Abstract
The phloem mobility of heavy metals is relevant to the redistribution of micronutrients and pollutants and, ultimately, to the quality of harvested plant parts. The relative mobility in wheat may vary considerably between different cations. In the study reported here, radio-labeled nickel (Ni), cobalt (Co), manganese (Mn), zinc (Zn) and cadmium (Cd) were introduced into either intact young winter wheat (Triticum aestivum L. cv. Arina) via a leaf flap, or detached maturing shoots via the cut stem. Elements fed into the lamina of the second leaf of 21-day-old plants were translocated to the younger (expanding) leaves and to the roots but not or only in trace amounts to the first (already fully expanded) leaf. The 63Ni and 65Zn were exported more rapidly compared with the other heavy metals. Most of 54Mn was retained in the originally labeled leaf. The peduncle of some maturing shoots was steam-girdled below the ear to distinguish between xylem and phloem transport. This phloem interruption reduced the content of 63Ni in the ear to about 25%. Intermediate effects were observed for 65Zn, 57Co, and 109Cd. Total 54Mn accumulation in the ear was hardly affected by steam-girdling, indicating a transport of this element within the xylem to the ear. These results suggest that the relative phloem mobility of Ni and Zn in young wheat plants and in maturing wheat shoots is higher than the mobility of Co and Cd, whereas the mobility of Mn is very low.
INTRODUCTION
Some heavy metals, such as zinc (Zn), manganese (Mn), copper (Cu) and nickel (Ni) are required in trace amounts by higher plants to complete their life cycle (CitationMarschner, 1995; CitationYang et al., 1996a, Citation1996b, Citation1997), while others [i.e., cadmium (Cd), lead (Pb), and mercury (Hg)] are non-essential. In extended concentrations, however, all heavy metals are toxic. Increasing applications of sludge to agricultural soils and continuing release of industrial wastes cause a redistribution of heavy metals in the environment (CitationCataldo et al., 1978). Heavy-metal pollution is becoming more and more serious with the expansion of industrial production and modern city development (CitationZhang et al., 1999). Therefore, research concerning the uptake, transport, and redistribution of heavy metals in cultivated plants is highly relevant in preventing or reducing these elements from entering the food chain.
The primary distribution via the xylem, the retranslocation via the phloem, and the transfer from the xylem to the phloem must be considered as important processes for the distribution of an element within a plant. The transport in the xylem is directed from the roots to the shoot, whereas the phloem transport takes place from sources to sinks and is more selective (CitationMarschner, 1995). Xylem-to-phloem transfer can take place all along the pathway from the roots to the shoot, and the stem plays an important role in this context (CitationMcNeil, 1980; CitationVan Bel, 1984; CitationHerren and Feller, 1997), most likely via transfer cells (CitationJeschke and Pate, 1991).
In general, cobalt (Co) and nickel are considered heavy metals with an intermediate mobility in the phloem (CitationMarschner, 1995; CitationWelch, 1995). It has been shown that Ni has a slightly better phloem mobility in wheat than Co (CitationZeller, 1998). In maturing detached wheat shoots, Ni was removed more efficiently from the xylem in the peduncle than was Co and as a consequence less Co than Ni accumulated in the grains (CitationZeller and Feller, 1998). Moreover, Ni accumulated more rapidly than did Co, and in higher quantities in the grains of intact plants that were treated with both elements (CitationZeller and Feller, 1999). The mobility of Mn is generally very low (CitationMarschner, 1995). In comparison with Mn, Zn may move more readily in the phloem, probably complexed with organic acids (CitationHocking, 1980; CitationWhite et al., 1981), but not as rapidly as potassium (K) and phosphorus (P). Source-sink relationships can change considerably during the switch from vegetative to reproductive growth (CitationMarschner, 1995). In young wheat it has been demonstrated that Zn is transported by phloem from leaves to roots after foliar application (CitationHaslett et al., 2001). In maturing wheat, Zn moved from leaves to developing grains, but not from leaves to roots (CitationWebb and Loneragan, 1990). Cadmium (Cd) is chemically similar to Zn (CitationChesworth, 1991) and may be taken up and transported in plants via similar pathways (CitationGrant et al., 1998).
The aim of this study was to investigate the relative phloem mobility of Co, Mn, Ni, Zn, and Cd in wheat. Retranslocation of appropriate heavy metals was observed during vegetative growth and during later stages of maturation.
MATERIALS AND METHODS
Redistribution of Heavy Metals in Young Winter Wheat
Winter wheat grains (Triticum aestivum L. cv. Arina) were germinated on wet paper in darkness for two days and then transferred for five days to quartz sand. The seedlings then grew for 1 week in standard nutrient solution (CitationHildbrand et al., 1994) (1:4 diluted) before starting the treatment. A flap was cut into the second leaf lamina (CitationSchenk and Feller, 1990) and 200 μL nutrient solution containing a combination of 57Co, 63Ni, and 54Mn or 65Zn and 109Cd were introduced from a tube directly into the cut xylem of the flap. Since only trace amounts of associated anions (chloride) were added together with the radioactive heavy metal cations, no special controls with different anion species were prepared. Plants absorbed the feeding solution within 24 h. After 1, 2, 7, 14, 21, and 28 d from treatment, plants were dissected into roots, grain and coleoptile, the various leaves and the flap. The plant parts were dried at room temperature (20°C).
The radioactivity of 57Co,54Mn, 65Zn, and 109Cd in the different plant parts was detected by a gamma counter (1480 Wizard 3, Wallac Oy, Turku, Finland). For 63Ni measurements, plant parts were ashed at 550°C for 8 h. The ash was solubilized with 2 mL citric acid (20 mM). Then 200 μL were transferred in Ready CapsTM (Beckman Instruments, USA) and dried at < 65°C for 4 h. The radioactivity was then detected by beta counter (Betamatic V, Kontron Instruments).
Heavy Metal Redistribution in Detached Wheat Shoots
Two weeks after anthesis, randomly selected wheat shoots were cut in a field near Bern (Zollikofen) directly above soil surface. The shoots were submerged in deionized water and immediately recut below the third node from the top. Standing in water, these shoots were transported to the laboratory and cut submerged to their final length below the flag leaf node. The phloem of some shoots was interrupted below the ear by steam-girdling (CitationMartin, 1982). Steam-girdling kills all living cells in a 1–2 cm long part of the peduncle while the xylem remains functional. The shoots (four replicates per treatment) were incubated standing in nutrient solution (25 mL) containing either 57Co, 63Ni, 54Mn, 65Zn, or 109Cd. The experiment was terminated after four days. The basal parts of the shoots were rinsed twice in deionized water and dried at 105°C for 10 h. The shoots were dissected into grains, glumes, rachis, peduncle (including flag leaf node), lamina, and sheath. Then the radioactivity of the different radionuclides in the plant parts was measured as described above.
The plants for these experiments were incubated in a culture room with a light/dark cycle of 14 h light (4 Philips TLD 36W/25 and 2 Osram Fluora L 36W/77 fluorescent tubes) and 10 h darkness. The average temperature was 26°C during the day and 22°C during the night.
RESULTS
Redistribution of Heavy Metals in Young Winter Wheat
Cutting a flap into the second leaf and introducing 200 μL feeding solution caused no major effect on the dry matter accumulation in the various plant parts (data not shown).
The distribution of simultaneously introduced 57Co, 63Ni and 54Mn was different (). Most of 54Mn was retained throughout the 28 days in the second leaf. Only trace amounts of 54Mn were transported to other plant parts. In contrast to 54Mn, a considerable percentage of applied 63Ni was exported from the second leaf during the 28 days. The 63Ni was easily transported basipetally to the roots and to the youngest leaves but not into the first leaf. 63Ni was imported as long as the leaves expanded and was then exported to the new developing leaves. The remobilization of 57Co is between that of 54Mn and 63Ni. The 57Co was redistributed to the roots and to the younger leaves but less rapidly and in smaller quantities than was 63Ni. The transport of 57Co to the first leaf was negligible, as for 63Ni.
Figure 1. Co, Ni and Mn redistribution in young winter wheat. Intact 21 d old plants were supplied with 200 μL nutrient solution containing 57Co, 63Ni and 54Mn via a flap cut into the lamina of the second leaf. Plants were harvested 1, 2, 7, 14, 21 and 28 d after the treatment. Means and standard errors of 4 plants are shown.
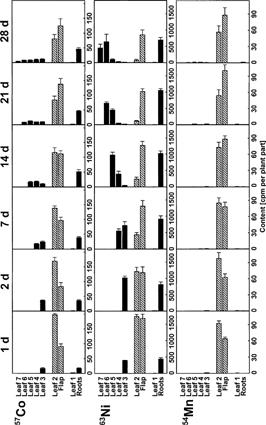
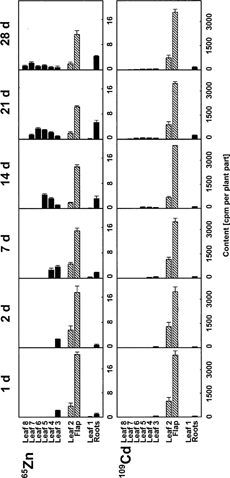
The results of an experiment with simultaneous application of 65Zn and 109Cd are shown in . Most of the radiolabel was detected in the leaf flap. Exported 65Zn was detected mainly in the roots and in the expanding leaves as observed for 63Ni, whereas 109Cd was exported only in minor quantities. Transport to the first leaf was barely detectable for 65Zn and 109Cd.
Figure 2. Zn and Cd redistribution in young winter wheat. Intact 21 d old plants were supplied with 200 μL nutrient solution containing 65Zn and 109Cd via a flap cut into the lamina of the second leaf. Plants were harvested 1, 2, 7, 14, 21, and 28 d after the treatment. Means and standard errors of 4 plants are shown.
Heavy Metal Redistribution in Detached Wheat Shoots
Steam-girdling below the ear of detached shoots had no major influence on total 54Mn accumulation in the ear and in the vegetative parts (). Within the ear more 54Mn reached the glumes, whereas less 54Mn was transported to the grains when plants were girdled (). The strongest effect of steam-girdling was observed for 63Ni. A phloem interruption reduced the content of 63Ni in the ear to about 25% and led to increased contents in the vegetative parts (). In shoots with an intact phloem, 63Ni was located mainly in the grains and the peduncle, whereas accumulation in the lamina was negligible (). When shoots were steam-girdled below the ear, 63Ni accumulated predominantly in the peduncle and in the sheath. In control plants 57Co and 65Zn were located mainly in the grains and in the peduncle (as observed for 63Ni). Similar to 54Mn, 109Cd was transported in those shoots in a higher percentage to the glumes and to the laminas.
Figure 3. Distribution of various heavy metals in field-grown wheat shoots detached below the flag leaf node. The shoots standing in a solution containing 2 mM SrCl2, 2 mM RbCl and 57Co, 63Ni, 54Mn, 65Zn or 109Cd were incubated for 4 d in a culture room. The phloem of some shoots was interrupted directly below the ear by steam-girdling immediately before the incubation. Means and standard errors of 4 plants are shown.
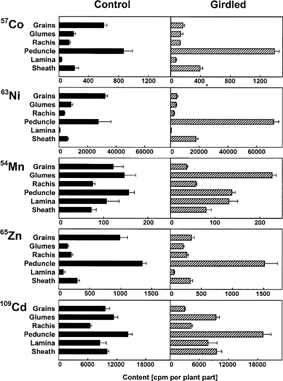
Table 1 Influence of steam-girdling below the ear on the accumulation of labelled heavy metals in the ear and in the vegetative parts [cpm plant part−1] of wheat shoots
DISCUSSION
The export of introduced heavy metals from the second leaf of young winter wheat was due mainly to phloem transport. Expanding leaves and roots represented major phloem sinks, whereas mature leaves acted as sources. Heavy metals did not accumulate in the primary leaf. Thus, there was neither a transport via the phloem from the labeled leaf, nor via the xylem from the pool of heavy metals that had been accumulated in the roots.
Maturing grains represented phloem sinks in detached shoots. The heavy metals reached the grains either directly, via the xylem, or were first transferred from the xylem to the phloem and then reached the grains within the phloem. This transfer from xylem to phloem might have happened in the leaf, in the peduncle, or in the glumes. After steam-girdling below the ear, transport of introduced heavy metals to the grains was no longer possible via the phloem. Therefore, heavy metals were transported in such plants only via the xylem, either directly to the grains or initially to the glumes and then retranslocated to the grains via the phloem. The 54Mn transport to the ear was barely affected when the phloem was interrupted, indicating that 54Mn reached the ear via the xylem. Nevertheless, steam-girdling had an effect on 54Mn redistribution. Less 54Mn was detected in the grains of girdled plants. The 54Mn accumulated in those shoots in the glumes, suggesting a reduced retranslocation of 54Mn from the glumes to the grains via the phloem.
The results demonstrated the differing relative phloem mobility of the introduced heavy metals in wheat. Compared with the other heavy metals, 63Ni showed the highest mobility in the phloem and 54Mn the lowest. Although 65Zn and 109Cd are chemically similar, their phloem mobility was different. The 65Zn was exported more rapidly and in larger amounts than was 109Cd. Moreover, 65Zn was more efficiently removed from the xylem sap compared with 109Cd, while a higher proportion of the latter was directed to organs with a high transpiration rate.
ACKNOWLEDGMENTS
We thank the Inforama Rütti in Zollikofen, Switzerland, for the supply of wheat plants. This work was a contribution to the National Centre of Competence in Research Plant Survival and was partially supported by the Swiss Federal Office of Environment, Forest and Landscape.
Related Research Data
REFERENCES
- Cataldo , D. A. , Garland , T. R. and Wildung , R. E. 1978 . Nickel in plants: Uptake kinetics using intact soybean plants . Plant Physiology , 62 : 563 – 565 .
- Chesworth , W. 1991 . “ Geochemistry of micronutrients ” . In Micronutrients in agriculture, , 2nd ed. , Edited by: Mortvedt , J. J. , Cox , R. R. , Shuman , L. M. and Welch , R. M. 1 – 30 . Madison, Wisconsin : Soil Science Society of America .
- Grant , C. A. , Buckley , W. T. , Bailey , L. D. and Selles , F. 1998 . Cadmium accumulation in crops . Canadian Journal of Plant Science , 78 : 1 – 17 . [CSA]
- Haslett , B. S. , Reid , R. J. and Rengel , Z. 2001 . Zinc mobility in wheat: Uptake and distribution of zinc applied to leaves or roots . Annals of Botany , 87 : 379 – 386 . [CROSSREF]
- Herren , T. and Feller , U. 1997 . Transport of cadmium via xylem and phloem in maturing wheat shoots: Comparison with the translocation of zinc, strontium and rubidium . Annals of Botany , 80 : 623 – 628 . [CROSSREF]
- Hildbrand , M. , Fischer , A. and Feller , U. 1994 . Protein catabolism in bean leaf discs: Accumulation of a soluble fragment of ribulose-1,5-bisphosphate carboxylase/oxygenase under oxygen deficiency . Journal of Experimental Botany , 45 : 1197 – 1204 .
- Hocking , P. J. 1980 . The composition of phloem exudate and xylem sap from tree tabacco (Nicotiana glauca Grah.) . Annals of Botany , 45 : 633 – 43 .
- Jeschke , W. D. and Pate , J. S. 1991 . Modelling of the partitioning, assimilation and storage of nitrate within root and shoot organs of castor bean Ricinus communis L.) . Journal of Experimental Botany , 42 : 1091 – 1103 .
- Marschner , H. 1995 . Mineral nutrition of higher plants , New York : Academic Press .
- Martin , P. 1982 . Stem xylem is a possible pathway for mineral translocation from senescing leaves to the ear in wheat . Australian Journal of Plant Physiology , 9 : 197 – 207 .
- McNeil , D. L. 1980 . The role of the stem in phloem loading of minerals in Lupinus albus L. cv. Ultra . Annals of Botany , 45 : 329 – 338 .
- Schenk , D. and Feller , U. 1990 . Rubidium export from individual leaves of maturing wheat . Journal of Plant Physiology , 137 : 175 – 179 . [CSA]
- Van Bel , A. J. E. 1984 . Quantification of the xylem-to-phloem transfer of amino acids by use of inulin (14C) carboxylic acid as xylem transport markers . Plant Science Letters , 35 : 81 – 85 . [CROSSREF]
- Webb , M. J. and Loneragan , J. F. 1990 . Zinc translocation to wheat roots and its implications for phosphorus/zinc interaction in wheat plants . Journal of Plant Nutrition , 13 : 1499 – 1512 .
- Welch , R. M. 1995 . Micronutrient nutrition of plants . Critical Reviews in Plant Sciences , 14 : 49 – 82 .
- White , M. C. , Becker , F. D. , Chaney , R. L. and Decker , A. M. 1981 . Metal complexation in xylem fluid . Plant Physiology , 67 : 292 – 300 .
- Yang , X. , Baligar , V. C. , Martens , D. C. and Clark , R. B. 1996a . Plant tolerance to nickel toxicity: I. Influx, transport, and accumulation of nickel in four species . Journal of Plant Nutrition , 19 : 73 – 85 .
- Yang , X. , Baligar , V. C. , Martens , D. C. and Clark , R. B. 1996b . Plant tolerance to nickel toxicity: II. Nickel effects on influx and transport of mineral nutrients in four species . Journal of Plant Nutrition , 19 : 265 – 279 .
- Yang , X. , Baligar , V. C. , Foster , J. C. and Martens , D. C. 1997 . Accumulation and transport of nickel in relation to organic acids in ryegrass and maize grown with different nickel levels . Plant and Soil , 196 : 271 – 276 . [CROSSREF]
- Zeller , S. and Feller , U. 1998 . Redistribution of cobalt and nickel in detached wheat shoots: Effects of steam-girdling and of cobalt and nickel supply . Biologia Plantarum , 41 : 427 – 434 . [CROSSREF]
- Zeller , S. and Feller , U. 1999 . Long-distance transport of cobalt and nickel in maturing wheat . European Journal of Agronomy , 10 : 91 – 98 . [CROSSREF]
- Zhang , H. , Ma , D. , Xie , Q. and Chen , X. 1999 . An approach to studying heavy metal pollution caused by modern city development in Nanjing, China . Environmental Geology , 38 : 223 – 228 . [CROSSREF]