.g. Little Egret Egretta garzetta (Fujioka Citation1989), Snail Kites Rostrhamus sociabilis (Beissinger & Snyder Citation1987), Barn Owl Tyto alba (Bèziers & Roulin Citation2016), Tengmalm’s Owl Aegolius funereus (Korpimäki et al. Citation2011), Rock Sparrow Petronia petronia (Pilastro et al. Citation2001), Blue-headed Vireo Vireo solitarius (Morton et al. Citation2010). However, this behaviour was not observed in a number of other species, e.g. Red-necked Grebe Podiceps grisegena (Kloskowski Citation2003), Purple Sandpipers Calidris maritima (Rae et al. Citation2011), Semipalmated Sandpiper Calidris pusilla (Gratto-Trevor Citation1991), Western Sandpiper Calidris mauri (Franks et al. Citation2020), Eurasian Curlew Numenius arquata (Currie et al. Citation2001), Little Auk (Dovekie) Alle alle (Wojczulanis-Jakubas & Jakubas Citation2012), Magnificent Frigatebird Fregata magnificens (Osorno & Székely Citation2004), Great Reed Warbler Acrocephalus arundinaceus (Urano Citation1992) and Hooded Warbler Setophaga citrina (Harrod & Mumme Citation2021).
Renesting does not appear to be a major direct benefit that Whiskered Tern females gain as a result of desertion, especially as renesting after desertion was unsuccessful. It cannot be also excluded that some females that deserted renested in the study area without being detected. However, in our opinion, possible few cases do not affect significantly the percentage of females engaging in breeding activity after desertion. We also cannot exclude the possibility that deserting females renest several hundred kilometres away in Southern Europe, e.g. in Hungary or Romania (examples from other species – Beissinger & Snyder Citation1987; Franz et al. Citation1987; Stenzel et al. Citation1994). But even though every year since 2010 a few dozen breeding females at least have been rung with coloured alphanumeric ringed in the Upper Vistula River Valley, we have received no reports of their breeding in distant colonies during the current breeding season. Furthermore, data from females equipped with geolocators did not confirm renesting in distant colonies during the current breeding season either (M. Ledwoń et al. unpublished data).
Some studies have shown that renesting is constrained mainly by time, and that renesting propensity is related most closely to the clutch initiation date (Amat et al. Citation1999; Wendeln et al. Citation2000; Arnold et al. Citation2010; Jamieson Citation2011; Béziers & Roulin Citation2016). We found that females who renested or were participating in courtship after desertion had been early breeders (). Because biparental care lasts a long time, about 50 days elapsed between the initiations of the first and second clutches. This interval was of a similar duration to that found in the related Kentish Plover (61–66 days, Amat et al. Citation1999), Snowy Plover (40 days, Page et al. Citation1995). In Barn Owl Tyto alba, which is not related to Whiskered Tern, females laid clutch at an interval of 85 days (Béziers & Roulin Citation2016). Whiskered Terns lay eggs in the study area from late May to the beginning of August (Betleja Citation2003; M. Ledwoń et al. unpublished data) and the latest chicks fledge in mid-September. Around 40% of females that deserted in the studied population (that laid eggs before 25 June) appeared to have sufficient time to renest so that the chicks would fledge before mid-September (Betleja Citation2003; M. Ledwoń et al. unpublished data). Conversely, late breeding may not be advantageous: the availability of food may decrease with breeding season advancement, and late nests could be at greater risk of predation because of the diminishing dilution effect (review in Moreno Citation1998). In our study population, chicks mortality indeed increase in relation to breeding season advancement (M. Ledwoń et al. unpublished data).
Several studies have shown that females desert more often than males because they can easily find new mates as a result of a male-biased operational sex ratio (Warriner et al. Citation1986; Fujioka Citation1989; CitationSzékely et al. 1996; Amat et al. Citation1999; Pilastro et al. Citation2001; Székely et al. Citation2014; Eberhart-Phillips et al. Citation2017; Citation2018). The operational sex ratio during breeding in Whiskered Tern is not known. It is very difficult to determine whether there is an excess of males in the period when females are deserting and new colonies forming. However, the sex ratios in hatchlings and fledglings do not differ from parity (M. Ledwoń et al. unpublished data).
Among other factors that may limit female renesting opportunities, there is the female’s poor body condition due to the considerable energy outlay on egg production (e.g. Arnold et al. Citation2010; Becker & Zhang Citation2010). The potentially costly egg production in Whiskered Tern seems to be at least partially compensated for during incubation, since female body condition improves during incubation (M. Ledwoń et al. unpublished data). Furthermore, common renesting after clutch or brood failure in this species suggests that cost of relaying is probably relatively low. Additionally, it has been shown in a few other species that renesting after desertion does not depend on female body condition (e.g. Amat et al. Citation1999, Citation2000). On the other hand, in the study population, we found that the high probability of a female desertion during chick-rearing period was correlated with the low amount of food provided to the chicks, which may suggest that females that desert earlier are in poorer condition than those that desert later (Ledwoń & Neubauer Citation2017). In addition, females that desert in the results of prolonged stress were in poorer condition than those that did not desert at that time (Ledwoń et al. Citation2019). All this may suggest a link between poor condition and desertion and thus insufficient energy reserves (low body condition) to relay. However, the cost of relaying has not been investigated comprehensively in the study population.
Renesting could also be limited by the availability of food (e.g. Eldegard & Sonerud Citation2009; Zárybnická Citation2009). In the study area, food availability does not appear to be a factor limiting female renesting. The great abundance of food in the carp ponds may be a factor favouring the high frequency of desertion. This enables males to increase their provisioning rates and at least partly compensate for the desertion of their mates (Ledwoń & Neubauer Citation2017). However, some studies in other species have shown that the production and care of a replacement clutch coincides with breeding season advancement and the concomitant deterioration of environmental and food conditions (reviewed by Moreno Citation1998; Verhulst & Nilsson Citation2008).
The fact that females desert their broods also at the end of the breeding season (late July and August) when it is too late to renest, suggests that females may derive benefits from desertion other than renesting opportunities. One obvious benefit for a deserter is to gain energy and improve the chances of survival, while another is to start moulting earlier or to set off on migration earlier (CitationSzékely et al. 1996; Lessells Citation2012). But these aspects with respect to Whiskered Tern still need to be studied. Female desertion in this species is not costly in terms of breeding success (Ledwoń & Neubauer Citation2017), so the benefits of desertion for females appear to outweigh the costs. It is obvious that by deserting, a female gains energy for her next breeding attempt, not necessarily in a given season with a new mate, but also for the next season with a permanent mate. Therefore, the male partner may benefit from female desertion if in the next season the same pair is very likely to breed together again (e.g. Gratto-Trevor Citation1991; Wojczulanis-Jakubas et al. Citation2020). The duration of pair bonds in Whiskered Tern should be studied. If mate fidelity in this species is high, it will be beneficial for males that the female does not mate with a new partner in a given season. The enhanced survival of their mates may be advantageous for males, as it probably results in the greater likelihood of a male obtaining a female in the following year.
In summary, renesting after desertion in the study population is not severely limited by the length of the breeding season. However, we found that deserting Whiskered Tern females did not appear to derive any major benefits by remating and renesting since the renesting rate after desertion is limited (c. 5%). In this long-lived species, females probably gained some other direct benefits, such as earlier moulting and/or earlier migration, or the indirect benefit that energy gains enhance survival.
CRediT authorship contribution statement
Mateusz Ledwoń: Conceptualization, data curation, formal analysis, investigation, methodology, funding acquisition, project administration, writing – original draft. Adam Flis, Agata Banach, Bartłomiej Kusal, Halszka Łożyńska, Nataly Atamas, Stanisław Broński, Jacek Betleja: Investigation, methodology, writing – review and editing. Agata Banach, Mateusz Ledwoń: Molecular sex determination.
Acknowledgements
We are grateful to anonymous reviewers for constructive suggestions on a previous draft of the manuscript. We greatly appreciate the cooperation of the fish farmers concerned; we are especially grateful to Jerzy Wojciech Adamek (Experimental Fish Farm in Zator). We thank Antonia Łobodzińska and Przemysław Kurek for their assistance in the field, Kasper Chlebowicz for his help in lab with molecular sex determination in 2015 and also Peter Senn for his linguistic guidance.
Disclosure statement
The study complied with current Polish laws. The fieldwork was carried out with the consent of the Local Ethical Committee and the Regional Directorate for Environmental Protection.
Additional information
Funding
References
- Amat JA, Fraga RM, Arroyo GM. 1999. Brood desertion and polygamous breeding in the Kentish Plover Charadrius alexandrinus. Ibis 141:596–607. DOI:10.1111/j.1474-919X.1999.tb07367.x.
- Amat JA, Visser GH, Pèrez-Hurtado A, Arroyo GM. 2000. Brood desertion by female shorebirds: A test of the differential parental capacity hypothesis on Kentish Plovers. Proceedings of the Royal Society B: Biological Sciences 267:2171–2176.
- Arnold TW, Devries JH, Howerter DW. 2010. Factors that affect renesting in Mallards (Anas platyrhynchos). The Auk 127:212–221. DOI:10.1525/auk.2009.09028.
- Banach A, Neubauer G, Flis A, Ledwoń M. 2021. Sex-specific growth of nestlings of the Whiskered Tern Chlidonias hybrida, a species with sexual size dimorphism and female brood desertion. Journal of Ornithology 162:1035–1047. DOI:10.1007/s10336-021-01911-y.
- Becker PH, Zhang H. 2010. Renesting of common terns Sterna hirundo in the life history perspective. Journal of Ornithology 152:213–225. DOI:10.1007/s10336-010-0639-0.
- Beissinger SR. 1986. Demography, environmental uncertainty, and the evolution of mate desertion in the Snail Kite. Ecology 67:1445–1459. DOI:10.2307/1939076.
- Beissinger SR. 1987. Anisogamy overcome: Female strategies in snail kites. American Naturalist 129:486–500. DOI:10.1086/284653.
- Beissinger SR. 1990. Experimental brood manipulations and the monoparental threshold in Snail Kites. American Naturalist 136:20–38. DOI:10.1086/285080.
- Beissinger SR, Snyder NF. 1987. Mate desertion in the snail kite. Animal Behaviour 35:477–487. DOI:10.1016/S0003-3472(87)80273-7.
- Betleja J 2003. Ecological conditions of the expansion of Whiskered Tern Chlidonias hybrida. Ph.D. dissertation, University of Wrocław, Poland (in Polish).
- Béziers P, Roulin A. 2016. Double brooding and offspring desertion in the barn owl (Tyto alba). Journal of Avian Biology 47:235–244. DOI:10.1111/jav.00800.
- Chambon R, Latraube F, Bretagnolle V, Paillisson JM. 2020. Sex-specific contributions to reproduction in Whiskered tern Chlidonias hybrida colonies of varying breeding density. Ardeola 67:113–125. DOI:10.13157/arla.67.1.2020.sc6.
- Clutton-Brock TH. 1991. The evolution of parental care. Princeton: Princeton University Press.
- Cockburn A. 2006. Prevalence of different modes of parental care in birds. Proceedings of the Royal Society B: Biological Sciences 273:1375–1383.
- Currie D, Valkama J, Berg Å, Boschert M, Norrdahl K, Hänninen M, Korpimäki E, Pöyri V, Hemminki O. 2001. Sex roles, parental effort and offspring desertion in the monogamous Eurasian Curlew Numenius arquata. Ibis 143:642–650. DOI:10.1111/j.1474-919X.2001.tb04892.x.
- Eberhart-Phillips LJ, Küpper C, Carmona-Isunza MC, Vincze O, Zefania S, Cruz-López M, Kosztolányi A, Miller TEX, Barta Z, Cuthill IC, Burke T, Székely T, Hoffman JI, Krüger O. 2018. Demographic causes of adult sex ratio variation and their consequences for parental cooperation. Nature Communications 9:1651. DOI: 10.1038/s41467-018-03833-5.
- Eberhart-Phillips LJ, Küpper C, Miller TE, Cruz-López M, Maher KH, Dos Remedios N, Stoffel MA, Hoffman JI, Krüger O, Székely T. 2017. Sex-specific early survival drives adult sex ratio bias in snowy plovers and impacts mating system and population growth. Proceedings of the National Academy of Sciences 114:E5474–E5481.
- Eldegard K, Sonerud GA. 2009. Female offspring desertion and male-only care increase with natural and experimental increase in food abundance. Proceedings of the Royal Society B: Biological Sciences 276:1713–1721.
- Ezaki Y. 1988. Mate desertion by male Great Reed Warblers Acrocephalus arundinaceus at the end of the breeding season. Ibis 130(4):427–437. DOI:10.1111/j.1474-919X.1988.tb01000.x.
- Franks S, Lank DB, Wilson WH. 2020. Western Sandpiper (Calidris mauri), version 1.0. In: Poole AF, editor. Birds of the World. Ithaca, NY, USA: Cornell Lab of Ornithology. DOI:10.2173/bow.wessan.01.
- Franz D, Theiss N, Graff H. 1987. Female of Penduline Tit (Remiz pendulinus) breeding successfully twice in one breeding-season at two places in a distance of more than 200 km. Journal of Ornithology 128:241–242. DOI:10.1007/BF01661698.
- Fujioka M. 1989. Mate and nest desertion in colonial little egrets. Auk 106:292–302.
- Gavin T, Székely T, Reynolds J. 2007. Sexual conflict and the evolution of breeding systems in Shorebirds. Advances in the Study of Behavior 37:271–342.
- Gochfeld M, Burger J, Kirwan GM, Garcia EFJ. 2020. Whiskered Tern (Chlidonias hybrida), version 1.0. In: Del Hoyo J, Elliott A, Sargatal J, Christie DA, de Juana E, editors. Birds of the World. Ithaca, NY, USA: Cornell Lab of Ornithology. DOI:10.2173/bow.whiter2.01.
- Goławski A, Kasprzykowski Z, Ledwoń M, Mróz E. 2016. Brood sex ratio in expansive and non-expansive tern species in east-central Poland. Bird Study 63:31–36. DOI:10.1080/00063657.2015.1122738.
- Gratto-Trevor CL. 1991. Parental care in Semipalmated Sandpipers Calidris pusilla: Brood desertion by females. Ibis 133:394–399. DOI:10.1111/j.1474-919X.1991.tb04587.x.
- Griggio M, Pilastro A. 2007. Sexual conflict over parental care in a species with female and male brood desertion. Animal Behaviour 74:779–785. DOI:10.1016/j.anbehav.2007.01.007.
- Gwiazda R, Ledwoń M. 2015. Sex-specific foraging behaviour of the Whiskered Tern Chlidonias hybrida during the breeding season. Ornis Fennica 92:15–22.
- Harrod W, Mumme R. 2021. Females compensate for moult‐associated male nest desertion in Hooded Warblers. Ibis 163:159–170. DOI:10.1111/ibi.12850.
- Hemborg C. 2004. Sexual differences in moult-breeding overlap and female reproductive costs in pied flycatchers, Ficedula hypoleuca. Journal of Animal Ecology 68:429–436. DOI:10.1046/j.1365-2656.1999.00295.x.
- Jamieson S. 2011. Pacific Dunlin Calidris alpina pacifica show a high propensity for second clutch production. Journal of Ornithology 152:1013–1021. DOI:10.1007/s10336-011-0691-4.
- Kenward RE. 2001. A manual of wildlife radio tagging. London: Academic Press.
- Kloskowski J. 2003. Offspring desertion in Red-Necked Grebes Podiceps grisegena. Ardea 91:25–34.
- Korpimäki E, Salo P, Valkama J. 2011. Sequential polyandry by brood desertion increases female fitness in a bird with obligatory bi-parental care. Behavioral Ecology and Sociobiology 65:1093–1102. DOI:10.1007/s00265-010-1118-6.
- Kosztolanyi A, Székely T, Cuthill I, Yılmaz K. 2006. Ecological constraints on breeding system evolution: The influence of habitat on brood desertion in Kentish plover. Journal of Animal Ecology 75:257–265. DOI:10.1111/j.1365-2656.2006.01049.x.
- Ledwoń M 2010. Male and female partitioning in parental care in Whiskered Tern Chlidonias hybrida. PhD Thesis, Polish Academy of Sciences, Kraków (in Polish).
- Ledwoń M. 2011. Sexual size dimorphism, assortative mating and sex identification in the Whiskered Tern Chlidonias hybrida. Ardea 99:191–198. DOI:10.5253/078.099.0209.
- Ledwoń M, Betleja J, Neubauer G. 2015. An effective method for trapping both parents and chicks in Whiskered Terns (Chlidonias hybrida) and its impact on breeding success. Waterbirds 3:290–295. DOI:10.1675/063.038.0309.
- Ledwoń M, Betleja J, Neubauer G. 2016. Different trapping schemes and variable disturbance intensity do not affect hatching success of Whiskered Terns Chlidonias hybrida. Bird Study 63:136–140. DOI:10.1080/00063657.2015.1136263.
- Ledwoń M, Neubauer G. 2017. Offspring desertion and parental care in the Whiskered Tern Chlidonias hybrida. Ibis 159:860–872.
- Ledwoń M, Neubauer G, Betleja J. 2013. Adult and pre-breeding survival estimates of the Whiskered Tern Chlidonias hybrida breeding in southern Poland. Journal of Ornithology 154:633–643. DOI:10.1007/s10336-012-0926-z.
- Ledwoń M, Neubauer G, Żmuda A, Flis A. 2019. Interaction between parent body condition and sex affects offspring desertion in response to acute stress. Journal of Ornithology 160:417–428. DOI:10.1007/s10336-019-01637-y.
- Ledwoń M, Szczys P. 2022. Extra-pair paternity in a species with frequent extra-pair courtship feedings, few extra-pair copulations, and male-biased parental care. Journal of Ornithology 163:437–444. DOI:10.1007/s10336-021-01954-1.
- Lessells CM. 2012. Sexual conflict. In: Royle NJ, Smiseth PT, Kölliker M, editors. The evolution of parental care. Oxford: University Press. pp. 150–170.
- McDonald GC, Cuthill IC, Székely T, Kosztolányi A. 2023. Remating opportunities and low costs underlie maternal desertion. Evolution 77:97–109. DOI:10.1093/evolut/qpac020.
- Minias P, Minias A, Dziadek J. 2014. Occurrence of extra-pair paternity and intraspecific brood parasitism in the Whiskered Tern Chlidonias hybrida. Bird Study 61:130–134. DOI:10.1080/00063657.2013.860949.
- Monaghan P, Nager RG. 1997. Why don’t birds lay more eggs? Trends in Ecology and Evolution 12:270–274. DOI:10.1016/S0169-5347(97)01094-X.
- Monaghan P, Nager RG, Houston DC. 1998. The price of eggs: Increased investment in egg production reduces the offspring rearing capacity of parents. Proceedings of the Royal Society B: Biological Sciences 265:1731–1735.
- Moreno J. 1998. The determination of seasonal declines in breeding success in seabirds. Etología 6:17–31.
- Morton ES, Stutchbury BJM, Chiver I. 2010. Parental conflict and brood desertion by females in blue-headed vireos. Behavioral Ecology and Sociobiology 64:947–954. DOI:10.1007/s00265-010-0910-7.
- Mumme R. 2018. The trade-off between molt and parental care in Hooded Warblers: Simultaneous rectrix molt and uniparental desertion of late-season young. The Auk 135:427–438. DOI:10.1642/AUK-17-240.1.
- Nisbet ICT. 2002. Common Tern (Sterna hirundo). In: Poole A, Gill F, editors. The Birds of North America, No. 618. Philadelphia: PA, The Birds of North America Inc. pp. 618. DOI:10.2173/tbna.
- Nisbet ICT, Mostello CS, Veit RR, Fox JW, Afanasyev V. 2011a. Migrations and winter quarters of five common terns tracked using geolocators. Waterbirds 34:32–39. DOI:10.1675/063.034.0104.
- Nisbet ICT, Szczys P, Mostello CS, Fox JW. 2011b. Female common terns Sterna hirundo start autumn migration earlier than males. Seabird 24:103–106.
- Osorno J, Székely T. 2004. Sexual conflict and parental care in magnificent frigatebirds: Full compensation by deserted females. Animal Behaviour 68:337–342. DOI:10.1016/j.anbehav.2003.06.027.
- Page GW, Warriner JS, Warriner JC, Paton PWC. 1995. Snowy Plover (Charadrius alexandrinus). In: Poole A, Gill F, editors. The Birds of North America: No. 154. Philadelphia & Washington, DC: The Academy of Natural Sciences and The American Ornithologists’ Union.
- Paillisson JM, Chambon R. 2021. Variation in male‐built nest volume with nesting‐support quality, colony, and egg production in whiskered terns. Ecology and Evolution 11:15585–15600. DOI: 10.1002/ece3.8162.
- Paillisson J-M, Latraube F, Reeber S. 2008. Assessing growth and age of Whiskered Tern Chlidonias hybrida chicks using biometrics. Ardea 96(2):271–277. DOI:10.5253/078.096.0212.
- Paillisson JM, Reeber S, Carpentier A, Marion L. 2007. Reproductive parameters in relation to food supply in the Whiskered Tern (Chlidonias hybrida). Journal of Ornithology 148:69–77. DOI:10.1007/s10336-006-0102-4.
- Parker GA, Royle NJ, Hartley IR. 2002. Intrafamiliar conflict and parental investment: A synthesis. Proceedings of the Royal Society B: Biological Sciences 357:295–307.
- Pilastro A, Biddau L, Marin G, Mingozzi T. 2001. Female brood desertion increases with the number of available mates in the rock sparrow. Journal of Avian Biology 32:68–72. DOI:10.1034/j.1600-048X.2001.320109.x.
- Rae R, Nicoll M, Summers RON, Stuart RAE, Brockie K. 2011. Breeding biology of Purple Sandpipers Calidris maritima on the Hardangervidda, southern Norway. Ornis Svecica 21:101–108. DOI:10.34080/os.v21.22606.
- Spina F. 1982. Contribution to the breeding biology of the Whiskered Tern Chlidonias hybrida in Val Campotto (Northern Italy). Avocetta 6:23–33.
- Stearns SC. 1992. The evolution of life histories. Oxford: Oxford University Press.
- Stenzel LE, Warriner JC, Warriner JS, Wilson KS, Bidstrup FC, Page GW. 1994. Long-distance breeding dispersal of Snowy Plovers in Western North America. The Journal of Animal Ecology 63:887–902. DOI:10.2307/5266.
- Székely T, Cuthill IC, Kis J. 1999. Brood desertion in Kentish plover: Sex differences in remating opportunities. Behavioral Ecology 10:185–190. DOI:10.1093/beheco/10.2.185.
- Székely T, Kosztolanyi A, Küpper C, Thomas GH. 2007. Sexual conflict over parental care: A case study of shorebirds. Journal of Ornithology 148:211–217. DOI:10.1007/s10336-007-0218-1.
- Székely T, Lessells CM. 1993. Mate change by Kentish Plovers Charadrius alexandrinus. Ornis Scandinavica 24:317–322. DOI:10.2307/3676794.
- Székely T, Reynolds JD. 1995. Evolutionary transitions in parental care of shorebirds. Proceedings of the Royal Society B: Biological Sciences 262:57–64.
- Székely T, Webb JN, Houston AI, McNamara JM.1996. An evolutionary approach to offspring desertion in birds. In: Nolan V Jr, Ketterson ED editors. Current ornithology. Vol. 6. New York: Springer. pp. 271–330.
- Székely T, Weissing FJ, Komdeur J. 2014. Adult sex ratio variation: Implications for breeding system evolution. Journal of Evolutionary Biology 27:1500–1512. DOI:10.1111/jeb.12415.
- Székely T, Williams TD. 1995. Costs and benefits of brood desertion in female Kentish plovers, Charadrius alexandrinus. Behavioral Ecology and Sociobiology 37:155–161. DOI:10.1007/BF00176712.
- Tavecchia G, Pradel R, Lebreton JD, Biddau L, Mingozzi T. 2002. Sex-biased survival and breeding dispersal probability in a patchy population of the rock sparrow Petronia petronia. Ibis 144:E79–E87. DOI:10.1046/j.1474-919X.2002.00059.x.
- Urano E. 1992. Early settling the following spring: A long-term benefit of mate desertion by male Great Reed Warblers Acrocephalus arundinaceus. Ibis 134:83–86. DOI:10.1111/j.1474-919X.1992.tb07235.x.
- Verhulst S, Nilsson J-A. 2008. The timing of birds’ breeding season: A review of experiments that manipulated timing of breeding. Philosophical Transactions of the Royal Society B: Biological Sciences 363:399–410. DOI:10.1098/rstb.2007.2146.
- Warriner JS, Warriner JC, Page GW, Stenzel LE. 1986. Mating system and reproductive success of a small population of polygamous snowy plovers. Wilson Bulletin 98:15–37.
- Webb JN, Houston AI, McNamara JM, Székely T. 1999. Multiple patterns of parental care. Animal Behaviour 58:983–993. DOI:10.1006/anbe.1999.1215.
- Wendeln H, Becker PH, Gonález-Solís J. 2000. Parental care of replacement clutches in common terns (Sterna hirundo). Behavioral Ecology and Sociobiology 47:382–392. DOI:10.1007/s002650050681.
- Williams GC. 1966. Natural selection, the costs of reproduction, and a refinement of Lack’s principle. American Naturalist 100:687–690. DOI:10.1086/282461.
- Wojczulanis-Jakubas K, Jakubas D. 2012. When and why does my mother leave me? The question of brood desertion in the Dovekie (Alle alle). Auk 129:632–637. DOI:10.1525/auk.2012.12095.
- Wojczulanis-Jakubas K, Jimenez Munoz M, Jakubas D, Kidawa D, Karnovsky N, Matechou E. 2020. Duration of female parental care and their survival in the little auk Alle alle – are these two traits linked? Behavioral Ecology and Sociobiology 74:1–11. DOI:10.1007/s00265-020-02862-9.
- Zárybnická M. 2009. Parental investment of female Tengmalm’s Owls Aegolius funereus: Correlation with varying food abundance and reproductive success. Acta Ornithologica 44:81–88. DOI:10.3161/000164509X464911.