Abstract
Background
Carnosine is a dipeptide that improves exercise performance. The carnosine synthesis mechanism through carnosine and β-alanine ingestion remains unclear. Therefore, we investigated the tissue distribution of carnosine synthase, ATP-grasp domain-containing protein-1 (ATPGD1) mRNA, and ATPGD1 and carnosine specific dipeptidase (CN1) gene expression profiles in mice that were given carnosine or β-alanine orally.
Methods
ddY mice (7-week-old) were randomly divided into three groups (n = 6 to 8 animals per group) and were orally given 2 g/kg body weight of carnosine, β-alanine, or water. After 15, 30, 60, 120, 180, or 360 min of treatment, the tissues (brain, blood, liver, kidneys, olfactory bulbs, hindleg muscles) were collected. The obtained tissues measured the expression of ATPGD1 and CN1 genes using quantitative PCR methods.
Results
The ATPGD1 gene was expressed in muscle and to a lesser extent in brain. The expression of ATPGD1 in the vastus lateralis muscle increased significantly at 180 min (P = 0.023) after carnosine ingestion and 60 (P = 0.023) and 180 min (P = 0.025) after β-alanine ingestion. Moreover, the carnosine group showed a significantly increased renal expression of the CN1 gene 60 min after ingestion (P = 0.0015).
Conclusions
The ATPGD1 gene showed high expression levels in brain and muscle. The β-alanine or carnosine administration significantly increased ATPGD1 and CN1 expression in mice.
Electronic supplementary material
The online version of this article (doi:10.1186/1550-2783-9-15) contains supplementary material, which is available to authorized users.
Background
Carnosine (β-alanyl-L-histidine) is a dipeptide abundant in mammalian skeletal muscles [Citation1, Citation2]. Various physiological actions have been ascribed to carnosine in muscle, including acting as an antioxidant [Citation3], regulating Ca2+ sensitivity [Citation4], protecting proteins against glycation by acting as a sacrificial peptide [Citation5], and preventing the formation of protein–protein cross links by reacting with protein-carbonyl groups [Citation6]. Primarily, carnosine with pH buffering capacity is widely used in the field of sports nutrition [Citation7]. Because the dissociation exponent (pKa) of carnosine is 6.83 [Citation8, Citation9], it is suggested that carnosine attenuates the reduction in blood pH by a large amount of H+ originating from the dissociation of lactic acid during strenuous exercise, and suppresses a loss of force [Citation10]. At the same time, muscle carnosine contents are positively correlated with high-intensity exercise performance [Citation11] and fast-twitch muscle fibers [Citation12]. Increase of muscle carnosine predominantly was due to the ingestion of histidine-containing dipeptide (HCD) such as carnosine, anserine (β-alanyl-1-methylhistidine) and balenine (β-alanyl-3-methylhistidine) or β-alanine. Although β-alanine could also be synthesized from the degradation of uracil, there are no reports on the relation between carnosine synthesis and pyrimidine catabolism. So, the majority of the previous research relating to the ergogenic effects of elevated muscle carnosine content via chicken breast extract, high in HCD content or β-alanine supplementation was performed using mice, horses and humans [Citation13–Citation17].
However, ingested carnosine is rapidly degraded by two forms of carnosinase (CN1, EC 3.4.13.20; and CN2, EC 3.4.13.18) [Citation18]. In humans, the CN1 gene is expressed in liver and brain tissue, and the protein is found in serum and brain tissue. Since the human CN1 specifically degrades both carnosine and homocarnosine, carnosine is absent in human blood. Whereas, CN1 in other mammals such as rodents is localized in the kidney, and a considerable amount of carnosine is contained in the blood [Citation19]. CN2, which is also a cytosolic non-specific dipeptidase, does not degrade homocarnosine, and exhibits a rather broad specificity towards various dipeptides. That is, ingestion of β-alanine or carnosine that was degraded by these carnosinases, was increased muscle carnosine and the increase of muscle carnosine may be involved in carnosine synthase. However, the details were not revealed.
Recently, carnosine synthase was purified from chicken pectoral muscle and identified as an ATP-grasp domain-containing protein 1 (ATPGD1) [Citation20]. It has been reported that ATPGD1 synthesizes carnosine using ATP, and the substrate specificity toward β-alanine (carnosine) in the presence of ATP and L-histidine is 14-fold higher than that of γ-aminobutyrate (homocarnosine). To verify that ATPGD1 acts as a carnosine synthase in vivo, we investigated the tissue distribution of ATPGD1 mRNA, and ATPGD1 and CN1 expression profiles in response to carnosine or β-alanine administration using quantitative PCR analysis.
Methods
Oral administration study in mice
Animal experiments were performed in accordance with the guidelines for Animal Experiments at Nippon Meat Packers Inc. and using minimum number of mice that dictated by an ethics committee ( n = 6 or 8). Male SPF-bred ddY (6-week-old) mice were purchased from Japan SLC, Inc. (Shizuoka, Japan). The mice were maintained under specifically controlled environmental conditions, namely, a 12-h light–dark cycle, a temperature of 23°C, and a relative humidity of 50%. At 7 weeks of age, the mice were randomly assigned by body weight into three groups (pre-administration group, n = 8, body weight of 32.5 g; water administration group, n = 6, body weight of 33.4 g; carnosine administration group, n = 6 or 8, body weight of 33.2 g; β-alanine administration group, n = 6, body weight of 34.0 g) and were orally given 2 g/kg body weight of carnosine (Hamari Chemicals Ltd., Osaka, Japan), β-alanine (Wako Pure Chemical Industries, Ltd., Osaka, Japan), or water (control). After 15, 30, 60, 120, 180, or 360 min of treatment, the mice were anesthetized with Forane (Abbott Japan Co. Ltd., Japan) and then the brain, blood, liver, kidneys, olfactory bulbs, soleus muscles and vastus lateralis muscles were collected. The collected tissues were weighed, rapidly frozen with liquid nitrogen, and stored at -80°C until analysis.
Extraction of total RNA
The frozen tissue samples were homogenized in 0.75 ml of Isogen (Nippon Gene Co. Ltd., Tokyo, Japan) and then mixed thoroughly with 0.15 ml of chloroform. The mixture was centrifuged (20,000 × g for 5 min), and then the aqueous phases were collected, and 0.4 ml of isopropanol was added. The precipitated total RNA was recovered and washed with 70% (v/v) ethanol. The purity and concentration of the total RNA thus obtained were confirmed using an Experion electrophoresis system (Bio-Rad Laboratories, Inc., California, USA) and a NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific K. K., Massachusetts, USA).
Construction of gene specific primers
Gene specific primers were designed by using Primer-BLAST (http://www.ncbi.nlm.nih.gov/tools/primer-blast/). The primers used were as follows: for ATPGD1 (NM_134148), forward primer, 5′-CCCTGGCCTTCGACCTCTCTCCAT-3′ and reverse primer, 5′-CGGCACTGGGGCCCATCCTTC-3′ to yield a 164-bp product; for CN1 (NM_177450), forward primer, 5′-TGGTGGCATCCTCAACGAACCA-3′ and reverse primer, 5′-TCCAGGAATTAGGATGTGGCCTGA-3′ to yield an 88-bp product; for β-actin (NM_007393), forward primer, 5′-ATGAGCTGCCTGACGGCCAGGTCATC-3′ and reverse primer, 5′-TGGTACCACCAGACAGCACTGTGTTG-3′ to yield a 192-bp product.
Quantification of mRNA levels
cDNA was synthesized by using a PrimeScript RT reagent Kit with gDNA Eraser (Takara Bio, Inc., Shiga, Japan). The genomic DNA in the RNAs extracted from tissues was eliminated with gDNA Eraser, which were then reverse-transcribed by PrimeScript RT. Each 25 μl of the PCR reaction mix contained a 2 μl template, 0.2 μM of each primer, and 1× ROX Reference Dye II in 1× SYBR Premix Ex Taq II (Takara Bio, Inc.). The reaction was performed at 95°C for 30 s; this was followed by 40 cycles at 95°C for 5 s and at 60°C for 20 s. The fluorescence was measured at the end of the extension step in each cycle. Following cycling, a melt curve analysis was performed after each quantitative PCR to ensure that a single product had been amplified per primer set. The fold-change of the gene expression was calculated using the 2-∆∆Ct method with β-actin as an internal control. Student’s t-test was used (P < 0.05 or P < 0.01) to test statistical significance.
Detection of carnosine in muscle and blood
Vastus lateralis muscle samples were deproteinized with 1 ml of 5% (w/v) sulfosalicylic acid. The samples were centrifuged at 20,000 × g for 5 min, and then the supernatants were filtered with a 0.45-μm filter. Blood samples were dissolved in 1 M perchloric acid (final concentration, 0.3 M) and centrifuged at 20,000 × g for 5 min. KOH (3 M) was added to the supernatants to realize a final concentration of 4.25% v/v. After centrifugation (20,000 × g for 5 min), the obtained supernatants were filtered and applied to a TSKgel ODS-80Ts column (Tosoh Co., Tokyo, Japan) equilibrated with 4% (v/v) acetonitrile, 100 mM sodium 1-pentanesulfonate, and 200 mM ammonium dihydrogen phosphate (pH 2.0). The carnosine was eluted with the same buffer, and absorbance was detected at a wavelength of 214 nm. Statistical analysis was performed with Tukey-Kramer test (P < 0.05 or P < 0.01).
Results
Tissue distribution of ATPGD1 mRNA
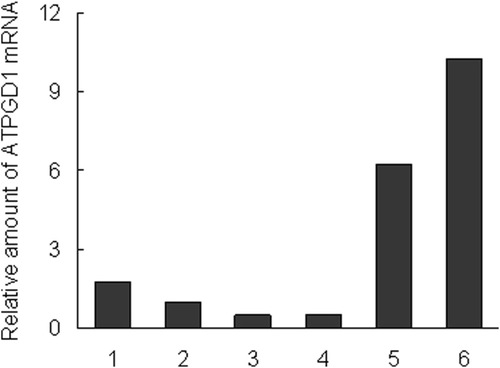
The localization of ATPGD1 mRNA from various tissue samples was investigated by quantitative PCR methods. ATPGD1 genes were detected in muscle, a few in brain, and hardly in liver and kidney. The expression of ATPGD1 was 10.2-fold higher in the vastus lateralis muscle, 6.3-fold higher in the soleus muscle and 1.8-fold higher in the brain than in the olfactory bulbs. In contrast, the expression of ATPGD1 in the liver and kidney was only 50% of that in the olfactory bulbs (Figure ).
Figure 1 Tissue distribution of ATPGD1 mRNA in mice. 1; brain, 2; olfactory bulbs, 3; kidneys, 4; liver, 5; soleus muscles, and 6; vastus lateralis muscles. β-actin gene (Actb) was used as an endogenous control gene.
Carnosine content of blood and muscle
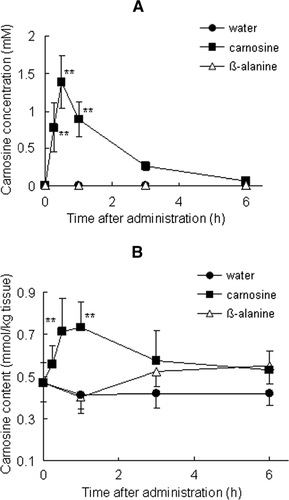
In mice that had ingested carnosine or β-alanine, we measured the carnosine content of the blood and vastus lateralis muscle by using an ODS-80Ts column. The carnosine content of the blood had significantly increased by 15 min after carnosine administration (P < 0.01); it peaked at 30 min (1.4 ± 0.3 mM, P < 0.01) and had nearly disappeared by 6 h (Figure ). No carnosine was detected in the blood of the groups that ingested β-alanine or water. As shown Figure , the carnosine content of the vastus lateralis muscle was 0.47 ± 0.09 mmol/kg tissue before administration. The carnosine level had increased significantly 30 to 60 min after it was administered (0.71 ± 0.15 mmol/kg tissue at 30 min, P < 0.01 and 0.74 ± 0.12 mmol/kg tissue at 60 min, P < 0.01) and then gradually decreased. The carnosine content of muscle in the group that ingested β-alanine did not increase significantly compared with that before administration (P > 0.05).
Figure 2 Time course of carnosine concentration in blood (A), vastus lateralis muscles (B) and following ingestion of carnosine, β-alanine, or water; 2 g/kg body weight carnosine (closed squares), β-alanine (open triangles), or water (closed circles) was orally administered to mice (n = 6–8). Values are means ± SD. Significant differences after administration were analyzed by using Tukey-Kramer test (**P < 0.01).
Gene expression of ATPGD1 and CN1
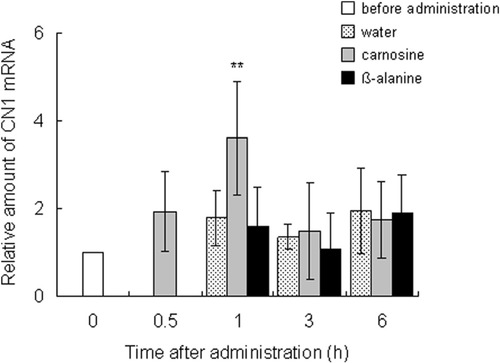
The expression profiles of carnosine synthesis-related genes were measured by using quantitative PCR. The ATPGD1 mRNA level in the vastus lateralis muscle was significantly elevated 3 h after carnosine administration (P = 0.023) and at 1 (P = 0.023) and 3 h (P = 0.025) after β-alanine administration, compared with the level before administration. Expression increased from 2.7 to 3.2 times that before ingestion (Figure ). After carnosine ingestion, the CN1 expression in the kidney peaked at 1 h and was significantly greater (3.6 times, P = 0.0015) than before ingestion (Figure ).
Figure 3 Effect of dietary carnosine and β-alanine on ATPGD1 mRNA expression in the vastus lateralis muscle of male mice; 2 g/kg body weight of carnosine, β-alanine, or water was orally administered to mice (n = 6–8). Values are means ± SD. Significant differences after administration were analyzed by using Student’s t-test (* P < 0.05).
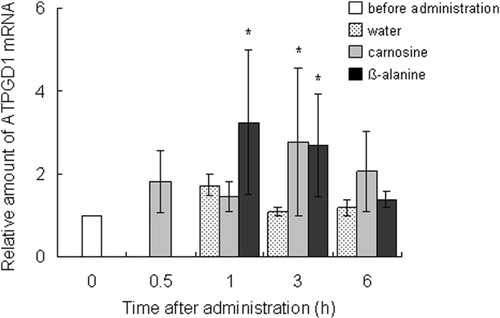
Figure 4 Effect of dietary carnosine and β-alanine on the CN1 mRNA expression in the kidneys of male mice; 2 g/kg body weight of carnosine, β-alanine, or water was orally administered to mice (n = 6–8). Values are means ± SD. Significant differences after administration were analyzed by using Student’s t-test (**P < 0.01).
Discussion
Carnosine synthase have been tried to purify from various sources [Citation21–Citation24] and Drozak et al. purified carnosine synthase from chicken pectoral muscle and the enzyme identified as ATPGD1, which is a member of the ATP-grasp family [Citation20]. This paper was investigated about whether ATPGD1 involved in carnosine synthesis in mice.
Firstly, the tissue distribution of the ATPGD1 gene was investigated. The ATPGD1 gene was expressed more in brain and muscle than in olfactory bulbs, liver and kidney and particularly in the vastus lateralis muscle. The expression of the ATPGD1 gene was 1.6-fold higher than that in the soleus muscle. The carnosine content in the vastus lateralis muscle (0.47 mmol/kg tissue) was higher than in the soleus muscle (0.35 mmol/kg tissue, P = 0.007, data not shown), indicating that the ATPGD1 mRNA level depends on the carnosine content.
Secondly, we investigated the carnosine content and the expression of carnosine synthesis-related genes after the ingestion of carnosine or β-alanine. The carnosine supplementation group increased the carnosine content in blood and muscle and the expression of CN1 in the kidneys. Carnosine was injected into the tail vein of proton-coupled oligopeptide transporter PEPT2 knockout mice and the kidney/plasma concentration ratio of carnosine in the PEPT2 null mice was one-sixth that in wild-type [Citation25]. Thus, it was considered that the ingested carnosine was eliminated from the serum by filtration into the urine and reabsorption into the kidney, and the reabsorbed carnosine increased the expression of CN1 in the kidney and would be hydrolyzed to β-alanine. Carnosine and β-alanine administration increased the ATPGD1 gene levels in the vastus lateralis muscles. This suggests that the hydrolyzed β-alanine in kidney increased ATPGD1 gene expression. Recently, Baguet et al. investigated the expression of ATPGD1 mRNA in human skeletal muscle. Twenty omnivorous subjects were randomly divided into a vegetarian and a mixed diet group, and took part in a five-week sprint training intervention (2–3 times per week). The ATPGD1 mRNA expression in the vegetarian diet group was decreased to 60 % (P = 0.023) by five weeks of sprint training [Citation26]. This is consistent with our result showing that β-alanine is an important factor in ATPGD1 expression.
Chronic chicken breast extract or β-alanine supplementation leads to improved performance in high-intensity exercise [Citation27, Citation28]. However, the loading of carnosine takes at least several weeks [Citation29], in contrast to the initial loading phase of one week for creatine [Citation30]. This paper suggests that ATPGD1 acts as a carnosine synthase in mice, and provides new insights to determine efficient muscle carnosine loading.
Conclusions
The present study shows that the ATPGD1 mRNA in mice was expressed highly in brain and muscle, moderately in olfactory bulbs, scarcely in liver and kidneys, and approximately 67 mg of β-alanine or carnosine administration in mice significantly increased ATPGD1 and CN1 expression.
Authors’ original file for figure 1
Download GIF Image (9.7 KB){kind=link}
Authors’ original file for figure 2
Download GIF Image (34.9 KB){kind=link}
Authors’ original file for figure 3
Download GIF Image (17.3 KB){kind=link}
Authors’ original file for figure 4
Download GIF Image (18.2 KB){kind=link}
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
TM is the principal investigator of the project. MS, HM, YT and FM designed the study; MS and HM collected the data; YT and FM conducted data analysis; TM, MS and HM wrote the manuscript. All authors have read and approved the final manuscript.
References
- CrushKGCarnosine and related substances in animal tissuesComp Biochem Physiol197034 3 30 1:CAS:528:DyaE3cXkt1Wns74%3D https://doi.org/10.1016/0010-406X(70)90049-6 4988625
- HarrisRCMarlinDJDunnettMSnowDHHultmanEMuscle buffering capacity and dipeptide content in the thoroughbred horse, greyhound dog and manComp Biochem Physiol A Physiol199097 249 251 1:STN:280:DyaK3M3ms1GktA%3D%3D https://doi.org/10.1016/0300-9629(90)90180-Z
- BoldyrevAAKoldobskiAKurellaEMaltsevaVStvolinskiSNatural histidine-containing dipeptide carnosine as a potent hydrophilic antioxidant with membrane stabilizing functionA biomedical aspect. Mol Chem Neuropathol199319 185 192 1:CAS:528:DyaK3sXltlSktrw%3D https://doi.org/10.1007/BF03160178 8363703
- BatrukovaMARubtsovAMHistidine-containing dipeptides as endogenous regulators of the activity of sarcoplasmic reticulum Ca-release channelsBiochim Biophys Acta19971324 142 150 1:CAS:528:DyaK2sXpt1yhtQ%3D%3D https://doi.org/10.1016/S0005-2736(96)00216-7 9059507
- HipkissARMichaelisJSyrrisPNon-enzymatic glycosylation of the dipeptide L-carnosine, a potential anti-protein-cross-linking agentFEBS Lett1995371 81 85 1:CAS:528:DyaK2MXotFCltrw%3D https://doi.org/10.1016/0014-5793(95)00849-5 7664889
- HipkissARCarnosine and protein carbonyl groups: a possible relationshipBiochemistry (Mosc)200065 771 778 1:CAS:528:DC%2BD3cXlvFOlsbw%3D
- DeraveWEveraertIBeeckmanSBaguetAMuscle carnosine metabolism and beta-alanine supplementation in relation to exercise and trainingSports Med201040 247 263 https://doi.org/10.2165/11530310-000000000-00000 20199122
- SmithECThe buffering of muscle in rigor; protein, phosphate and carnosineJ Physiol193892 336 343 1395289 1:CAS:528:DyaA1cXjsFyrtw%3D%3D https://doi.org/10.1113/jphysiol.1938.sp003605 16994977
- TanokuraMTasumiMMiyazawaT1 H nuclear magnetic resonance studies of histidine-containing di- and tripeptides, Estimation of the effects of charged groups on the pKa value of the imidazole ringBiopolymers197615 393 401 1:CAS:528:DyaE28Xht1agsL0%3D https://doi.org/10.1002/bip.1976.360150215 2351
- BaguetAKoppoKPottierADeraveWBeta-alanine supplementation reduces acidosis but not oxygen uptake response during high-intensity cycling exerciseEur J Appl Physiol2010108 495 503 1:CAS:528:DC%2BC3cXmsVGku7s%3D https://doi.org/10.1007/s00421-009-1225-0 19841932
- SuzukiYItoOMukaiNTakahashiHTakamatsuKHigh level of skeletal muscle carnosine contributes to the latter half of exercise performance during 30-s maximal cycle ergometer sprintingJpn J Physiol200252 199 205 1:CAS:528:DC%2BD38Xmt1ait7o%3D https://doi.org/10.2170/jjphysiol.52.199 12139778
- BaguetAEveraertIHespelPPetrovicMAchtenEDeraveWA new method for non-invasive estimation of human muscle fiber type compositionPLoS One20116 e21956 3131401 1:CAS:528:DC%2BC3MXpsFCgu70%3D https://doi.org/10.1371/journal.pone.0021956 21760934
- HaradaRTaguchiYUrashimaKSatoMOhmoriTMorimatsuFEnhancement of swimming endurance in mice by chicken breast extractJ Jpn Soc Nutr Food Sci (in Japanese)200255 73 78 1:CAS:528:DC%2BD38Xkt1CjsrY%3D https://doi.org/10.4327/jsnfs.55.73
- DunnettMHarrisRCInfluence of oral beta-alanine and L-histidine supplementation on the carnosine content of the gluteus mediusEquine Vet J Suppl199930 499 504 10659307
- SatoMSuzukiYMorimatsuFTakamatsuKEffect of carnosine concentration in muscle and improvement of exercise performances due to long-term intake of chicken breast extractJpn J Phys Fitness Sports Med (in Japanese)200352 255 264
- HarrisRCTallonMJDunnettMBoobisLCoakleyJKimHJFallowfieldJLHillCASaleCWiseJAThe absorption of orally supplied beta-alanine and its effect on muscle carnosine synthesis in human vastus lateralisAmino Acids200630 279 289 1:CAS:528:DC%2BD28Xktlaksrk%3D https://doi.org/10.1007/s00726-006-0299-9 16554972
- SaleCSaundersBHarrisRCEffect of beta-alanine supplementation on muscle carnosine concentrations and exercise performanceAmino Acids201039 321 333 1:CAS:528:DC%2BC3cXotVaisL4%3D https://doi.org/10.1007/s00726-009-0443-4 20091069
- TeufelMSaudekVLedigJPBernhardtABoularandSCarreauACairnsNJCarterCCowleyDJDuvergerDGanzhornAJGuenetCHeintzelmannBLaucherVSauvageCSmirnovaTSequence identification and characterization of human carnosinase and a closely related non-specific dipeptidaseJ Biol Chem2003278 6521 6531 1:CAS:528:DC%2BD3sXht1ejsLY%3D https://doi.org/10.1074/jbc.M209764200 12473676
- SauerhöferSYuanGBraunGSDeinzerMNeumaierMGretzNFloegeJKrizWvan der WoudeFMoellerMJL-Carnosine, a substrate of carnosinase-1, influences glucose metabolismDiabetes200756 2425 2432 https://doi.org/10.2337/db07-0177 17601992
- DrozakJVeiga-da-CunhaMVertommenDStroobantVSchaftingenEVMolecular identification of carnosine synthase as ATP-grasp domain-containing protein 1 (ATPGD1)J Biol Chem2010285 9346 9356 2843183 1:CAS:528:DC%2BC3cXjsFehsLw%3D https://doi.org/10.1074/jbc.M109.095505 20097752
- KalyankarGDMeisterAEnzymatic synthesis of carnosine and related beta-alanyl and gamma-aminobutyryl peptidesJ Biol Chem1959234 3210 3218 1:CAS:528:DyaF3cXivFaitw%3D%3D 14404206
- SteneshJJWinnickTCarnosine-anserine synthetase of muscle. 4. Partial purification of the enzyme and further studies of beta-alanyl peptide synthesisBiochem J196077 575 581 1205078 1:CAS:528:DyaF3MXjvVaitQ%3D%3D https://doi.org/10.1042/bj0770575 16748858
- SkaperSDDasSMarshallFDSome properties of a homocarnosine-carnosine synthetase isolated from rat brainJ Neurochem197321 1429 1445 1:CAS:528:DyaE2cXpt1Gnsw%3D%3D https://doi.org/10.1111/j.1471-4159.1973.tb06027.x 4358880
- HorinishiHGrilloMMargolisFLPurification and characterization of carnosine synthetase from mouse olfactory bulbsJ Neurochem197831 909 919 1:CAS:528:DyaE1MXhsVOntbs%3D https://doi.org/10.1111/j.1471-4159.1978.tb00127.x 100576
- KamalMAJiangHHuYKeepRFSmithDEInfluence of genetic knockout of Pept2 on the in vivo disposition of endogenous and exogenous carnosine in wild-type and Pept2 null miceAm J Physiol Regul Integr Comp Physiol2009296 986 991 https://doi.org/10.1152/ajpregu.90744.2008
- BaguetAEveraertINaeyerHDReyngoudtHStegenSBeeckmanSAchtenEVanheeLVolkaertAPetrovicMTaesYDeraveWEffects of sprint training combined with vegetarian or mixed diet on muscle carnosine content and buffering capacityEur J Appl Physiol2011111 2571 2580 1:CAS:528:DC%2BC3MXht1SitbzK https://doi.org/10.1007/s00421-011-1877-4 21373871
- MaemuraHGotoKYoshiokaTSatoMTakahataYMorimatsuFTakamatsuKEffects of Carnosine and Anserine Supplementation on Relatively High Intensity Endurance PerformanceInternational Journal of Sport and Health Science20064 86 94 https://doi.org/10.5432/ijshs.4.86
- DeraveWOzdemirMSHarrisRCPottierAReyngoudtHKoppoKWiseJAAchtenEbeta-Alanine supplementation augments muscle carnosine content and attenuates fatigue during repeated isokinetic contraction bouts in trained sprintersJ Appl Physiol2007103 1736 1743 1:CAS:528:DC%2BD2sXhtlyrtrzK https://doi.org/10.1152/japplphysiol.00397.2007 17690198
- HillCAHarrisRCKimHJHarrisBDSaleCBoobisLHKimCKWiseJAInfluence of beta-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacityAmino Acids200732 225 233 1:CAS:528:DC%2BD2sXhtFyhtbo%3D https://doi.org/10.1007/s00726-006-0364-4 16868650
- CaseyAGreenhaffPLDoes dietary creatine supplementation play a role in skeletal muscle metabolism and performance?Am J Clin Nutr200072 607S 617S 1:CAS:528:DC%2BD3cXlsFyhsb8%3D 10919967
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.