
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Livestock production in the Colombian Amazonia is typically dual-purpose (meat and milk) based on silvopastoral systems. We aimed to assess how the ecological condition of these systems correspond to the ecosystem services they generate based on an assessment of farmers’ knowledge of the tree species. We evaluated 159 paddocks. We recorded 143 tree species from 37 families, with 22 species being most common (relative occurrence frequency = 57%). Based on tree species composition, we characterized four states of silvopastoral condition (hereon ‘paddock condition types’) using clustering techniques: (i) High structural complexity and highest tree species richness (HSCR); (ii) High species diversity and tall trees (HDTT); (iii) High species diversity and medium-sized trees (HDMT); (iv) Structurally simple with lowest species richness (SSLR). Tree species richness was significantly higher in HSCR (11.70 ± 1.47 per paddock), than in SSLR (2.86 ± 0.80). HDTT and HDMT had similar richness, with intermediate values (5.55 ± 0.82 and 6.38 ± 0.51, respectively). Farmers appreciate a number of ecosystem services provided by the silvopastoral system, but a limited number of tree species are valued. This indicates the need for additional biodiversity conservation measures in these landscapes, including measures to improve knowledge about the value of tree species with few occurrences or low densities.
EDITED BY:
Introduction
Deforestation in the Colombian Amazon region has increased in recent years. In 2016, the deforested area was 70,074 ha, doubling by 2017 (144,147 ha). In the same year, the department of Caquetá in the Colombian Amazon reported a deforested area of 60,373 ha (IDEAM Citation2017). The causes of deforestation in this region are mainly wood extraction, cropping, mining, and extensive livestock management practices (Staal et al. Citation2020; Dávalos et al. Citation2021).
Livestock rearing is therefore a major driver of land transformation and use patterns in the Colombian Amazon region. Processes of occupation, and transformation of natural landscapes have resulted in extensive reduction of natural forest areas (Fonseca Citation2009), with negative impacts on biodiversity and key ecosystem services (ES) provided by forest ecosystems including fodder, wood, water protection, shade, nutrient inputs, water flow regulation and carbon storage (Armenteras et al. Citation2019). Besides, the quest for higher grass production has led to a decrease in the tree component in the landscape matrix. These changes also have consequences in terms of the ES generated by trees in the silvopastoral systems that support livestock production. Also, the lack of tree cover has negative impacts on animal production, affecting the sustainability of the production system in tropical regions (Villacis et al. Citation2003; Villanueva et al. Citation2003; Álvarez et al. Citation2021a).
Some livestock production in the Colombian Amazon is supported by silvopastoral systems with dispersed or grouped trees (Sotelo et al. Citation2017). They are the result of natural regeneration and/or are remnants of the original forest vegetation, hence, these tropical silvopastoral systems are often considered to contribute to the conservation of native biodiversity at the same time that they have a higher capacity to deliver multiple benefits (like thermal regulation, enhanced nutrient and carbon stocks, wood and fodder production (Pagiola et al. Citation2007; Casals et al. Citation2014) compared to treeless pastures grown after clear-cutting (Jose and Dollinger Citation2019). Tree species richness, abundance and diversity in silvopastoral systems are largely affected by the impacts of livestock grazing and browsing (Esquivel et al. Citation2008), but also by the active selection of trees by the farmers (Barton et al. Citation2016). The management and selection of scattered trees in livestock systems have been related to farmers’ local knowledge on tree ES (Harvey et al. Citation2011; Suárez et al. Citation2013; Sánchez et al. Citation2017). Important provisioning ES generated by these systems are fodder, firewood and material for e.g. construction (Harvey and Haber Citation1999; Cajas-Giron Citation2001; Pascual et al. Citation2010; Sánchez et al. Citation2017). In addition, trees in pastures are important for regulating services including microclimate regulation and the enhancement of carbon and nutrient stocks (Esquivel et al. Citation2003; López et al. Citation2004; Ruiz et al. Citation2005; Harvey et al. Citation2006; Manning et al. Citation2006; Esquivel et al. Citation2008; Suárez et al. Citation2013; Casals et al. Citation2014; Hoosbeek et al. Citation2018). Farmers also select and maintain trees to modify the microclimate of grazing areas to improve animal welfare (Blackshaw Citation1994), which in turn, has a positive effect on the livestock production (Villacis et al. Citation2003; Villanueva et al. Citation2003; Álvarez et al. Citation2021a). Tree cover is also likely to affect macroclimatic processes such as ‘flying rivers’ (Pearce Citation2019). Local knowledge on the ES generated by individual tree species is likely critical for maintaining native tree species diversity, hence playing a role in biodiversity conservation in livestock production systems, and often increasing the level of connectivity among forest remnants in landscape (Guevara et al. Citation1998; Fischer and Lindenmayer Citation2002; Harvey et al. Citation2006; Medina et al. Citation2007; Gutiérrez et al. Citation2012).
The generation of ES depends on the structure and function of natural and managed ecosystems (e.g. Grondard et al. Citation2021). There are many studies on the composition and structure of trees in silvopastoral systems reporting on the variability in tree density and species composition in the tropical America (Cajas-Giron Citation2001; Sánchez et al. Citation2005; Griscom et al. Citation2011; Harvey et al. Citation2011; Lombo et al. Citation2011), in Europe (Hartel et al. Citation2017, Citation2018) and Asia (Hosseininasab et al. Citation2017). However, a structured and quantitative assessment of the linkages between ecosystem characteristics (ecosystem condition sensu United Nations (Citation2021)), and the capacity to generate ecosystem services is lacking. In this study, using an example of the silvopastoral systems in the Colombian Amazon foothills, and aligned with the principles of natural capital accounts (United Nations Citation2021), we aimed to understand how the different structures of silvopastoral paddocks in terms of tree cover, diversity and density (i.e. the ecological condition of the silvopastoral system), correspond to the delivery of ES as defined by the uses and ecological knowledge of the farmers who manage these systems. Our assumption is that systems with higher tree diversity have the capacity to generate multiple ecosystem services. Our hypothesis was, then, that silvopastoral paddocks with high tree diversity and density can generate more ES than paddocks with more simple structure.
Specifically, we aimed to i) characterize the silvopastoral systems condition in terms of their tree species abundance, composition, and structure; ii) assess the farmers’ knowledge and appreciation on the ES provided by tree species in these silvopastoral systems; iii) assess how tree species composition and structure in these systems correspond to the generation of ES; and iv) to evaluate the role of these systems in Neotropical tree species conservation.
For that purpose, we first identified paddock types based on tree species composition, diversity, and structure in a set of silvopastoral paddocks. Then, we assessed the main ES provided by trees in the study region based on farmers’ knowledge and uses, and thirdly, we related the silvopastoral paddock types to the set ES provided by trees within these paddocks. We finally discuss the interactions between farmers’ knowledge and paddock types.
Materials and methods
Study area
The department of Caquetá is in the upper basin of the continental Amazon and has an area of 8,895,600 ha. In this region, and especially in the foothills of the Colombian Amazon, deforestation has occurred mainly due to livestock production activities (dual-purpose i.e. meat and milk production) (Ramírez et al. Citation2012; Suárez et al. Citation2013). This system has a population of 2 175 065 cattle, making it the fifth largest cattle stock in Colombia and the fifth largest milk production basin in the country (1 873 482 kg day−1) (Torrijos Citation2022). This production is supported by an area of 1 640 171 ha of pasture (Murcia et al. Citation2014).
The study was conducted on selected farms located in the municipalities of Belén de los Andaquíes, Morelia, Florencia, and El Doncello in the Amazon foothills of the department of Caquetá. These farms are located within the Tropical Humid Forest zone defined in Holdridge (Citation1978), with rainfall of 3 600 mm yr−1, mean temperature of 28.5°C and relative humidity of 84% (IGAC Citation1993). The soils correspond to entisols, inceptisols, ultisols and oxisols which are soils with low fertility and high saturation of aluminum and iron (IGAC Citation2014).
Farm selection
This study is part of a larger project that aimed at evaluating the role of tree cover in silvopastoral systems for dual-purpose livestock production in the Colombian Amazon. The project involved 60 farms representative of the livestock production systems in the study area, selected through directed sampling (Glaser et al. Citation1968) tree to the following selection criteria: (i) the farms had a maximum distance of 80 km between each other, and a minimum of 3 km; (ii) the farmer was willing to collaborate; (iii) the farmer resided on the farm or the farm had regular visits from the resident administrator or manager; (iv) the availability of farm technical records: information on the productive and administrative data of the farm; (v) year-round access to the farm by the group of farmers, technicians and researchers; (vi) the presence of a minimum of investment in physical infrastructure to manage the cattle, indicating a managed system; (vii) the willingness of the farmers to benefit from the results obtained from the project; (viii) good accessibility to the farm (road infrastructure in good condition) and access to basic public services (electricity, drinking water). From the 60 farmers/farms described above, we randomly selected: 48 to evaluate local knowledge on tree ES and 12 to characterize tree structure and composition ().
Table 1. Main characteristics of the 12 farms selected for paddock composition and structure sampling.
A. Characterization of paddock tree composition and structure
Paddock composition
In the selected 12 farms, we identified 159 silvopastoral paddocks, with an average area of 5.78 ha ±0.09 per paddock. Paddock area ranged from 0.11 to 20.69 ha. The size of the sampled area per farm ranged from 2.7 to 107.8 ha (), giving a total sampled area of 430.8 ha. In each paddock, all the trees with a circumference at breast height (CBH) greater than 10 cm were inventoried. Species identification was conducted at the herbarium of the Universidad de la Amazonia, with material collected in the field. We recorded tree diameter at breast height (DBH), commercial height (height of the trunk under the first branch), total height, the largest diameter of the tree crown, and the average crown diameter calculated based on diameters measured at 45°, 90° and 135° with the largest diameter direction as a reference ().
Table 2. Calculation of paddock structural characteristics.
Relative importance of tree species in paddock composition
To assess the relative importance of each tree species in the paddocks, we computed their Importance Value Index (IVI) (Curtis and Mcintosh Citation1950). The calculation of IVI was based on the species’ relative abundance RA (%), relative dominance RD (%) and relative frequency RF (%) according to Formula 1:
where,
and where IVIi is the Importance Value Index for species i, Ni is the number of individual trees of species i, N the total number of individuals, BAi is the basal area of species i, BAT is the basal area summed across all the species, NPi is the number of paddocks where species i is present, NPT the total number paddocks.
Paddock tree species composition and diversity
We characterized tree composition and diversity in each paddock in terms of i) species richness, ii) species abundance (trees ha−1), iii) Shannon index, and iv) Simpson index. Since paddocks varied in size, calculations of species richness and diversity indices were based on expected number of species using species rarefaction curves. With this analysis, the expected number of species per hectare, the percentage of expected species and the sampling effort necessary to reach the expected value were estimated by adjusting the Clench´s species rarefaction model (Soberón Citation1993). All indices were calculated using Qeco-Quantitative ecology software (Di Rienzo et al. Citation2010) considering differences in paddock area.
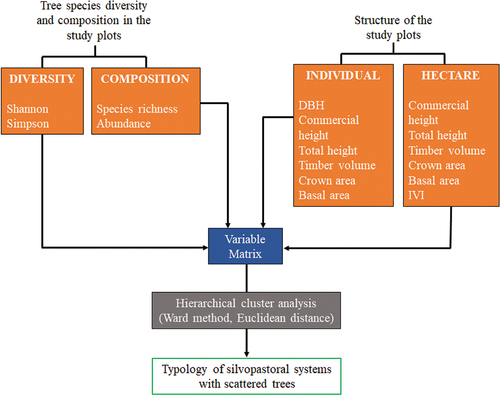
Typology of paddock systems based on tree composition and structural characteristics
We built a typology of silvopastoral paddock condition in the region based on the following variables: species richness, abundance, Shannon diversity, Simpson diversity, diameter at breast height, commercial height, total height, timber volume per individual, timber volume per hectare, canopy area per tree, basal area per individual, basal area per hectare, tree density, canopy area per hectare, tree cover, and importance value index (). Since dominant species largely determine key ecosystem functions (Grime Citation1998), which are in turn associated with the capacity to generate ES, we quantified the level of dominance and relative abundance of individual tree species by computing 80% of the cumulative relative tree frequency within each paddock.
Using the 16 variables listed above (), we carried out a hierarchical cluster analysis (Ward method, Euclidean distance) to define paddock types (). We used the gDGC test to identify multivariate statistical differences between clusters (Valdano and Di Rienzo Citation2007, α = 0.05). At a second stage, we used linear mixed models (LMM) to assess differences in the composition and structure of trees among the paddock types (fixed effect) including farm effect as random factor to consider differences among farms (Casanoves et al. Citation2005). We applied the Fisher LSD test for mean differences (α = 0.05). To explore the main factors that determined differences among clusters, we performed a Principal Component Analysis (PCA) using the InfoStat program (Di Rienzo et al. Citation2018) and R 3.4.4 software (R Core Development Team Citation2018), using the Ade 4.
Figure 1. Conceptual model of paddock system typology based on tree composition, diversity, and structure.
B. Perceived ecosystem services provided by tree species in the study livestock systems surveys
To understand and analyze the different ES attributed to trees in livestock systems in the region and the level of appreciation of these benefits, a three-step process was followed with the 48 livestock farmers that participated in the study.
We collected information on the goods and services provided by trees in the livestock systems by conducting a semi-structured survey. The survey (Supplementary 1) was applied to each farmer without sequence or order, allowing the informants to speak freely and refer to their experiences with the tree resources, but with a focus on the interactions of the trees with the soil and the pastures (regulating services), as well as the uses, the effects of the tree component with the other elements of the farm, and specific benefits obtained from the trees.
The interviews were transcribed without interpretation (in the producer’s own words) and from each interview unit, statements were extracted and recognized as opinions on some specific topic (short sentences that express a clear and precise idea) (Dixon et al. Citation2001).
With this information, we built a database of the ES identified by the farmers as generated by the trees in the livestock production systems in the region.
According to the identified ES (livestock feed, timber production, water protection, shade for livestock and nutrient inputs to the soil), the average number of ES provided by trees per paddock was calculated. The level of ES provision per paddock was estimated quantitatively by multiplying the ES generated by each tree species times the number of individuals of the species in each paddock.
Farmers’ valuation of tree species and ES
To assess quantitatively the value attached to each species, and indirectly to the ES it generates, we computed the index of cultural importance (ICI) for each species following the methodology proposed by Sánchez et al. (Citation2017):
where Iu (Intensity of Use) represents, the number of ES attributed to a given species, Fm is the Frequency by which each species was mentioned by the informants, i.e. the sum of the number of times a species was referred to in the interviews, considering all uses and all informants, Vux (the Use value) is the proportion of times in which each species was associated with a given ES. The sum of the ICI across tree species was calculated and divided by the paddock total area, and therefore converted to an ICI per hectare.
C. Relationship between the relative importance of the tree species in the paddocks and the knowledge assessment of tree ecosystem services in the study paddocks
To assess the relationship between the biophysical importance of each tree species in the paddocks (IVI) and the value attached to the species (ICI), a Spearman correlation analysis was performed between area-based ICI and IVI. We then analyzed how ICI and IVI varied across paddock types, using an ANOVA.
The data generated in the study have been published at https://osf.io/uyr43
Results
Tree species composition and diversity in the study paddocks
A total of 4 657 trees were surveyed in 430.8 ha, in 159 paddocks with an average area per paddock of 5.78 ± 0.09 ha. We found 143 species of trees belonging to 37 families. The species Bellucia pentamera (Pomo) showed the highest values for RA (12.83%) and RF (6.65%), as well as IVI (26.08%) and Aegiphila integrifolia (Tabaquillo, included in the category ‘other species’) presented the lowest values for Relative abundance (RA) (0.01%), Relative dominance (RD) (0.00%), Relative Frequency (RF) (0.11%) and Importance Value Index (IVI) (0.12) (). Gmelina arborea (Melina) had the highest RD (11.86%). B. pentamera, Psidium guajava (Guayabo) and G. arborea were the three tree species with a highest IVI (26.08; 21.13; 19.59 respectively). The two species with highest IVI are fruit-producing species, which are important for cattle feeding ().
Table 3. Importance Value Index (IVI) computed for the tree species found in the paddocks, based on relative abundance RA (%), relative dominance RD (%) and relative frequency RF (%).
Typology of paddocks based on tree species composition and diversity structure
The cluster analysis based on 16 variables of tree species composition and diversity allowed to determine four paddock types () that we defined as: i) High structural complexity and highest tree species richness (HSCR); ii) High species diversity and tall trees (HDTT); iii) High species diversity and medium-sized trees (HDMT); and iv) Structurally simple with lowest species richness (SSLR).
Figure 2. Sample-size-based rarefaction (solid lines). The 95% confidence intervals (gray-shaded regions) were obtained by a bootstrap method based on 1000 replications. The expected coverage for the sampling effort in the study paddocks was 99%.
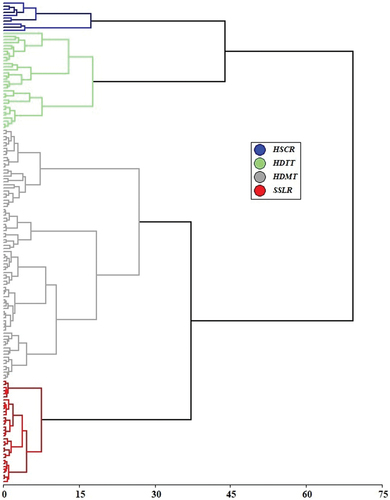
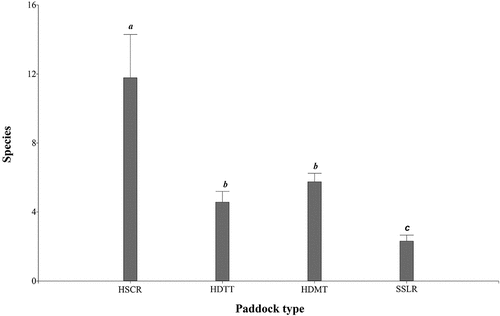
Paddock types differed significantly in species richness and number of trees (p < 0.0001) (), but we found no differences in species diversity estimated by the Shannon and Simpson indices. Regarding structural variables, we found highly significant differences among paddock condition states in terms of mean DBH, commercial height, total height, timber volume per individual and per hectare, canopy area per individual and per hectare, basal area per individual and per hectare, tree cover and IVI (p < 0.0001) (). A detailed description of the paddock types is provided below.
Table 4. Tree species composing 80% of cumulative abundance in each silvopastoral typology in livestock farms under the dual-purpose system in the humid Amazonian tropic of Colombia.
Table 5. Description of identified types, according to the used variables of composition, diversity, and structure.
High structural complexity and highest tree species richness (HSCR)
This paddock type represented 6.8% of the total number of paddocks (n = 10). It is characterized by the highest tree species richness (11.70 ± 2.53) and the highest tree abundance per paddock (87.70 ± 33.81) (). In terms of structure, this paddock type has the highest number of individuals per hectare (40.15 ± 2.66 ind ha−1), the highest timber volume (7.67 ± 0.65 m3 ha−1), the largest basal area (2.22 ± 0.16 m2 ha−1), the largest canopy area per ha (2725.44 ± 134.66 m2 ha−1), and the highest percentage of tree cover (27.25 ± 1.35%). These variables indicate high complexity of tree composition, which was represented by the highest IVI (3.32 ± 0.37) (). Thirteen tree species composed 80% of the cumulative relative frequency, including Zygia longifolia (Carbón), Guarea Guidonia (Bili bil) Psidium guajava, Bellucia pentamera and Ficus donnell (Higerón) ().
High species diversity and tall trees (HDTT)
This paddock type represented 20% of the total number of paddocks (n = 32). It is characterized by intermediate tree species richness (5.56 ± 0.74), and big trees with mean DBH = 33.48 ± 1.43, total height = 9.59 ± 0.40 m, basal area = 0.12 ± 0.01 m2 ind−1. This type had the largest timber volume per individual (0.46 ± 0.05 m3). The commercial height (3.44 ± 0.21 m), the canopy area per tree (79.96 ± 6.23 m2 ind−1) and IVI (2.77 ± 0.22), were not significantly different from HSCR paddocks (). Fourteen tree species composed 80% cumulative relative frequency including Zygia longifolia, Psidium guajava, Andira inermis (Cobre), Gmelina arborea, Ficus yoponensis ().
High species diversity and medium-sized trees (HDMT)
Paddock type HDMT had the largest number of paddocks (n = 83), representing 52.2% of the total number of study paddocks. Similarly, to HDTT, this type was characterized by intermediate tree species richness 6.36 ± 0.51 (). Their structural variables DBH (20.58 ± 1.15), total height (6.97 ± 0.36 m), timber volume (0.11 ± 0.03 m3 ind−1) and basal area (0.04 ± 0.01 m2 ind−1) per individual were similar to those in HSCR; while the number of individuals per hectare (10.15 ± 1.24 ind ha−1), tree canopy (474.82 ± 48.1 m2 ha−1) and the percentage of tree cover (4.75 ± 0.48%) was similar to HDTT paddocks (). However, the commercial height, the canopy area and the IVI in this group were significantly lower compared to HSCR and HDTT. Twenty tree species made up 80% of the cumulative relative frequency, such as Bellucia pentamera, Psidium guajava, Sapium glandulosum, Gmelina arborea and Ocotea longifolia ().
Structurally simple with lowest species richness (SSLR)
SSLR represented 21% of the total number of paddocks (n = 34). It is characterized by the lowest tree species richness (2.85 ± 0.42) and abundance (). Regarding structural variables, this group also presents the lowest means, except for the number of individuals per hectare (6.67 ± 1.64 ind ha−1), which was similar to values of HDTT (). These types present the highest skewness in dominance, with only six tree species accounting for 80% cumulative relative frequency, including Sapium glandulosum, Psidium guajava, Bellucia pentamera, Miconia minutiflora, Astrocaryum murumuru ().
The different types of paddocks were represented in a biplot constructed from a principal components analysis to show the ordering of the clustering and the variables most associated with each group ().
Figure 3. Biplot constructed with structure and diversity variables: Species richness (Sp), Abundance (Ab), Shannon diversity (Sd), Simpson diversity (Simd), Diameter at Breast Height (DBH), Commercial height (Ch), Total height (Th), Timber volume per individual (TvInd) Timber volume per ha (TvHa), Canopy area per tree (CaInd), Basal area per individual (BaInd) and Basal area per hectare (BaHa), Tree density (IndHa), Canopy area per hectare (CaHa), Tree cover (TC), Importance Value Index (IVI). The plot shows the four paddock condition types identified: HSCR: High structural complexity and highest tree species richness, HDTT: High species diversity and tall trees, HDMT: High species diversity and medium-sized trees and SSLR: Structurally simple with lowest species richness.
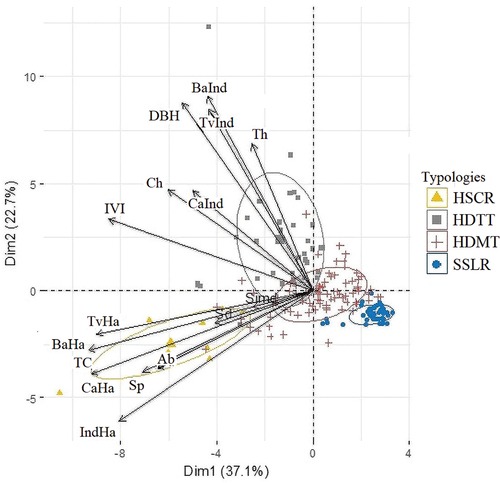
The variables most associated with the HSCR type were species richness and abundance, and the Shannon and Simpson indices (); likewise, this type presented the highest number of trees per ha, canopy area, tree cover, basal area, and timber volume. The HDTT type was associated with high individual basal area, canopy area and timber volume, indicating the dominance of large, tall trees, with trees with the highest total and commercial height and DBH. The HDMT and SSLR types were not associated with any variable (). Tree species richness varied between paddock types, indicating differences among them in terms of the biodiversity conservation value. The HSCR type was characterized by the highest tree species richness, HDTT and HDMT types, presented intermediate species richness values and SSLR type were associated to the lowest values ().
Figure 4. Average number of tree species richness by paddock condition type.
Ecosystem services provided by trees in livestock systems
Our work also aimed at characterizing the provision of ES by the trees in dual-purpose livestock systems in the Amazonian foothills of the department of Caquetá, Colombia. The main ES identified by the farmers were provisioning services (i.e. livestock feed and timber) and regulating services (i.e. protection of water sources, shade for livestock and maintenance of soil fertility). In this study, 22 species with the highest IVI had a cumulative relative frequency of 57% of the total tree species surveyed. Six of these species did not provide any ecosystem services for the livestock system recognized by the interviewed farmers: Guarea guidonia, Vitex orinocensis (Flor morado), Hieronyma rufa (Chuguacá), Mangifera indica (Mango), Myrcia splendens (Arrayán), Piptocoma discolor (Boca indio). Sixteen species generated between four and one ES valued by the farmers ().
Table 6. Provision of goods and ES recognized by 48 interviewed farmers for the main tree species (ordered according to their relative frequency) in the study livestock farms.
Ecosystem services provided by paddock types
The number of trees per hectare providing a given ES was calculated in the different paddock types. Based on estimates of tree density, HSCR paddocks generate multiple ES including, livestock feed, wood production, water protection and shade for cattle, with 10.80 ± 1.57, 18.41 ± 1.79, 5.71 ± 0.51 and 15.10 ± 1.45 trees ha−1, respectively. We found no significant differences among paddock types in terms of nutrient inputs/maintenance of soil fertility. (p = 0.1047) ().
Table 7. ES weight average per hectare based on average number of trees and basal area of the tree species occurring in the paddock type.
Based on basal area estimates, HSCR paddocks presented highly significant differences (p < 0.0001) compared to other paddock types for provisioning ES: feed for livestock, wood production and shade for livestock. Regarding regulating ES (protection of water sources), HSCR paddocks presented higher levels (p = 0.0005) than the other paddock types (DTT, DMT and LR). Also in this case, there were no significant differences in the ES soil nutrient inputs among types ().
The index of Cultural Importance (ICI) across paddock types
Amongst the 143 tree species present in the paddock systems, only 22 species had an ICI according to the farmers. The average ICI values per hectare showed differences between the different paddock types (p = 0.0002). HSCR paddocks had the highest average ICI per hectare (37.52 ± 7.48). We did not find significant differences between the LR and DMT types, but the average ICI values per hectare of these two types were higher than that of the DTT type ().
Figure 5. Relationship between the Index of Cultural Importance (ICI) and the Importance Value Index (IVI) in the four paddock condition types (HSCR= High structural complexity and highest tree species richness; HDTT= High species diversity and tall trees; HDMT= High species diversity and medium-sized trees; SSLR= Structurally simple with lowest species richness. Different letters indicate differences among means for IVI (capital letters) and for ICI (small letters) at p< 0.05.
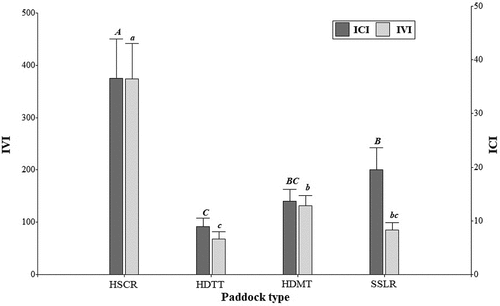
Relationship between the Index of Cultural Importance (ICI) and the Importance Value Index (IVI) in the identified types
The value of the Spearman’s correlation coefficient between the IVI and the ICI was r = 0.53 (n = 31, p = 0.0021). The IVI average values per hectare showed significant differences between the different paddock types (p < 0.0001). HSCR paddocks had the highest average IVI per hectare. We did not find significant differences between DMT and LR types, nor between LR and DTT types but the average IVI per hectare of the DTM type was higher than the value of the DTT type ().
Discussion
Tree species conservation in the study livestock systems
The structure of ecosystems, in our case, agro-pastoral ecosystems, given by the way that biodiversity is integrated and managed in these systems, determines their condition and their capacity to deliver ecosystem services (United Nations Citation2021). This study presents, to our knowledge, the first characterization of the main silvopastoral systems in the Colombian Amazon, based on the tree composition, diversity, and structure in 159 paddocks set up in the region of Caquetá, and linked to a quantitative assessment of their capacity to generate ecosystem services and of the value attached to these services.
According to Rodríguez-León et al. (Citation2022), the landscape of Caquetá is home to almost 1000 species of trees and shrubs. In a total sampled area of 430.8 ha, we found 143 tree species of 37 families, indicating the role of these systems in maintaining the diversity of trees in the region. This is related to many important functions provided by neotropical trees in production systems: i.e. regulating temperature, providing shade for livestock, retaining soil water, improving and maintaining soil fertility (Jose and Dollinger Citation2019). The diversity of this silvo-pastoral system appears to be lower than similar silvo-pastoral systems studied in Nicaragua, by Esquivel et al. (Citation2009) and Sánchez et al. (Citation2005), who reported 85 species in 46 ha and 180 species in 21.4 ha, respectively. Other references to the area include Villanueva-Partida et al. (Citation2016) who report 64 species in 64 ha of trees dispersed in pastures in southern Tabasco, Mexico; Esquivel et al. (Citation2003), 99 woody perennial species in 836 ha; and Grande et al. (Citation2010), 75 tree species belonging to 31 families in 41.7 ha in livestock systems in Sierra Tabasco. Further, compared to regions within Colombia, the silvopastoral systems in the Caquetá region appear to offer more opportunities for the conservation of native biodiversity than, for instance, the Caribbean microregions (Litoral, Golfo de Morrosquillo, Sabanas and Valle del Sinu), for which Cajas-Giron (Citation2001) reported a total of 96 tree species in 54 silvopastoral paddocks with three size classes (<50; 50 to 100; and >100 ha). Regional differences in terms of number of tree species reflect both different environmental factors and ecological conditions (Harvey et al. Citation2011), habitats (Sánchez et al. Citation2005) and climatic characteristics (temperature, rainfall, solar radiation), in addition to farmers’ preferences, their knowledge about the importance of trees in livestock production system, and the choices they make about which species to maintain and/or conserve in silvopastoral paddocks (Cáceres et al. Citation2015; Barton et al. Citation2016).
Tree species composition of livestock systems
The features of the silvopastoral paddocks we identified are likely associated with different factors shaping the livestock systems in the region, including the capability of tree species to cope with anthropogenic changes in the soil and micro-climate, i.e. an environment with higher temperatures and more light than in the close-canopy forest that develops under the same ecological conditions (Aitken et al. Citation2008). Further, land-use changes and the history of use and management (Wassie et al. Citation2009) lead to differences in colonization rates of tree species in the silvopastoral landscape, determining tree species composition and richness of paddocks tropical silvopastoral systems (Medina et al. Citation2004; Vílchez et al. Citation2004). These changes are caused by shifts in the local climate, but also through differences in seed dispersal by wildlife (birds, monkeys) (Wunderle Citation1997; Messeder et al. Citation2020) and cattle (Harvey et al. Citation2011; Pignataro et al. Citation2017), and by wind (Pignataro et al. Citation2017). For instance, B. pentamera and P. guajava are species with a high natural regeneration capacity and early-succession colonization traits in livestock systems in the Colombian Amazon. In our study, these two species showed high abundance and had the highest IVI compared to the other tree species ().
The condition of tropical silvopastoral systems also depends on the choices the farmer makes to maintain particular species at the time of clearing the forest, which relies on the farmer’s knowledge of the attributes and functions of the trees in the livestock systems (Barton et al. Citation2016), but which is further constrained by the characteristics of the forest before changing the land use from forest to silvopastoral system. In the Caquetá region, secondary forest, which is widely dominated by light demanding species, is common. We found species that are likely well adapted to regenerate in the open canopy of silvopastoral systems. Harvey et al. (Citation2011) and Esquivel et al. (Citation2009) found that trees can regenerate naturally in silvopastoral system and identified species with active regeneration (the case of B. pentamera and P. guajava in this investigation) and those with limited natural regeneration. These authors report that P. guajava, G. ulmifolia, and E. cyclocarpum showed high abundance in paddocks in Nicaragua and Costa Rica (Harvey et al. Citation2011), but Esquivel et al. (Citation2009) report that P. guajava only had a high regenerative capacity in the juvenile stage. Trampling and grazing by livestock is also a critical factor in determining regeneration rates in silvopastoral systems (Esquivel et al. Citation2009; Rusch et al. Citation2017). Hence, forest succession and farmers knowledge both determine the composition and the structure currently found in the paddocks in this study: i.e. large, low density, and tall trees, with few dominant species; or medium size trees with a large number of species.
Local knowledge of farmers and its impact on tree species conservation
The knowledge of the farmer and how this affects the management of the silvopastoral farm depends on the type of livestock systems and farmers’ needs (Esquivel et al. Citation2003; Sánchez et al. Citation2005; Esquivel et al. Citation2008; Wassie et al. Citation2009; Harvey et al. Citation2011; Suárez et al. Citation2013; Barton et al. Citation2016; Sánchez et al. Citation2017). Farmers, with their knowledge of trees and on the ES they generate (forage, fruit, shade, timber, and firewood) can manage the natural regeneration of preferred trees by protecting saplings and adjusting the intensity of cattle grazing (Cajas-Giron Citation2001). Also, some indigenous communities have developed silvopastoral systems based on their traditional knowledge of the local natural resource (Pignataro et al. Citation2016). Management practices such as weeding in paddocks with the use of herbicides or manual methods also affect the composition of the trees in the paddocks (Camargo et al. Citation2000; Harvey et al. Citation2011). All these management practices affect paddock tree structure, composition, and diversity, as we found in our study.
The tree species composition we found in the study paddocks is coherent with the knowledge the farmers have of the trees. The selective preservation of some trees is related to the ES they provide in the farms (livestock feed, timber provision, water protection, shade for livestock and nutrient inputs to the soil). Also, Suárez et al. (Citation2013) determined 15 perennial woody species that provide shade to the animals in the same livestock production system in the Colombian Amazon. Shade reduces heat stress, showed by lower rectal temperatures and respiratory rates, and higher production under the trees shade (Valtorta et al. Citation1997), but see also Ainsworth et al. (Citation2012) for conflicting results. Farmers appear to have a clear understanding of this function by trees, and often qualify the trees according to its properties to provide a benign climate in the grazing areas for the animals. Further, Barton et al. (Citation2016) demonstrated how the identification of a multifunctional tree portfolios can meet a desired and farmer-prioritized ecosystem services profile, seeking to ensure that ecosystem services improve provisioning and regulating services in the livestock system. Local knowledge about the value generated by the tree cover therefore seems to help maintain many species in livestock systems, thus contributing to protect a good number of species in the Colombian Amazon.
However, amongst the 143 species we found in the study systems, only 22 were ubiquitous (cumulative relative frequency = 57%). The 22 species were typical of secondary forest succession. From these 22 tree species, 16 were recognized as producing ES relevant for animal production. Further, managing species diverse systems can considerably increase the demand for labor, which can be a severe constraint in silvopastoral systems worldwide (Barton et al. Citation2017; Nyberg et al. Citation2020). Harvey et al. (Citation2011) suggested that the recognition by farmers of the provision of ecosystem goods and services from trees leads to an increase in the number of trees in grazing areas, but on the other hand may limit the tree composition to a few favored species. Hence, this dependence on few tree species and their ES in silvopastoral systems is not sufficient to protect biodiversity in the livestock production systems in the Colombian Amazon, especially for species depending on primary forest tree species and habitats (Manning et al. Citation2006). It further highlights the need for strategies that combine set asides of patches of primary forest embedded in biodiversity-friendly production landscapes to achieve the conservation and sustainable use of tropical forest biodiversity (IPBES Citation2019).
On the other hand, some species appear to persist in the livestock production system despite not being specifically valued by the farmers. For instance, G. guidonia, V. orinocensis, C. latifolia, H. rufa, M. splandens and P. discolor were not recognized by the farmers as important for the generation of ES, but this group of tree species are early-succession colonizers and are dispersed by cattle, birds or wind and have high reproductive capacities (Stevens et al. Citation2001). These characteristics favor the regeneration of these tree species, allowing them to successfully overcome low seed availability, high solar radiation, high evapotranspiration, degraded soils, grazing and cattle trampling (Nepstad et al. Citation1996; Holl Citation1999).
The sustainability of the paddocks and the livestock system in the Amazon region will be achieved by increasing and conserving the diversity of trees in the grazing areas. It has been reported (Esquivel et al. Citation2008; Rusch et al. Citation2017) that there are limitations to recruitment of new trees in silvopastoral systems due to trampling and browsing pressure caused by animals. For this reason, it is necessary to protect selected saplings (e.g. with small exclosures) to allow the establishment of tree samplings, a practice that is common in e.g. wooded meadows. It may be necessary also to change weed control practices with fire and herbicides. Such practices have in some instances positive effects on tree regeneration (Spooner et al. Citation2002; Esquivel et al. Citation2008; Fischer et al. Citation2009), but have negative effects on soil conservation and climate change mitigation (Pagiola et al. Citation2007; Verchot et al. Citation2007; Casals et al. Citation2014, Hoosbeek et al. Citation2018; Álvarez et al. Citation2021b). Tree species conservation and sustainable use may be also enhanced through incentives such as direct payments (e.g. Payments for Ecosystem Services) in silvopastoral systems in Colombia (Calle et al. Citation2013; Lee et al. Citation2020) promoting tree species preservation in livestock systems. These instruments can facilitate the transition towards new technologies and reduce the constraints that arise from the lack of labor (Barton et al. Citation2017; Nyberg et al. Citation2020) and access to knowledge (Calle et al. Citation2013).
Relation between the Index Cultural Importance (ICI) and livestock system types
The ES encompass contributions of nature to people (Díaz et al. Citation2015), and are a way of conceptualizing and assessing the importance attributed by humans to nature. ES thus imply a conceptualization of socio-ecological system (Pascual et al. Citation2010; Haines-Young Citation2011), characterized by a set of interlinked elements which enables the analysis of the capacity of an ecosystem to generate these benefits and the level of their appreciation by people (Barton et al. Citation2018). In this context, the cultural value index is a quantitative representation of the importance of the benefits attributed to different tree species in these silvopastoral systems.
The relationship between silvopastoral paddock typologies and cultural value indexes is closely linked to the functional diversity of tree species and the diversity of ES that support the production of livestock systems. Díaz et al. (Citation2011) state that the strategy of the farmers tends to maximize the benefits through the intensive and specialized use of a small number of ES that support the products for markets, although as shown in our study, and in agreement with other studies on silvopastoral systems in Latin America (Rusch et al. Citation2017), regulating and cultural services are important too.
In this study, only 16 tree species presented indices of cultural value, linked to a high dependence on some ES provided by trees. This coincides with the work carried out by Cáceres et al. (Citation2015), in the Gran Chaco region of Argentina, where large farmers and ranchers recognized a dependence of only a small number of ES generated by tree species, but contrasts with a high diversity of uses and knowledge of silvopastoral trees in African systems (Barton et al. Citation2016). ES assessments need to consider that different social actors perceive, access and use ecosystems and their functional diversity in very different ways (Díaz et al. Citation2011).
Silvopastoral paddock features, farmers’ decisions and ecosystem services
The tree composition of paddock types in the Colombian Amazon region appears to be shaped by the biophysical characteristics of the sites, the history of land-use, the regeneration capacity of the species under grazing, and finally by the management decisions of farmers. These particularities are given by the combination of different management strategies, land use history, cropping systems and socio-economic conditions (Harvey et al. Citation2011). Based on their perception about the benefits and costs related to keeping and promoting trees in the farm, farmers can actively design the composition and structure of the paddock types using their knowledge on tree species, trees’ costs and benefits, their expressed needs, and aspirations, as well as scientific knowledge regarding the ES and benefits they generate (Barton et al. Citation2016). Further, the use of an ES approach in our study, enabled to reveal a plurality of values associated with the silvopastoral ecosystem in the Colombian Amazon region, including regulating functions and their contribution to maintaining neotropical biodiversity (IPBES Citation2022). As called for by Pérez Marulanda et al. (Citation2020), this step can enable the inclusion of additional environmental benefits generated by these silvopastoral landscapes which are unaccounted in current assessments of socio-economic and environmental sustainability, and can help identify possible conflicts among multiple objectives when managing production systems, as well as address and solve trade-offs (Rusch et al. Citation2017).
The HSCR type corresponded to paddocks with complex structure and the greatest richness of tree species. The type is dominated by old trees, of great height and diameter, and low densities. These types of paddocks corresponded to only 10 paddocks out of the 159 paddocks comprised in the study. However, the conservation of trees in low densities in the paddock allows to maintain many ES in the livestock system which farmers value. Martínez-Enciso and Villanueva-López (Citation2013) found very similar results, reporting that large trees (large size in diameter and height) were less frequent in the farms evaluated in the Sierra de Tabasco, Mexico. This type likely has its origin in mature forests that were selectively clear-cut, where the farmer has maintained large and tall trees for the generation of ecosystem services (shade, feed, scenic beauty), and seeking the welfare of the cattle (Barton et al. Citation2016). Esquivel et al. (Citation2011) and Harvey et al. (Citation2011) reach to similar conclusions for systems in Central America, indicating that big and sparse trees are remnants after clearing primary forest. The trees have been preserved in the pasture areas to generate shade for the animals and are considered not to compete for pasture production (Anfinnsen et al. Citation2009). This kind of management is likely due to the farmer’s perception of the benefits of shade for the livestock, and at the same time to their knowledge about how to manage levels of shade that do not affect pasture productivity (Rusch et al. Citation2014; Esquivel et al. 2019; Álvarez et al. Citation2021a).
DMT paddocks encompass more than half of the paddocks in the study. They have characteristics such as the presence of young trees, medium heights and diameters, low abundance of individual trees species and high tree species richness. These characteristics are likely due to two factors: i) farmers tend to manage a wide diversity of species in grazing areas to meet different needs and ii) the colonization of trees in the paddocks is not controlled by regular practices of shrub clearing, burning, or the use of rotational grazing with temporary high grazing pressures. Farmers can selectively reduce the damage and mortality of preferred trees by protecting seedlings and saplings in their livestock systems (Camargo et al. Citation2000). Species with large canopy are kept at low densities, while species with smaller canopy areas are found at high densities (Esquivel et al. Citation2003). In our study, the composition, diversity, and structure of HDMT paddocks seem to be determined by the farmers weighting in other benefits than grass production in their decisions about the tree composition of the silvopastoral paddocks. Maintaining diverse paddocks can be a means to resolve tradeoffs among ES, such as the production of feed by trees during the dry season and grass production (Harvey et al. Citation2011). High tree density in livestock systems can be limited by the higher workload in managing these systems (Barton et al. Citation2016) and lower proportions of off-farm revenue in smallholder farms (Nyberg et al. Citation2020).
On the other hand, the lowest tree species richness of SSLR type can be due to a different understanding about the function and the generation of ES by silvopastoral systems. Esquivel et al. (Citation2003) state that tree cover and density are low, irregular, and dispersed in paddocks when most of the trees are eliminated during the establishment of monoculture pastures. It can indicate a form of degradation of the silvopastoral system, likely due to the farmers facing stronger fodder constraints and perceiving the benefits generated by the trees as less important.
In summary, our findings show highly different composition of paddock types, which are likely the result of a complex association of diverse factors, emerging from the ecological and socio-economic context in which these ecosystem services are produced and managed.
Conclusions
The complexity of the tree cover in the silvopastoral paddocks analyzed was related to the number of ES offered by trees including the provision of shade, the supply of wood and the protection of water sources, as the attributes mentioned by the farmers to justify tree selection in the pastures.
In this first study on the kinds of livestock systems in the Colombian Amazonia, we built a typology of silvopastoral paddocks condition. The typology is based on variables of species composition and tree measurements within these systems. Differences among types are mainly related to species richness, density and structure of shade tree species. These typologies are also related to the knowledge of the farmers, the ES they generate, and the value given by the farmers.
A high diversity of tree species has persisted in this silvopastoral landscape despite the expansion of livestock. This persistence is related to the favorable perception of the livestock farmers on the different ES generated by some tree species selected during the processes of natural regeneration, planted in the paddocks or by remnant trees from original forest.
These results are even more important with the current dynamics of the territory in the post-conflict context of the Colombian Amazon (Armenteras et al. Citation2006). We believe they are a basis to encourage the conservation of the diversity of trees in silvopastoral systems in the region. Future work may consider improving the knowledge by farmers on the potential of diverse systems to generate ES and about the important tree diversity the livestock systems may host. Setting aside non-use areas of forest with low human impact within the landscape is critical because the silvopastoral system helps maintain only a subset of the regional species pool. Making available plant material to reproduce native tree species in the livestock production system is also of great importance, so that trees are not replaced by a few exotics, globally distributed species. Focusing on farmers’ knowledge on tree ES can help in this direction of strengthening native tree species conservation and related ES within the livestock production landscape.
Supplementary Material
Download PDF (257.5 KB)Acknowledgment
We wish to thank the livestock producers at the municipalities of Belén de los Andaquíes, Morelia, Florencia, and El Doncello in the department of Caquetá - Colombia for their enthusiasm and cooperation with the project. We also thank the Colombian Ministry of Science, Technology and Innovation for financing the project “Effect of tree cover on pastures in dual-purpose livestock systems in the Colombian Amazon” Code 1131 - 521 - 28272 with contract number RC N° 514 - 2011 between the Universidad de la Amazonia and the Ministry of Science, Technology and Innovation. GMR was funded by the Norwegian Institute for Nature Research – Researchers’ funds through the Research Council of Norway.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary data
Supplemental data for this article can be accessed online at https://doi.org/10.1080/26395916.2023.2213784.
References
- Ainsworth JA, Moe SR, Skarpe C. 2012. Pasture shade and farm management effects on cow productivity in the tropics. Agric Ecosyst Environ. 155:105–19. doi:10.1016/j.agee.2012.04.005.
- Aitken SN, Yeaman S, Holliday JA, Wang T, Curtis‐mclane S. 2008. Adaptation, migration or extirpation: climate change outcomes for tree populations. Evol Appl. 1(1):95–111. doi:10.1111/j.1752-4571.2007.00013.x.
- Álvarez F, Casanoves F, Suárez JC 2021b. Influence of scattered trees in grazing areas on soil properties in the Piedmont region of the Colombian Amazon. PLos One. 16(12):e0261612. doi:10.1371/journal.pone.0261612.
- Álvarez F, Casanoves F, Suárez JC, Pezo D 2021a. The effect of different levels of tree cover on milk production in dual-purpose livestock systems in the humid tropics of the Colombian Amazon region. Agroforest Syst. 95:93–102. doi:10.1007/s10457-020-00566-7.
- Anfinnsen B, Aguilar-Støen M, Vatn A 2009. Actitudes de los productores ganaderos de El Petén, Guatemala, respecto a la implementación de sistemas silvopastoriles. Agroforestería en las Américas. 47:20–26.
- Armenteras D, Murcia U, González TM, Barón OJ, Arias JE. 2019. Scenarios of land use and land cover change for NW Amazonia: impact on forest intactness. Global Ecol Conserv. 17:e00567. doi:10.1016/j.gecco.2019.e00567.
- Armenteras D, Rudas G, Rodriguez N, Sua S, Romero M. 2006. Patterns and causes of deforestation in the Colombian Amazon. Ecol Indic. 6(2):353–368. doi:10.1016/j.ecolind.2005.03.014.
- Barton DN, Benjamin T, Cerdán CR, DeClerck F, Madsen AL, Rusch GM, Salazar ÁG, Sánchez D, Villanueva C. 2016. Assessing ecosystem services from multifunctional trees in pastures using Bayesian belief networks. Iss Environ Sci Tech. 18:165–174. doi:10.1016/j.ecoser.2016.03.002.
- Barton DN, Cisse Y, Kaya B, N’Diaye I, Yossi H, Diarra A, Keita S, Dembele A, Maiga D, Rusch GM, et al. 2017. Diagnosing agrosilvopastoral practices using Bayesian networks. Agroforest Syst. 91(2):325–334. doi:10.1007/s10457-016-9931-1.
- Barton DN, Kelemen E, Dick J, Martín-López B, Gómez-Baggethun E, Jacobs S, Hendriks CM, Termansen M, García-Llorente M, Primmer E., et al. 2018. (Dis) integrated valuation–Assessing the information gaps in ecosystem service appraisals for governance support. Iss Environ Sci Tech. 29:529–541. doi:10.1016/j.ecoser.2017.10.021.
- Blackshaw JK, y Blackshaw AW. 1994. Heat stress in cattle and the effect of shade on production and behaviors: a review. Aust J Exp Agr. 34(2):285–295. doi:10.1071/EA9940285.
- Cáceres DM, Tapella E, Quétier F, Díaz S. 2015. The social value of biodiversity and ecosystem services from the perspectives of different social actors. Ecol Soc. 20(1). doi:10.5751/ES-07297-200162.
- Cajas-Giron YS, y Sinclair FL. 2001. Characterization of multistrata silvopastoral systems on seasonally dry pastures in the Caribbean region of Colombia. Agroforest Syst. 53(2):215–225. doi:10.1023/A:1013384706085.
- Calle Z, Murgueitio E, Chará J, Molina CH, Zuluaga AF, Calle A. 2013. A strategy for scaling-up intensive silvopastoral systems in Colombia. J Sustain Forestry. 32(7):677–693. doi:10.1080/10549811.2013.817338.
- Camargo JC, Ibrahim M, Somarriba E, Finegan B, y Current D. 2000. Factores ecológicos y socioeconómicos que influyen en la regeneración natural de laurel en sistemas silvopastoriles del trópico húmedo y subhúmedo de Costa Rica. Agroforestería en las Américas. 7(26):46.
- Casals P, Romero J, Rusch GM, Ibrahim M. 2014. Soil organic C and nutrient contents under trees with different functional characteristics in seasonally dry tropical silvopastures. Plant Soil. 374(1–2):643–659. doi:10.1007/s11104-013-1884-9.
- Casanoves F, Macchiavelli R, Balzarini M. 2005. Error variation in multienvironment peanut trials: within-trial spatial correlation and between-trial heterogeneity. Crop Sci. 45(5):1927–1933. doi:10.2135/cropsci2004.0547.
- Curtis JT, Mcintosh RP. 1950. The interrelations of certain analytic and synthetic phytosociological characters. Ecology. 31(3):434–455. doi:10.2307/1931497.
- Dávalos LM, Dávalos E, Holmes J, Tucker C, Armenteras D. 2021. Bosques, coca y conflicto: la dinámica de la frontera de pasturas y deforestación en la Amazonía andina. J Illicit Econ Dev. 3(1):74–98. doi:10.31389/jied.87.
- Díaz S, Demissew S, Carabias J, Joly C, Lonsdale M, Ash N, Larigauderie A, Adhikari JR, Arico S, Báldi A, et al. 2015. The IPBES conceptual framework - connecting nature and people. Curr Opin Env Sust. 14:1–16. doi:10.1016/j.cosust.2014.11.002.
- Díaz S, Quétier F, Cáceres DM, Trainor SF, Pérez-Harguindeguy N, Bret-Harte MS, Finegan B, Peña-Claros M, Poorter L. 2011. Linking functional diversity and social actor strategies in a framework for interdisciplinary analysis of nature’s benefits to society. Proceedings of the National Academy of Sciences, 108(3):895–902.
- Di Rienzo JA, Casanoves F, Balzarini MG, González L, Tablada M, y Robledo CW. 2018. InfoStat, versión 2018. Argentina: Grupo InfoStat, FCA, Universidad Nacional de Córdoba.
- Di Rienzo JA, Casanoves F, Pla L, Vilchez S, Di Rienzo MJ. 2010. Qeco-Quantitative ecology software: a collaborative approach. Nota Informativa, Revista Latinoamericana de Conservación. 1:73–75.
- Dixon HJ, Dolores JW, Joshi L, Sinclair FL. 2001. Agroecological knowledge toolkit for windows: methodological guidelines, computer software and manual for AKT5. In: School of agriculture and forest scinces. Bangor: University of Wales; p. 171.
- Esquivel MJ, Harvey CA, Finegan B, Casanoves F, Skarpe C. 2008. Effects of pasture management on the natural regeneration of neotropical trees. J Appl Ecol. 45(1):371–380. doi:10.1111/j.1365-2664.2007.01411.x.
- Esquivel MJ, Harvey CA, Finegan B, Casanoves F, Skarpe C, y Nieuwenhuyse A. 2009. Regeneración natural de árboles y arbustos en potreros activos de Nicaragua. Agroforestería en las Américas. 47:76–84.
- Esquivel H, Ibrahim M, Harvey CA, Benjamin T, Sinclair FL. 2011. Dispersed trees in pasturelands of cattle farms in a tropical dry ecosystem. Trop Subtropical Agroecosyst. 14(3):933–941.
- Esquivel H, Ibrahim M, Harvey CA, Villanueva C, Benjamin T, Sinclair FL. 2003. Árboles dispersos en potreros de fincas ganaderas en un ecosistema seco de Costa Rica. Agroforestería en las Américas. 10:24–29.
- Fischer J, y Lindenmayer DB. 2002. The conservation value of paddock trees for birds in a variegated landscape in southern New South Wales. 2. Paddock trees as steppingstones. Biodivers Conserv. 11(5):833–849. doi:10.1023/A:1015318328007.
- Fischer J, Stott J, Zerger A, Warren G, Sherren K, Forrester RI. 2009. Reversing a tree regeneration crisis in an endangered ecoregion. 106: 10386–10391. doi:10.1073/pnas.0900110106.
- Fonseca D et al. 2009. Cobertura de la tierra: una mirada desde la perspectiva del cambio climáticoMonitoreo de los bosques y otras coberturas de la Amazonia colombiana. Instituto Sinchi. Ministerio de Medio Ambiente, Vivienda y Desarrollo Territorial. Bogotá. 83–98.
- Glaser BG, Strauss AL, Strutzel E. 1968. The discovery of grounded theory; strategies for qualitative research. Nurs Res. 17(4):364. doi:10.1097/00006199-196807000-00014.
- Grande D, de León F, Nahed J, y Pérez-Gil F. 2010. Importance and function of scattered trees in pastures in the Sierra of Tabasco, Mexico. Res J Biol Sci. 5(1):75–87. doi:10.3923/rjbsci.2010.75.87.
- Grime JP. 1998. Benefits of plant diversity to ecosystems: immediate, filter and founder effects. J Ecol. 86(6):902–910. doi:10.1046/j.1365-2745.1998.00306.x.
- Griscom HP, Connelly AB, Ashton MS, Wishnie MH, Deago J. 2011. The structure and composition of a tropical dry forest landscape after land clearance; Azuero Peninsula, Panama. J Sustain Forestry. 30(8):756–774. doi:10.1080/10549811.2011.571589.
- Grondard N, Hein L, Van Bussel LGJ. 2021. Ecosystem accounting to support the common agricultural policy. Ecol Indic. 131:108157. doi:10.1016/j.ecolind.2021.108157.
- Guevara S, Laborde J, Sánchez G 1998. Are isolated remnant trees in pastures a fragmented canopy?. Selbyana. 19(1):34–43.
- Gutiérrez G, Suarez J, Álvarez CF, Orjuela JA. 2012. Árboles dispersos en potreros y conectividad estructural en el paisaje de fincas ganaderas en la amazonia colombiana. Ingenieria Amazonia. 5(1):30–41.
- Haines-Young R. 2011. Exploring ecosystem service issues across diverse knowledge domains using Bayesian belief networks. Progress Phys Geograph. 35(5):681–699. doi:10.1177/0309133311422977.
- Hartel T, Hanspach J, Moga CI, Holban L, Á S, Tamás R, Réti KO 2018. Abundance of large old trees in wood-pastures of Transylvania (Romania). Sci Total Environ. 613:263–270.
- Hartel T, Réti KO, Craioveanu C. 2017. Valuing scattered trees from wood-pastures by farmers in a traditional rural region of Eastern Europe. Agric Ecosyst Environ. 236:304–311. doi:10.1016/j.agee.2016.11.019.
- Harvey CA, y Haber WH. 1999. Remnant trees and the conservation of biodiversity in Costa Rican pastures. Agroforest Syst. 44:37–68. doi:10.1023/A:1006122211692.
- Harvey CA, Medina A, Sanchez D, Vilchez S, Hernandez B, Saenz J, Maes JM, Casanoves F, Sinclair FL. 2006. Patterns of animal diversity associated with different forms of tree cover retained in agricultural landscapes. Ecol Appl. 16:1986–1999. doi:10.1890/1051-0761(2006)016[1986:POADID]2.0.CO;2.
- Harvey CA, Villanueva C, Esquivel H, Gómez R, Ibrahim M, Lopez M, y Villacís J. 2011. Conservation value of dispersed tree cover threatened by pasture management. For Ecol Manage. 261(10):1664–1674. doi:10.1016/j.foreco.2010.11.004.
- Holdridge L. 1978. Ecología basada en zonas de vida. San José, Costa Rica: IICA; p. 216.
- Holl KD 1999. Factors limiting tropical rain forest regeneration in abandoned pasture: seed rain, seed germination, microclimate, and soil. Biotropica. 31(2):229–242.
- Hoosbeek MR, Remme RP, Rusch GM. 2018. Trees enhance soil carbon sequestration and nutrient cycling in a silvopastoral system in south-western Nicaragua. Agroforest Syst. 92(2):263–273.
- Hosseininasab MS, Barani H, Akbarlou M, Moayeri MH. 2017. The structure of plant population of forested rangeland in different legal definitions (case study: Sabzkouh Region, Chaharmahal & Bakhtiari Province, Iran). J Rangel Sci. 7(4):361–375.
- IDEAM. 2017. Resultados monitoreo de la deforestación 2017. Instituto de Hidrología, Meteorología y Estudios Ambientales IDEAM. Centro de Documentación e Información Científico Técnica. Bogotá – Colombia. 45.
- IGAC. 1993. Aspectos ambientales para el ordenamiento territorial del Occidente del Departamento de Caquetá. Bogotá, DC: Instituto Geográfico Agustín Codazzi —IGAC—. Programa Investigaciones para la Amazonia —INPA—. Serie Estudios en la Amazonia Colombiana VI. Tercer Mundo Editores.
- IGAC. 2014. Estudio general de suelos y zonificación de tierras: departamento de Caquetá, escala 1/100.000. Subdirección de agrología. Instituto Geográfico Agustín Codazzi. 410.
- IPBES. 2019. Summary for policymakers of the global assessment report on biodiversity and ecosystem services. Paris: Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES).
- IPBES. 2022. Summary for policymakers of the methodological assessment regarding the diverse conceptualization of multiple values of nature and its benefits, including biodiversity and ecosystem functions and services (assessment of the diverse values and valuation of nature). Bonn: Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES).
- Jose S, Dollinger J. 2019. Silvopasture: a sustainable livestock production system. Agroforest Syst. 93(1):1–9. doi:10.1007/s10457-019-00366-8.
- Lee S, Bonatti M, Löhr K, Palacios V, Lana MA, Sieber S, Li F. 2020. Adoption potentials and barriers of silvopastoral system in Colombia: case of Cundinamarca region. Cogent Environ Sci. 6(1):1823632. doi:10.1080/23311843.2020.1823632.
- Lombo DF, Tobar D, Villanueva C. 2011. Estado de la cobertura arbórea en pasturas activas en dos paisajes agropecuarios en el neotrópico americano. En: Villanueva et al. editor. Manejo agroecológico como ruta para lograr la sostenibilidad de fincas con café y ganadería. Turrialba, CR: CATIE; p. 93.
- López M, Gómez R, Harvey CA, Villanueva C. 2004. Caracterización del componente arbóreo en los sistemas ganaderos de Rivas, Nicaragua. Encuentro. 36(68):114–133. doi:10.5377/encuentro.v0i68.4261.
- Manning AD, Fischer J, Lindenmayer D. 2006. Scattered trees are keystone structures – Implications for conservation. Biol Conserv. 132(3):311–321. doi:10.1016/j.biocon.2006.04.023.
- Martínez-Enciso C, Villanueva-López G, y Casanova-Lugo F. 2013. México. Agrociencias. 47(5):483–496.
- Medina A, Harvey CA, Sánchez D, Vilchez S, y Hernández B. 2004. Diversidad y composición de Chiropteros en un paisaje fragmentado de bosque seco en Rivas, Nicaragua. Encuentros. 68(68):24–43. doi:10.5377/encuentro.v0i68.4255.
- Medina A, Harvey CA, Sánchez D, Vílchez S, y Hernández B. 2007. Bat diversity and movement in a neotropical agricultural landscape. Biotropica. 39:120–128. doi:10.1111/j.1744-7429.2006.00240.x.
- Messeder JVS, Silveira FA, Cornelissen TG, Fuzessy LF, Guerra TJ. 2020. Frugivory and seed dispersal in a hyperdiverse plant clade and its role as a keystone resource for the Neotropical fauna. Ann Bot. doi:10.1093/aob/mcaa189.
- Murcia U, Medina R, Rodríguez JM, Castellanos H, Hernández A, y Herrera E. 2014. Monitoreo de los bosques y otras coberturas de la Amazonia Colombiana, a escala 1:100.000. Datos del periodo 2012. Instituto Amazónico de Investigaciones Científicas Sinchi. Bogotá, D. C., 190.
- Nepstad DC, Uhl C, Pereira CA, Da Silva JMC. 1996. A comparative study of tree establishment in abandoned pasture and mature forest of eastern Amazonia. Oikos. 25–39.
- Nyberg Y, Wetterlind J, Jonsson M, Öborn I. 2020. The role of trees and livestock in ecosystem service provision and farm priorities on smallholder farms in the Rift Valley, Kenya. Agric Syst. 181:102815. doi:10.1016/j.agsy.2020.102815.
- Pagiola S, Ramírez E, Gobbi J, de Haan C, Ibrahim M, Murguetitio E, Ruiz JP. 2007. Paying for the environmental services of silvopastoral practices in Nicaragua. Ecol Econ. 64:374–385. doi:10.1016/j.ecolecon.2007.04.014.
- Pascual U, Muradian R, Rodríguez LC, Duraiappah A. 2010. Exploring the links between equity and efficiency in payments for environmental services: a conceptual approach. Ecol Econ. 69(6):1237–1244. doi:10.1016/j.ecolecon.2009.11.004.
- Pearce F. 2019. Rivers in the sky. New Scientist. 244(3254):40–43.
- Pérez Marulanda L, Lavelle P, Rudbeck Jepsen M, Castro-Nunez A, Francesconi W, Camilo K, Vanegas M, Romero M, Suárez JC, Solarte A, et al. 2020. Farmscape composition and livelihood sustainability in deforested landscapes of Colombian Amazonia. Agriculture. 10(12):588. doi:10.3390/agriculture10120588.
- Pignataro AG, Levy-Tacher SI, Aguirre-Rivera JR, Nahed-Toral J, González-Espinosa M, González-Arzac A, Biganzoli F. 2017. Natural regeneration of tree species in pastures on peasant land in Chiapas, Mexico. Agric Ecosyst Environ. 249:137–143. doi:10.1016/j.agee.2017.08.020.
- Pignataro AG, Tacher SIL, Rivera JRA, Toral JN, Espinosa MG, Carmona NR. 2016. Silvopastoral systems of the Chol Mayan ethnic group in southern Mexico: strategies with a traditional basis. J Environ Manage. 181:363–373. doi:10.1016/j.jenvman.2016.06.036.
- Ramírez BL, Lavelle P, Orjuela JA, y Villanueva O. 2012. Characterization of cattle farms and adoption of agroforestry systems as a proposal for soil management in Caquetá, Colombia. Revista Colombiana de Ciencias Pecuarias. 25(3):391–401.
- R core team. 2018. R: a language and environment for statistical computing. Vienna: R foundation for statistical computing. [accessed 2018 Nov 20].
- Rodríguez-León CH, Roa-Fuentes LL, Sterling A, Suárez JC. 2022. Plant biodiversity homogenization across the chronosequence in highly fragmented landscapes in the Colombian Andean–Amazonian transition. Forests. 13(9):1422. doi:10.3390/f13091422.
- Ruiz F, Gomez R, Harvey CA. 2005. Caracterización del componente arbóreo en los sistemas ganaderos de Matiguás, Nicaragua. Managua, Nicaragua: Revista Tropitecnica de Nitlapan, UCA.
- Rusch VE, López D, Cavallero L, Rusch GM, Garibaldi LA, Grosfeld JE, Peri P. 2017. State-and-Transition model of Ñire forest in NW Patagonia as a tool for sustainable silvopastoral management. Ecología Austral. 27:266–278. (In Spanish). doi:10.25260/EA.17.27.2.0.240.
- Rusch VE, Rusch GM, Goijman AP, Varela SA, Claps L. 2017. Ecosystem services to support environmental and socially sustainable decision-making. Ecología Austral. 27:162–176. doi:10.25260/EA.17.27.1.1.295.
- Rusch GM, Zapata P, Casanoves F, Casals P, Ibrahim M, DeClerck F. 2014. Determinants of grassland primary production in seasonally-dry silvopastoral systems in Central America. Agroforest Syst. 88(3):517–526. doi:10.1007/s10457-014-9711-8.
- Sánchez MD, Harvey CA, Grijalva A, Medina A, Vílchez S, Hernández B. 2005. Diversidad, composición y estructura de la vegetación en un agropaisaje ganadero en Matiguás, Nicaragua. Revista de biología tropical. 53(3–4):387–414. doi:10.15517/rbt.v53i3-4.14601.
- Sánchez YKKÁ, Tapia MEP, Salazar JCS. 2017. Importancia cultural de vegetación arbórea en sistemas ganaderos del Municipio de San Vicente del Caguán, Colombia. Revista UDCA Actualidad Divulgación Científica. 20(2):393–401. doi:10.31910/rudca.v20.n2.2017.397.
- Soberón JM, y Llorente JB. 1993. The use of species accumulation functions for the prediction of species richness. Conserv Biol. 7(3):480–488. doi:10.1046/j.1523-1739.1993.07030480.x.
- Sotelo M, Suárez Salazar JC, Álvarez Carrillo F, Castro Núñez A, Calderón Soto VH, Arango J. 2017. Sistemas sostenibles de producción ganadera en el contexto amazónico - Sistemas silvopastoriles: ¿una opción viable? Publicación CIAT No. 448. Cali, Colombia: Centro Internacional de Agricultura Tropical (CIAT). p. 24.
- Spooner P, Lunt I, Robinson W. 2002. Is fencing enough? The short-term effects of stock exclusion in remnant grassy woodlands in southern NSW. Ecol Manage Restor. 3:117–126.
- Staal A, Flores BM, Aguiar APD, Bosmans JH, Fetzer I, Tuinenburg OA. 2020. Feedback between drought and deforestation in the Amazon. Environ Res Lett. 15(4):044024. doi:10.1088/1748-9326/ab738e.
- Stevens WD, Ulloa C, Pool A, Montiel OM 2001. Flora de Nicaragua (Vol. 85, No. 1, p. 943). St. Louis: Missouri Botanical Garden Press.
- Suárez J, Álvarez CF, Orjuela JA, y Aldana OD. 2013. Conocimiento local sobre el manejo de la cobertura arbórea en sistemas ganaderos bajo el sistema de doble propósito. En: Álvarez CF, editor. Árboles dispersos en potreros en fincas ganaderas del piedemonte amazónico. Florencia – Caquetá – Colombia: Universidad de la Amazonia; p. 54–78.
- Suárez J, Álvarez CF, Orjuela JA, Ocaña H, Londoño SM, Ceballos M, Rojas P, López N, García F, y Torres O. 2013. Caracterización de fincas ganaderas bajo el sistema de doble propósito en el departamento del Caquetá. En: Álvarez CF, editor. Árboles dispersos en potreros en fincas ganaderas del piedemonte amazónico. Florencia – Caquetá – Colombia: Universidad de la Amazonia; p. 39–52.
- Torrijos, RR. 2022. Cifras de contexto ganadero regional 2022. Ed. Comité Departamental .de Ganaderos del Caquetá. Florencia, Caquetá, Colombiap. 32.
- United Nations. 2021. Guidelines on biophysical modelling for ecosystem accounting - 2.0.
- Valdano SG, Di Rienzo JA. 2007. Discovering meaningful groups in hierarchical cluster analysis. An extension to the multivariate case of a multiple comparison method based on cluster analysis. Available via InterStat Journal. [accessed 2016 Oct 13]. http://interstat.statjournals.netsubordinate-document/http://interstat.statjournals.net/YEAR/2007/articles/0704002.pdf.
- Valtorta SE, Leva PE, Gallardo MR. 1997. Evaluation of different shades to improve dairy cattle well-being in Argentina. Int J Biometeorol. 41(2):65–67. doi:10.1007/s004840050055.
- Verchot LV, Van Noordwijk M, Kandi S, Tomich T, Ong C, Albrecht A, Mackensen J, Anupama KV, Palm C. 2007. Climate change: linking adaptation and mitigation through agroforestry. Mitig Adapt Strateg Glob Change. 12:901–918. doi:10.1007/s11027-007-9105-6.
- Vílchez S, Harvey CA, Sánchez D, Medina A, y Hernández B. 2004. Diversidad de aves en un paisaje fragmentado de bosque seco en Rivas, Nicaragua. Managua, Nicaragua. Encuentros. 68:60–75. doi:10.5377/encuentro.v0i68.4257.
- Villacis J, Harvey CA, Ibrahim M, Villanueva C. 2003. Relaciones entre la cobertura arbórea y el nivel de intensificación de las fincas ganaderas en Rio Frio, Costa Rica. Agroforestería en las Américas. 10:17–23.
- Villanueva C, Ibrahim M, Harvey C, Esquivel H. 2003. Tipologías de fincas con ganadería bovina y cobertura arbórea en pasturas en el trópico seco de Costa Rica. Agroforestería en las Américas. 10:9–16.
- Villanueva-Partida C, Casanova-Lugo F, Villanueva-López G, González-Valdivia N, Oros-Ortega I, Díaz-Echeverría V. 2016. Influence of the density of scattered trees in pastures on the structure and species composition of tree and grass cover in southern Tabasco, Mexico. Agric Ecosyst Environ. 232:1–8. doi:10.1016/j.agee.2016.07.020.
- Wassie A, Sterck FJ, Teketay D, y Bongers F. 2009. Effects of livestock exclusion on tree regeneration in church forests of Ethiopia. For Ecol Manage. 257(3):765–772. doi:10.1016/j.foreco.2008.07.032.
- Wunderle JM Jr. 1997. The role of animal seed dispersal in accelerating native forest regeneration on degraded tropical lands. For Ecol Manage. 99(1–2):223–235. doi:10.1016/S0378-1127(97)00208-9.