
Abstract
Optimized body composition provides a competitive advantage in a variety of sports. Weight reduction is common among athletes aiming to improve their strength-to-mass ratio, locomotive efficiency, or aesthetic appearance. Energy restriction is accompanied by changes in circulating hormones, mitochondrial efficiency, and energy expenditure that serve to minimize the energy deficit, attenuate weight loss, and promote weight regain. The current article reviews the metabolic adaptations observed with weight reduction and provides recommendations for successful weight reduction and long term reduced-weight maintenance in athletes.
Electronic supplementary material
The online version of this article (doi:10.1186/1550-2783-11-7) contains supplementary material, which is available to authorized users.
Copyright comment
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
Introduction
In a variety of competitive sports, it is considered advantageous to achieve low levels of body fat while retaining lean body mass. The metabolic effects of this process have been given little context within athletics, such as physique sports (i.e. bodybuilding, figure), combat sports (i.e. judo, wrestling), aesthetic sports (i.e. gymnastics, figure skating), and endurance sports. Previous literature has documented cases of male bodybuilders reducing body fat to less than 5% of total body mass [Citation1, Citation2], and studies documenting physiological profiles of male wrestlers [Citation3] and judo athletes [Citation4] present body fat ranges that extend below 5%. A study on elite female gymnasts and runners reported an average body fat percentage (BF%) of 13.72% for the entire sample, with subgroups of middle-distance runners and artistic gymnasts averaging 12.18% and 12.36%, respectively [Citation5]. Elite female runners have also reported percent body fat levels below 10% [Citation6]. Energy deficits and extremely low levels of body fat present the body with a significant physiological challenge. It has been well documented that weight loss and energy restriction result in a number of homeostatic metabolic adaptations aimed at decreasing energy expenditure, improving metabolic efficiency, and increasing cues for energy intake [Citation7–Citation9]. While the unfavorable endocrine effects of contest preparation have been documented in male bodybuilders [Citation1, Citation2, Citation10], anecdotal reports from physique athletes also describe a state in which metabolic rate has slowed to an extent that exceeds the predicted magnitude, making weight loss increasingly difficult despite low caloric intakes and high training volumes. Although such reports could potentially be related to inaccurate dietary reporting [Citation11, Citation12], these claims may be substantiated by a number of metabolic adaptations to weight loss, including adaptive thermogenesis [Citation13–Citation15], increased mitochondrial efficiency [Citation16–Citation19], and hormonal alterations that favor decreased energy expenditure, decreased satiety, and increased hunger [Citation1, Citation2, Citation10]. As a dieting phase progresses, such adaptations may threaten dietary adherence, make further weight loss increasingly difficult, and predispose the individual to rapid weight regain following the cessation of the diet. Although data documenting the attainment and recovery from extreme changes in body composition is limited, the present article aims to investigate the condition of metabolic adaptation described by competitors and identify potential mechanisms to explain such a phenomenon.
The endocrine response to an energy deficit
A number of hormones play prominent roles in the regulation of body composition, energy intake, and energy expenditure. The hormones of the thyroid gland, particularly triiodothyronine (T3), are known to play an important and direct role in regulating metabolic rate. Increases in circulating thyroid hormones are associated with an increase in the metabolic rate, whereas lowered thyroid levels result in decreased thermogenesis and overall metabolic rate [Citation20]. Leptin, synthesized primarily in adipocytes, functions as an indicator of both short and long-term energy availability; short-term energy restriction and lower body fat levels are associated with decreases in circulating leptin. Additionally, higher concentrations of leptin are associated with increased satiety and energy expenditure [Citation21]. Insulin, which plays a crucial role in inhibiting muscle protein breakdown [Citation22] and regulating macronutrient metabolism, is considered another “adiposity signal” [Citation23]. Similar to leptin, high levels of insulin convey a message of energy availability and are associated with an anorexigenic effect. Conversely, the orexigenic hormone ghrelin functions to stimulate appetite and food intake, and has been shown to increase with fasting, and decrease after feeding [Citation24]. Testosterone, known primarily for its role in increasing muscle protein synthesis and muscle mass [Citation22], may also play a role in regulating adiposity [Citation25]. Changes in fat mass have been inversely correlated with testosterone levels, and it has been suggested that testosterone may repress adipogenesis [Citation25]. More research is needed to delineate the exact mechanism (s) by which testosterone affects adiposity. Cortisol, a glucocorticoid that influences macronutrient metabolism, has been shown to induce muscle protein breakdown [Citation22], and increased plasma cortisol within the physiologic range has increased proteolysis in healthy subjects [Citation26]. Evidence also suggests that glucocorticoids may inhibit the action of leptin [Citation27].
Results from a number of studies indicate a general endocrine response to hypocaloric diets that promotes increased hunger, reduces metabolic rate, and threatens the maintenance of lean mass. Studies involving energy restriction, or very low adiposity, report decreases in leptin [Citation1, Citation10, Citation28], insulin [Citation1, Citation2], testosterone [Citation1, Citation2, Citation28], and thyroid hormones [Citation1, Citation29]. Subsequently, increases in ghrelin [Citation1, Citation10] and cortisol [Citation1, Citation30, Citation31] have been reported with energy restriction. Further, there is evidence to suggest that unfavorable changes in circulating hormone levels persist as subjects attempt to maintain a reduced body weight, even after the cessation of active weight loss [Citation32, Citation33].
Low energy intake and minimal body fat are perceived as indicators of energy unavailability, resulting in a homeostatic endocrine response aimed at conserving energy and promoting energy intake. It should be noted that despite alterations in plasma levels of anabolic and catabolic hormones, losses of lean body mass (LBM) often fail to reach statistical significance in studies on bodybuilding preparation [Citation1, Citation2]. Although the lack of significance may relate to insufficient statistical power, these findings may indicate that unfavorable, hormone-mediated changes in LBM can potentially be attenuated by sound training and nutritional practices. Previous research has indicated that structured resistance training [Citation34] and sufficient protein intake [Citation35–Citation37], both commonly employed in bodybuilding contest preparation, preserve LBM during energy restriction. Further, Maestu et al. speculate that losses in LBM are dependent on the magnitude of weight loss and degree of adiposity, as the subjects who lost the greatest amount of weight and achieved the lowest final body fat percentage in the study saw the greatest losses of LBM [Citation2]. The hormonal environment created by low adiposity and energy restriction appears to promote weight regain and threaten lean mass retention, but more research is needed to determine the chronic impact of these observed alterations in circulating anabolic and catabolic hormones.
Weight loss and metabolic rate
An individual’s total daily energy expenditure (TDEE) is comprised of a number of distinct components (Figure ). The largest component, resting energy expenditure (REE), refers to the basal metabolic rate (BMR) [Citation8]. The other component, known as non-resting energy expenditure (NREE), can be further divided into exercise activity thermogenesis (EAT), non-exercise activity thermogenesis (NEAT), and the thermic effect of food (TEF) [Citation8].
Figure 1 Components of total daily energy expenditure (TDEE). BMR = basal metabolic rate; NEAT = non-exercise activity thermogenesis; TEF = thermic effect of food; EAT = exercise activity thermogenesis; REE = resting energy expenditure; NREE = non-resting energy expenditure. Adapted from Maclean et al., 2011.
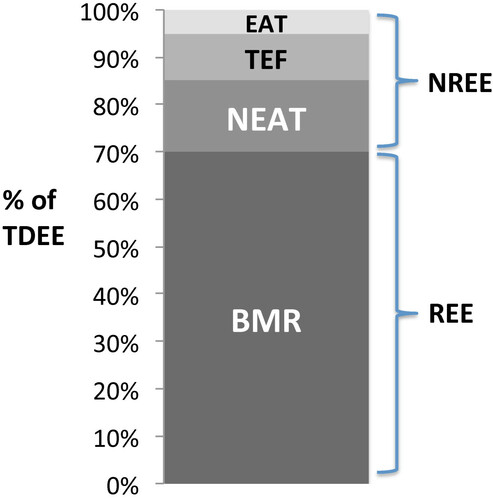
Metabolic rate is dynamic in nature, and previous literature has shown that energy restriction and weight loss affect numerous components of energy expenditure. In weight loss, TDEE has been consistently shown to decrease [Citation38, Citation39]. Weight loss results in a loss of metabolically active tissue, and therefore decreases BMR [Citation38, Citation39]. Interestingly, the decline in TDEE often exceeds the magnitude predicted by the loss of body mass. Previous literature refers to this excessive drop in TDEE as adaptive thermogenesis, and suggests that it functions to promote the restoration of baseline body weight [Citation13–Citation15]. Adaptive thermogenesis may help to partially explain the increasing difficulty experienced when weight loss plateaus despite low caloric intake, and the common propensity to regain weight after weight loss.
Exercise activity thermogenesis also drops in response to weight loss [Citation40–Citation42]. In activity that involves locomotion, it is clear that reduced body mass will reduce the energy needed to complete a given amount of activity. Interestingly, when external weight is added to match the subject’s baseline weight, energy expenditure to complete a given workload remains below baseline [Citation41]. It has been speculated that this increase in skeletal muscle efficiency may be related to the persistent hypothyroidism and hypoleptinemia that accompany weight loss, resulting in a lower respiratory quotient and greater reliance on lipid metabolism [Citation43].
The TEF encompasses the energy expended in the process of ingesting, absorbing, metabolizing, and storing nutrients from food [Citation8]. Roughly 10% of TDEE is attributed to TEF [Citation44, Citation45], with values varying based on the macronutrient composition of the diet. While the relative magnitude of TEF does not appear to change with energy restriction [Citation46], such dietary restriction involves the consumption of fewer total calories, and therefore decreases the absolute magnitude of TEF [Citation41, Citation46]. NEAT, or energy expended during “non-exercise” movement such as fidgeting or normal daily activities, also decreases with an energy deficit [Citation47]. There is evidence to suggest that spontaneous physical activity, a component of NEAT, is decreased in energy restricted subjects, and may remain suppressed for some time after subjects return to ad libitum feeding [Citation29]. Persistent suppression of NEAT may contribute to weight regain in the post-diet period.
In order to manipulate an individual’s body mass, energy intake must be adjusted based on the individual’s energy expenditure. In the context of weight loss or maintaining a reduced body weight, this process is complicated by the dynamic nature of energy expenditure. In response to weight loss, reductions in TDEE, BMR, EAT, NEAT, and TEF are observed. Due to adaptive thermogenesis, TDEE is lowered to an extent that exceeds the magnitude predicted by losses in body mass. Further, research indicates that adaptive thermogenesis and decreased energy expenditure persist after the active weight loss period, even in subjects who have maintained a reduced body weight for over a year [Citation14, Citation48]. These changes serve to minimize the energy deficit, attenuate further loss of body mass, and promote weight regain in weight-reduced subjects.
Adaptations in mitochondrial efficiency
A series of chemical reactions must take place to derive ATP from stored and ingested energy substrates. In aerobic metabolism, this process involves the movement of protons across the inner mitochondrial membrane. When protons are transported by ATP synthase, ATP is produced. Protons may also leak across the inner membrane by way of uncoupling proteins (UCPs) [Citation49]. In this “uncoupled respiration”, energy substrate oxidation and oxygen consumption occur, but the process does not yield ATP. Proton leak is a significant contributor to energy expenditure, accounting for roughly 20-30% of BMR in rats [Citation50–Citation52].
In the condition of calorie restriction, proton leak is reduced [Citation16–Citation19]. Uncoupling protein-1 and UCP-3, the primary UCPs of brown adipose tissue (BAT) and skeletal muscle [Citation53], are of particular interest due to their potentially significant roles in energy expenditure and uncoupled thermogenesis. Skeletal muscle’s large contribution to energy expenditure [Citation54] has directed attention toward literature reporting decreases in UCP-3 expression in response to energy restriction [Citation55, Citation56]. Decreased UCP-3 expression could potentially play a role in decreasing energy expenditure, and UCP-3 expression has been negatively correlated with body mass index and positively correlated with metabolic rate during sleep [Citation57]. Despite these correlations, more research is needed to determine the function and physiological relevance of UCP-3 [Citation58], as contradictory findings regarding UCP-3 and weight loss have been reported [Citation18].
Uncoupling Protein-1 appears to play a pivotal role in the uncoupled thermogenic activity of BAT [Citation59]. Energy restriction has been shown to decrease BAT activation [Citation60] and UCP-1 expression [Citation61], indicating an increase in metabolic efficiency. Along with UCP-1 expression, thyroid hormone and leptin affect the magnitude of uncoupled respiration in BAT. Thyroid hormone (TH) and leptin are associated with increased BAT activation, whereas glucocorticoids oppose the BAT-activating function of leptin [Citation59]. Evidence indicates that TH plays a prominent role in modulating the magnitude of proton leak [Citation53], with low TH levels associated with decreased proton leak [Citation62]. The endocrine response to energy restriction, including increased cortisol and decreased TH and leptin [Citation1, Citation10, Citation28–Citation31], could potentially play a regulatory role in uncoupled respiration in BAT. It is not clear if decreases in proton leak and UCP expression persist until weight reverts to baseline, but there is evidence to suggest a persistent adaptation [Citation19, Citation55, Citation56], which mirrors the persistent downregulation of TH and leptin [Citation32, Citation33].
Changes observed in proton leak, UCP expression, and circulating hormones appear to influence metabolic efficiency and energy expenditure. In the context of energy restriction, the observed changes are likely to make weight loss increasingly challenging and promote weight regain. It has been reported that females have more BAT than males [Citation63], and that energy-restricted female rats see greater decreases in BAT mass and UCP-1 than males [Citation64], indicating a potential sex-related difference in uncoupled respiration during weight loss. Subjects identified as “diet-resistant” show decreased proton leak and UCP-3 expression compared to “diet-responsive” subjects during maintenance of a reduced bodyweight [Citation65]. More research is needed to determine if these differential responses to hypocaloric diets make sustained weight loss more difficult for females and certain predisposed “diet-resistant” individuals. While future research may improve our understanding of the magnitude and relative importance of mitochondrial adaptations to energy restriction, current evidence suggests that increased mitochondrial efficiency, and a decline in uncoupled respiration, might serve to decrease the energy deficit in hypocaloric conditions, making weight maintenance and further weight reduction more challenging.
Practical applications for weight loss in athletes
Hypocaloric diets induce a number of adaptations that serve to prevent further weight loss and conserve energy. It is likely that the magnitude of these adaptations are proportional to the size of the energy deficit, so it is recommended to utilize the smallest possible deficit that yields appreciable weight loss. This may decrease the rate of weight loss, but attenuate unfavorable adaptations that challenge successful reduction of fat mass. Weight reduction should be viewed as a stepwise process in this context; as weight loss begins to plateau, energy intake or expenditure should be adjusted to “re-open” the energy deficit. Large caloric deficits are also likely to induce greater losses of LBM [Citation66, Citation67] and compromise athletic performance and recovery [Citation68, Citation69], which are of critical importance to athletes. Participation in a structured resistance training program [Citation34] and sufficient protein intake [Citation35–Citation37] are also likely to attenuate losses in LBM. Additionally, high protein diets (≥25%PRO) are associated with increased satiety and thermogenesis, making them a better option for the calorie-restricted athlete [Citation70].
In the world of physique sports, periodic “refeeding” has become common in periods of extended dieting. A refeed consists of a brief overfeeding period in which caloric intake is raised slightly above maintenance levels, and the increase in caloric intake is predominantly achieved by increasing carbohydrate consumption. While studies have utilized refeeding protocols that last three days [Citation71, Citation72], physique athletes such as bodybuilders and figure competitors often incorporate 24-hour refeeds, once or twice per week. The proposed goal of periodic refeeding is to temporarily increase circulating leptin and stimulate the metabolic rate. There is evidence indicating that leptin is acutely responsive to short-term overfeeding [Citation72], is highly correlated with carbohydrate intake [Citation71, Citation73], and that pharmacological administration of leptin reverses many unfavorable adaptations to energy restriction [Citation33]. While interventions have shown acute increases in leptin from short-term carbohydrate overfeeding, the reported effect on metabolic rate has been modest [Citation71]. Dirlewanger et al. reported a 7% increase in TDEE; this increase amounts to approximately 138 kilocalories of additional energy expenditure, of which 36 kilocalories can be attributed to the thermic effect of carbohydrate intake [Citation71]. More research is needed to determine if acute bouts of refeeding are an efficacious strategy for improving weight loss success during prolonged hypocaloric states. A theoretical model of metabolic adaptation and potential strategies to attenuate adaptations is presented in Figure .
Figure 2 A theoretical model of metabolic adaptation and potential strategies to attenuate adaptations. A/A/T hormones = Anabolic, Anorexigenic, and Thermogenic hormones; O/C hormones = Orexigenic and Catabolic hormones. Dotted lines represent inhibition.
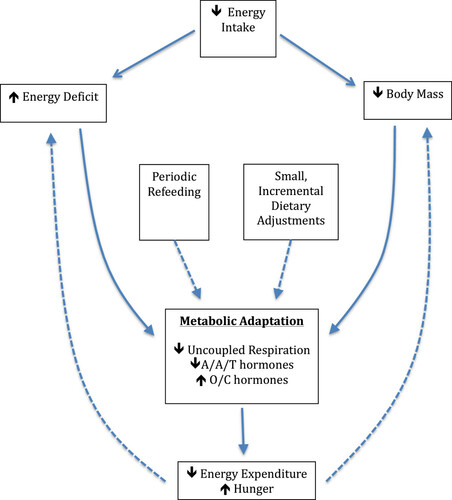
In the period shortly after cessation of a restrictive diet, body mass often reverts toward pre-diet values [Citation29, Citation74, Citation75]. This body mass is preferentially gained as fat mass, in a phenomenon known as post-starvation obesity [Citation29]. While many of the metabolic adaptations to weight loss persist, a dramatic increase in energy intake results in rapid accumulation of fat mass. It is common for individuals to “overshoot” their baseline level of body fat, and leaner individuals (including many athletes) may be more susceptible to overshooting than obese individuals [Citation74, Citation75]. In such a situation, the individual may increase body fat beyond baseline levels, yet retain a metabolic rate that has yet to fully recover. There is evidence to suggest that adipocyte hyperplasia may occur early in the weight-regain process [Citation76], and that repeated cycles of weight loss and regain by athletes in sports with weight classes are associated with long-term weight gain [Citation77]. Therefore, athletes who aggressively diet for a competitive season and rapidly regain weight may find it more challenging to achieve optimal body composition in subsequent seasons.
To avoid rapid fat gain following the cessation of a diet, “reverse dieting” has also become popular among physique athletes. Such a process involves slowly increasing caloric intake in a stepwise fashion. In theory, providing a small caloric surplus might help to restore circulating hormone levels and energy expenditure toward pre-diet values, while closely matching energy intake to the recovering metabolic rate in an effort to reduce fat accretion. Ideally, such a process would eventually restore circulating hormones and metabolic rate to baseline levels while avoiding rapid fat gain. While anecdotal reports of successful reverse dieting have led to an increase in its popularity, research is needed to evaluate its efficacy.
Limitations
Although there is a substantial body of research on metabolic adaptations to weight loss, the majority of the research has utilized animal models or subjects that are sedentary and overweight/obese. Accordingly, the current article is limited by the need to apply this data to an athletic population. If the adaptations described in obese populations serve to conserve energy and attenuate weight loss as a survival mechanism, one might speculate that the adaptations may be further augmented in a leaner, more highly active population. Another limitation is the lack of research on the efficacy of periodic refeeding or reverse dieting in prolonged weight reduction, or in the maintenance of a reduced bodyweight. Until such research is available, these anecdotal methods can only be evaluated from a mechanistic and theoretical viewpoint.
Conclusion
Weight loss is a common practice in a number of sports. Whether the goal is a higher strength-to-mass ratio, improved aesthetic presentation, or more efficient locomotion, optimizing body composition is advantageous to a wide variety of athletes. As these athletes create an energy deficit and achieve lower body fat levels, their weight loss efforts will be counteracted by a number of metabolic adaptations that may persist throughout weight maintenance. Changes in energy expenditure, mitochondrial efficiency, and circulating hormone concentrations work in concert to attenuate further weight loss and promote the restoration of baseline body mass. Athletes must aim to minimize the magnitude of these adaptations, preserve LBM, and adequately fuel performance and recovery during weight reduction. To accomplish these goals, it is recommended to approach weight loss in a stepwise, incremental fashion, utilizing small energy deficits to ensure a slow rate of weight loss. Participation in a structured resistance training program and adequate protein intake are also imperative. More research is needed to verify the efficacy of periodic refeeding and reverse dieting in supporting prolonged weight reduction and attenuating post-diet fat accretion.
Abbreviations
| BAT | = | Brown adipose tissue |
| BF% | = | Body fat percentage |
| BMR | = | Basal metabolic rate |
| EAT | = | Exercise activity thermogenesis |
| LBM | = | Lean body mass |
| NEAT | = | Non-exercise activity thermogenesis |
| NREE | = | Non-resting energy expenditure |
| REE | = | Resting energy expenditure |
| TDEE | = | Total daily energy expenditure |
| TEF | = | Thermic effect of food |
| TH | = | Thyroid Hormone |
| T3 | = | Triiodothyronine |
| UCP | = | Uncoupling protein. |
Authors’ original file for figure 1
Download PDF (35.6 KB)Authors’ original file for figure 2
Download PDF (6.3 MB)Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
ETT conceived of the review topic and drafted the manuscript. AES conceived, drafted and revised the manuscript. LEN helped to draft and revise the manuscript. All authors read and approved the final manuscript.
Related Research Data
References
- RossowLMFukudaDHFahsCALoennekeJPStoutJRNatural bodybuilding competition preparation and recovery: a 12-month case studyInt J Sports Physiol Perform20138 582 592 23412685
- MaestuJEliakimAJurimaeJValterIJurimaeTAnabolic and catabolic hormones and energy balance of the male bodybuilders during the preparation for the competitionJ Strength Cond Res201024 1074 1081 https://doi.org/10.1519/JSC.0b013e3181cb6fd3 20300017
- YoonJPhysiological profiles of elite senior wrestlersSports Med200232 225 233 https://doi.org/10.2165/00007256-200232040-00002 11929352
- FranchiniEDel VecchioFBMatsushigueKAArtioliGGPhysiological profiles of elite judo athletesSports Med201141 147 166 https://doi.org/10.2165/11538580-000000000-00000 21244106
- DeutzRCBenardotDMartinDECodyMMRelationship between energy deficits and body composition in elite female gymnasts and runnersMed Sci Sports Exerc200032 659 668 1:STN:280:DC%2BD3c7ptlaktA%3D%3D https://doi.org/10.1097/00005768-200003000-00017 10731010
- WilmoreJHBrownCHDavisJABody physique and composition of the female distance runnerAnn N Y Acad Sci1977301 764 776 1:STN:280:DyaE1c%2FlvFSntA%3D%3D https://doi.org/10.1111/j.1749-6632.1977.tb38245.x 270950
- DullooAGJacquetJAdaptive reduction in basal metabolic rate in response to food deprivation in humans: a role for feedback signals from fat storesAm J Clin Nutr199868 599 606 1:CAS:528:DyaK1cXlvFWnsbo%3D 9734736
- MacleanPSBergouignanACornierMAJackmanMRBiology’s response to dieting: the impetus for weight regainAm J Physiol Regul Integr Comp Physiol2011301 R581 R600 3174765 1:CAS:528:DC%2BC3MXht1SkurjF https://doi.org/10.1152/ajpregu.00755.2010 21677272
- MacLeanPSHigginsJAJackmanMRJohnsonGCFleming-ElderBKWyattHRMelansonELHillJOPeripheral metabolic responses to prolonged weight reduction that promote rapid, efficient regain in obesity-prone ratsAm J Physiol Regul Integr Comp Physiol2006290 R1577 R1588 1:CAS:528:DC%2BD28XlvVeku70%3D https://doi.org/10.1152/ajpregu.00810.2005 16455763
- MaestuJJurimaeJValterIJurimaeTIncreases in ghrelin and decreases in leptin without altering adiponectin during extreme weight loss in male competitive bodybuildersMetabolism200857 221 225 1:CAS:528:DC%2BD1cXlvVGnsg%3D%3D https://doi.org/10.1016/j.metabol.2007.09.004 18191052
- LichtmanSWPisarskaKBermanERPestoneMDowlingHOffenbacherEWeiselHHeshkaSMatthewsDEHeymsfieldSBDiscrepancy between self-reported and actual caloric intake and exercise in obese subjectsN Engl J Med1992327 1893 1898 1:STN:280:DyaK3s%2Fos1Kntw%3D%3D https://doi.org/10.1056/NEJM199212313272701 1454084
- GarriguetDUnder-reporting of energy intake in the Canadian community health surveyHealth Rep200819 37 45 19226926
- DoucetESt-PierreSAlmerasNDespresJPBouchardCTremblayAEvidence for the existence of adaptive thermogenesis during weight lossBr J Nutr200185 715 723 1:CAS:528:DC%2BD3MXlsVartr8%3D https://doi.org/10.1079/BJN2001348 11430776
- RosenbaumMHirschJGallagherDALeibelRLLong-term persistence of adaptive thermogenesis in subjects who have maintained a reduced body weightAm J Clin Nutr200888 906 912 1:CAS:528:DC%2BD1cXht1Gks7jN 18842775
- RosenbaumMLeibelRLAdaptive thermogenesis in humansInt J Obes201034 Suppl 1 S47 S55 https://doi.org/10.1038/ijo.2010.184
- AsamiDKMcDonaldRBHagopianKHorwitzBAWarmanDHsiaoAWardenCRamseyJJEffect of aging, caloric restriction, and uncoupling protein 3 (UCP3) on mitochondrial proton leak in miceExp Gerontol200843 1069 1076 2614627 1:CAS:528:DC%2BD1cXhsVWms7nE https://doi.org/10.1016/j.exger.2008.09.010 18852040
- BevilacquaLRamseyJJHagopianKWeindruchRHarperMEEffects of short- and medium-term calorie restriction on muscle mitochondrial proton leak and reactive oxygen species productionAm J Physiol Regul Integr Comp Physiol2004286 E852 E861 1:CAS:528:DC%2BD2cXkt1emu7o%3D
- BevilacquaLRamseyJJHagopianKWeindruchRHarperMELong-term caloric restriction increases UCP3 content but decreases proton leak and reactive oxygen species production in rat skeletal muscle mitochondriaAm J Physiol Endocrinol Metab2005289 E429 E438 1:CAS:528:DC%2BD2MXhtVWksb3F https://doi.org/10.1152/ajpendo.00435.2004 15886224
- HagopianKHarperMERamJJHumbleSJWeindruchRRamseyJJLong-term calorie restriction reduces proton leak and hydrogen peroxide production in liver mitochondriaAm J Physiol Endocrinol Metab2005288 E674 E684 1:CAS:528:DC%2BD2MXjtlyjsr8%3D https://doi.org/10.1152/ajpendo.00382.2004 15562252
- KimBThyroid hormone as a determinant of energy expenditure and the basal metabolic rateThyroid200818 141 144 1:CAS:528:DC%2BD1cXitVKnt7g%3D https://doi.org/10.1089/thy.2007.0266 18279014
- MargeticSGazzolaCPeggGGHillRALeptin: a review of its peripheral actions and interactionsInt J Obes Relat Metab Disord200226 1407 1433 1:CAS:528:DC%2BD38XoslGlsbc%3D https://doi.org/10.1038/sj.ijo.0802142 12439643
- RooyackersOENairKSHormonal regulation of human muscle protein metabolismAnnu Rev Nutr199717 457 485 1:CAS:528:DyaK2sXkvFGmtLk%3D https://doi.org/10.1146/annurev.nutr.17.1.457 9240936
- StrohackerKMcCafferyJMMacleanPSWingRRAdaptations of leptin, ghrelin or insulin during weight loss as predictors of weight regain: a review of current literatureInt J Obes2013 1 9 http://www.nature.com/ijo/journal/vaop/ncurrent/full/ijo2013118a.html
- AriyasuHTakayaKTagamiTOgawaYHosodaKAkamizuTSudaMKohTNatsuiKToyookaSAriyasuHTakayaKTagamiTOgawaYHosodaKAkamizuTSudaMKohTNatsuiKToyookaSShirakamiGUsuiTShimatsuADoiKHosodaHKojimaMKangawaKNakaoKStomach is a major source of circulating ghrelin, and feeding state determines plasma ghrelin-like immunoreactivity levels in humansJ Clin Endocrinol Metab200186 4753 4758 1:CAS:528:DC%2BD3MXptVars70%3D https://doi.org/10.1210/jcem.86.10.7885 11600536
- De MaddalenaCVodoSPetroniAAloisiAMImpact of testosterone on body fat compositionJ Cell Physiol2012227 3744 3748 1:CAS:528:DC%2BC38Xht1CgtrfN https://doi.org/10.1002/jcp.24096 22495883
- SimmonsPSMilesJMGerichJEHaymondMWIncreased proteolysis. An effect of increases in plasma cortisol within the physiologic rangeJ Clin Invest198473 412 420 425032 1:CAS:528:DyaL2cXhtFGhs7k%3D https://doi.org/10.1172/JCI111227 6365973
- ZakrzewskaKECusinISainsburyARohner-JeanrenaudFJeanrenaudBGlucocorticoids as counterregulatory hormones of leptin: toward an understanding of leptin resistanceDiabetes199746 717 719 1:CAS:528:DyaK2sXitFKqsLc%3D https://doi.org/10.2337/diab.46.4.717 9075817
- HagmarMBerglundBBrismarKHirschbergALBody composition and endocrine profile of male Olympic athletes striving for leannessClin J Sport Med201323 197 201 https://doi.org/10.1097/JSM.0b013e31827a8809 23275346
- WeyerCWalfordRLHarperITMilnerMMacCallumTTataranniPARavussinEEnergy metabolism after 2 y of energy restriction: the biosphere 2 experimentAm J Clin Nutr200072 946 953 1:CAS:528:DC%2BD3cXnt1amt74%3D 11010936
- WitbrachtMGLaugeroKDVan LoanMDAdamsSHKeimNLPerformance on the Iowa gambling task is related to magnitude of weight loss and salivary cortisol in a diet-induced weight loss intervention in overweight womenPhysiol Behav2012106 291 297 1:CAS:528:DC%2BC38XksFensb0%3D https://doi.org/10.1016/j.physbeh.2011.04.035 21565212
- TomiyamaAJMannTVinasDHungerJMDejagerJTaylorSELow calorie dieting increases cortisolPsychosom Med201072 357 364 2895000 1:CAS:528:DC%2BC3cXlvVehsb0%3D https://doi.org/10.1097/PSY.0b013e3181d9523c 20368473
- SumithranPPrendergastLADelbridgeEPurcellKShulkesAKriketosAProiettoJLong-term persistence of hormonal adaptations to weight lossN Engl J Med2011365 1597 1604 1:CAS:528:DC%2BC3MXhsVSrsrbP https://doi.org/10.1056/NEJMoa1105816 22029981
- RosenbaumMGoldsmithRBloomfieldDMagnanoAWeimerLHeymsfieldSGallagherDMayerLMurphyELeibelRLLow-dose leptin reverses skeletal muscle, autonomic, and neuroendocrine adaptations to maintenance of reduced weightJ Clin Invest2005115 3579 3586 1297250 1:CAS:528:DC%2BD2MXhtlSqtLjM https://doi.org/10.1172/JCI25977 16322796
- BrynerRWUllrichIHSauersJDonleyDHornsbyGKolarMYeaterREffects of resistance vs. aerobic training combined with an 800 calorie liquid diet on lean body mass and resting metabolic rateJ Am Coll Nutr199918 115 121 1:STN:280:DyaK1M3hvFGhsg%3D%3D https://doi.org/10.1080/07315724.1999.10718838 10204826
- MettlerSMitchellNTiptonKDIncreased protein intake reduces lean body mass loss during weight loss in athletesMed Sci Sports Exerc201042 326 337 1:CAS:528:DC%2BC3cXpsVSntg%3D%3D https://doi.org/10.1249/MSS.0b013e3181b2ef8e 19927027
- LaymanDKBoileauRAEricksonDJPainterJEShiueHSatherCChristouDDA reduced ratio of dietary carbohydrate to protein improves body composition and blood lipid profiles during weight loss in adult womenJ Nutr2003133 411 417 1:CAS:528:DC%2BD3sXhtV2ntb4%3D 12566476
- BoppMJHoustonDKLenchikLEasterLKritchevskySBNicklasBJLean mass loss is associated with low protein intake during dietary-induced weight loss in postmenopausal womenJ Am Diet Assoc2008108 1216 1220 3665330 https://doi.org/10.1016/j.jada.2008.04.017 18589032
- RavussinEBurnandBSchutzYJequierEEnergy expenditure before and during energy restriction in obese patientsAm J Clin Nutr198541 753 759 1:STN:280:DyaL2M7ntFyjtg%3D%3D 3984927
- LeibelRLRosenbaumMHirschJChanges in energy expenditure resulting from altered body weightN Engl J Med1995332 621 628 1:STN:280:DyaK2M7ksl2gsw%3D%3D https://doi.org/10.1056/NEJM199503093321001 7632212
- WeigleDSContribution of decreased body mass to diminished thermic effect of exercise in reduced-obese menInt J Obes198812 567 578 1:STN:280:DyaL1M7ntlGmuw%3D%3D 3235273
- WeigleDSBrunzellJDAssessment of energy expenditure in ambulatory reduced-obese subjects by the techniques of weight stabilization and exogenous weight replacementInt J Obes199014 Suppl 1 69 77 2228420 discussion 77–81
- DoucetEImbeaultPSt-PierreSAlmerasNMauriegePDespresJPBouchardCTremblayAGreater than predicted decrease in energy expenditure during exercise after body weight loss in obese menClin Sci2003105 89 95 https://doi.org/10.1042/CS20020252 12617720
- RosenbaumMVandenborneKGoldsmithRSimoneauJAHeymsfieldSJoanisseDRHirschJMurphyEMatthewsDSegalKRLeibelRLEffects of experimental weight perturbation on skeletal muscle work efficiency in human subjectsAm J Physiol Regul Integr Comp Physiol2003285 R183 192 1:CAS:528:DC%2BD3sXlsFGhs7k%3D https://doi.org/10.1152/ajpregu.00474.2002 12609816
- TappyLThermic effect of food and sympathetic nervous system activity in humansReprod Nutr Dev199636 391 397 1:STN:280:DyaK2s%2Fjt1CktA%3D%3D https://doi.org/10.1051/rnd:19960405 8878356
- RavussinELilliojaSAndersonTEChristinLBogardusCDeterminants of 24-hour energy expenditure in man. Methods and results using a respiratory chamberJ Clin Invest198678 1568 1578 423919 1:STN:280:DyaL2s%2FlvVWisw%3D%3D https://doi.org/10.1172/JCI112749 3782471
- MilesCWWongNPRumplerWVConwayJEffect of circadian variation in energy expenditure, within-subject variation and weight reduction on thermic effect of foodEur J Clin Nutr199347 274 284 1:STN:280:DyaK3s3lvFWjsw%3D%3D 8491165
- LevineJANon-exercise activity thermogenesis (NEAT)Best Pract Res Clin Endocrinol Metab200216 679 702 https://doi.org/10.1053/beem.2002.0227 12468415
- LeibelRLHirschJDiminished energy requirements in reduced-obese patientsMetabolism198433 164 170 1:STN:280:DyaL2c7hs12gug%3D%3D https://doi.org/10.1016/0026-0495(84)90130-6 6694559
- JastrochMDivakaruniASMookerjeeSTrebergJRBrandMDMitochondrial proton and electron leaksEssays Biochem201047 53 67 3122475 1:CAS:528:DC%2BC3cXpvFKkt78%3D https://doi.org/10.1042/bse0470053 20533900
- RolfeDFBrandMDContribution of mitochondrial proton leak to skeletal muscle respiration and to standard metabolic rateAm J Physiol1996271 C1380 1389 1:CAS:528:DyaK28XmsF2msLg%3D 8897845
- RolfeDFBrownGCCellular energy utilization and molecular origin of standard metabolic rate in mammalsPhysiol Rev199777 731 758 1:CAS:528:DyaK2sXltlWlurg%3D 9234964
- RolfeDFNewmanJMBuckinghamJAClarkMGBrandMDContribution of mitochondrial proton leak to respiration rate in working skeletal muscle and liver and to SMRAm J Physiol1999276 C692 699 1:CAS:528:DyaK1MXit1Srtbk%3D 10069997
- ThrushABDentRMcPhersonRHarperMEImplications of mitochondrial uncoupling in skeletal muscle in the development and treatment of obesityFEBS J2013280 5015 5029 1:CAS:528:DC%2BC3sXhsFOgsr3P https://doi.org/10.1111/febs.12399 23786211
- ZurloFLarsonKBogardusCRavussinESkeletal muscle metabolism is a major determinant of resting energy expenditureJ Clin Invest199086 1423 1427 296885 1:CAS:528:DyaK3cXmt1yguro%3D https://doi.org/10.1172/JCI114857 2243122
- EsterbauerHOberkoflerHDallingerGBrebanDHellEKremplerFPatschWUncoupling protein-3 gene expression: reduced skeletal muscle mRNA in obese humans during pronounced weight lossDiabetologia199942 302 309 1:CAS:528:DyaK1MXhsVyisrc%3D https://doi.org/10.1007/s001250051155 10096782
- Vidal-PuigARosenbaumMConsidineRCLeibelRLDohmGLLowellBBEffects of obesity and stable weight reduction on UCP2 and UCP3 gene expression in humansObes Res19997 133 140 1:STN:280:DyaK1M3gtVeksA%3D%3D https://doi.org/10.1002/j.1550-8528.1999.tb00694.x 10102249
- SchrauwenPXiaJBogardusCPratleyRERavussinESkeletal muscle uncoupling protein 3 expression is a determinant of energy expenditure in Pima IndiansDiabetes199948 146 149 1:CAS:528:DyaK1MXhvFSjtQ%3D%3D https://doi.org/10.2337/diabetes.48.1.146 9892236
- HarperMEDentRMBezaireVAntoniouAGauthierAMonemdjouSMcPhersonRUCP3 and its putative function: consistencies and controversiesBiochem Soc Trans200129 768 773 1:CAS:528:DC%2BD3MXpt1Crsb4%3D https://doi.org/10.1042/bst0290768 11709072
- CannonBNedergaardJBrown adipose tissue: function and physiological significancePhysiol Rev200484 277 359 1:CAS:528:DC%2BD2cXotl2qsQ%3D%3D https://doi.org/10.1152/physrev.00015.2003 14715917
- RothwellNJStockMJEffect of chronic food restriction on energy balance, thermogenic capacity, and brown-adipose-tissue activity in the ratBiosci Rep19822 543 549 1:CAS:528:DyaL38XlvVSqsro%3D https://doi.org/10.1007/BF01314214 7139069
- YoungJBSavilleERothwellNJStockMJLandsbergLEffect of diet and cold exposure on norepinephrine turnover in brown adipose tissue of the ratJ Clin Invest198269 1061 1071 370170 1:CAS:528:DyaL38XktVWjtbY%3D https://doi.org/10.1172/JCI110541 7068845
- HarperMEBrandMDThe quantitative contributions of mitochondrial proton leak and ATP turnover reactions to the changed respiration rates of hepatocytes from rats of different thyroid statusJ Biol Chem1993268 14850 14860 1:CAS:528:DyaK3sXkvVKrtLY%3D 8392060
- CypessAMLehmanSWilliamsGTalIRodmanDGoldfineABKuoFCPalmerELTsengYHDoriaACypessAMLehmanSWilliamsGTalIRodmanDGoldfineABKuoFCPalmerELTsengYHDoriaAKolodnyGMKahnCRIdentification and importance of brown adipose tissue in adult humansN Engl J Med2009360 1509 1517 2859951 1:CAS:528:DC%2BD1MXltlSqsro%3D https://doi.org/10.1056/NEJMoa0810780 19357406
- ValleACatala-NiellAColomBGarcia-PalmerFJOliverJRocaPSex-related differences in energy balance in response to caloric restrictionAm J Physiol Endocrinol Metab2005289 E15 22 1:CAS:528:DC%2BD2MXmvVertbo%3D https://doi.org/10.1152/ajpendo.00553.2004 15701677
- HarperMEDentRMonemdjouSBezaireVVan WyckLWellsGKavaslarGNGauthierATessonFMcPhersonRDecreased mitochondrial proton leak and reduced expression of uncoupling protein 3 in skeletal muscle of obese diet-resistant womenDiabetes200251 2459 2466 1:CAS:528:DC%2BD38XlvFKiu78%3D https://doi.org/10.2337/diabetes.51.8.2459 12145158
- ChastonTBDixonJBO’BrienPEChanges in fat-free mass during significant weight loss: a systematic reviewInt J Obes200731 743 750 1:STN:280:DC%2BD2s3kvVOmsQ%3D%3D
- GartheIRaastadTRefsnesPEKoivistoASundgot-BorgenJEffect of two different weight-loss rates on body composition and strength and power-related performance in elite athletesInt J Sport Nutr Exerc Metab201121 97 104 1:CAS:528:DC%2BC3MXmtF2jurg%3D 21558571
- RodriguezNRDi MarcoNMLangleyS American Dietetic A, Dietitians of C, American College of Sports M American College of Sports Medicine position stand. Nutrition and athletic performanceMed Sci Sports Exerc200941 709 731 https://doi.org/10.1249/MSS.0b013e31890eb86 19225360
- BurkeLMLoucksABBroadNEnergy and carbohydrate for training and recoveryJ Sports Sci200624 675 685 https://doi.org/10.1080/02640410500482602 16766497
- Paddon-JonesDWestmanEMattesRDWolfeRRAstrupAWesterterp-PlantengaMProtein, weight management, and satietyAm J Clin Nutr200887 1558S 1561S 1:CAS:528:DC%2BD1cXmtFehtrw%3D 18469287
- DirlewangerMdi VettaVGuenatEBattilanaPSeematterGSchneiterPJequierETappyLEffects of short-term carbohydrate or fat overfeeding on energy expenditure and plasma leptin concentrations in healthy female subjectsInt J Obes Relat Metab Disord200024 1413 1418 1:CAS:528:DC%2BD3cXosFymsr0%3D https://doi.org/10.1038/sj.ijo.0801395 11126336
- Chin-ChanceCPolonskyKSSchoellerDATwenty-four-hour leptin levels respond to cumulative short-term energy imbalance and predict subsequent intakeJ Clin Endocrinol Metab200085 2685 2691 1:CAS:528:DC%2BD3cXnslaiu7o%3D 10946866
- JenkinsABMarkovicTPFleuryACampbellLVCarbohydrate intake and short-term regulation of leptin in humansDiabetologia199740 348 351 1:CAS:528:DyaK2sXhs1CktLw%3D https://doi.org/10.1007/s001250050686 9084976
- DullooAGJacquetJGirardierLPoststarvation hyperphagia and body fat overshooting in humans: a role for feedback signals from lean and fat tissuesAm J Clin Nutr199765 717 723 1:CAS:528:DyaK2sXhs1Gmu78%3D 9062520
- DullooAGJacquetJMontaniJPHow dieting makes some fatter: from a perspective of human body composition autoregulationProc Nutr Soc201271 379 389 https://doi.org/10.1017/S0029665112000225 22475574
- JackmanMRSteigAHigginsJAJohnsonGCFleming-ElderBKBessesenDHMacLeanPSWeight regain after sustained weight reduction is accompanied by suppressed oxidation of dietary fat and adipocyte hyperplasiaAm J Physiol Regul Integr Comp Physiol2008294 R1117 1129 1:CAS:528:DC%2BD1cXkvVyks7s%3D https://doi.org/10.1152/ajpregu.00808.2007 18287221
- SaarniSERissanenASarnaSKoskenvuoMKaprioJWeight cycling of athletes and subsequent weight gain in middleageInt J Obes200630 1639 1644 1:STN:280:DC%2BD28njsVGrtA%3D%3D https://doi.org/10.1038/sj.ijo.0803325
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.