ABSTRACT
Soil frost formation, snow distribution, and winter/spring/summer terminal infiltration rates (TIRs) were quantified in Icelandic Andisols with contrasting vegetation cover types (grassland, spruce and birch woodland, lupine, and sparsely vegetated lava site). TIRs (mm h−1; determined with double-ring infiltrometers) were generally higher in unfrozen than in frozen soils (102–369 vs. 9–306, respectively in sandy soils; 28–94 vs. 3–72 in finer-textured soils) and differed between land cover types, being consistently highest in birch woodlands. TIR was an inverse function of soil frost depth. Lowest TIRs were associated with deep and dense soil frost, which formed in spruce woodland and grassland communities where snow depth was shallow. Results suggest conditions conducive to erosion by water are most likely to occur during winter warm spells and in spring in vegetation types where snow cover is low or ephemeral. Threefold increases in TIRs occurred one year after livestock grazing was discontinued, suggesting Andisols are hydrologically resilient where vegetation cover is relatively continuous and soil organic carbon content is high.
Introduction
When ecosystems become dysfunctional, hydrological processes are significantly altered, often resulting in high runoff and loss of soil and nutrients. Hence, criteria related to hydrologic function are often used to evaluate ecosystem stability and resistance to disturbance. The capacity of a site to capture and store precipitation, run-on, and snowmelt, and the stability of this capacity, are key determinants of ecohydrological function (CitationLudwig et al., 1997; CitationPellant et al., 2000; CitationLudwig et al., 2005). Infiltration is an important component of water capture potential.
Vegetation enhances infiltration by increasing surface roughness, by reducing raindrop impact, and by improving soil porosity via root channels, addition of organic matter, and providing habitat for soil biota (e.g., CitationBranson et al., 1981; CitationThurow, 1991). These influences vary with plant functional groups, with infiltration generally being highest in soils under trees and shrubs, lower in grasslands, and least in sparsely vegetated areas (CitationThurow et al., 1986). In high latitude ecosystems, vegetation additionally influences infiltration via its effects on radiant energy exchange and snow capture which, in turn, affect soil frost formation (e.g., CitationTyrtikov, 1976; CitationJones et al., 2001). Occurrence of soil frost and depth of freezing in relation to vegetative cover has been found to decrease in the rank order: bare field > grassland > evergreen conifer forest > deciduous forest (CitationPierce et al., 1958; CitationShanley and Chalmers, 1999).
Soil frost can impede or enhance infiltration. Concrete frost, which consists of many thin ice lenses and small crystals, may be essentially impermeable to water (CitationPost and Dreibelbis, 1942; CitationTrimble et al., 1958; CitationDunne and Black, 1971). Soil frost with loose crystals may be porous (CitationPost and Dreibelbis, 1942; CitationStorey, 1955); and may have infiltration rates greater than their unfrozen counterparts (CitationTrimble et al., 1958; CitationHaupt, 1967; CitationBlackburn and Wood, 1990) due to soil structural changes accompanying freezing (e.g., enhanced aggregation; CitationLuo et al., 2003). Concrete frost tends to develop in soils with no or delayed snow cover (CitationHaupt, 1967; CitationFahey and Lang, 1975) and in open land susceptible to deep frost penetration (CitationPierce et al., 1958; CitationStoeckeler and Weitzman, 1960); it is less common in forested areas, particularly deciduous forests (CitationPierce et al., 1958; CitationStoeckeler and Weitzman, 1960; CitationFahey and Lang, 1975). High soil water content promotes formation of dense soil frosts that retard infiltration (CitationKane and Stein, 1983). Consequently, rainfall or snowmelt during warm spells followed by freezing temperatures enhances concrete frost formation (CitationProulx and Stein, 1997; CitationShanley and Chalmers, 1999). An understanding of the ecohydrology of high latitude ecosystems thus requires an understanding of how vegetation and climate interact to affect the type of soil frost that may form.
Soils in the Andisol order are common in volcanically active regions of the world (CitationSoil Survey Staff, 1999). Although Andisols have high infiltration rates (CitationForsythe, 1975; CitationShoji et al., 1993), they are highly vulnerable to erosion by water (CitationNishimura et al., 1993; CitationPoulenard et al., 2001) and wind (CitationArnalds, 2000; CitationArnalds et al., 2001), due to the low-density soil aggregates, lack of particle cohesion, and high soil water retention (CitationWada et al., 1992; CitationKimble et al., 2000). High water retention also makes high latitude Andisols susceptible to frost heaving and needle ice cryoturbation. Natural or anthropogenic reduction of the plant biomass thermal barrier may thus intensify cryogenic processes that hasten formation of bare patches (CitationArcher and Stokes, 2000) while promoting the formation of impermeable soil frost types.
Andisols are the dominant soil order in Iceland (CitationArnalds, 2004) where extensive vegetation change, loss of plant cover, and soil erosion have occurred (CitationAradottir and Arnalds, 2001; CitationArnalds et al., 2001). Since settlement about 1100 years ago, birch woodland cover is estimated to have declined from 25% (CitationSigurdsson, 1977) to only 1% (CitationGudjónsson and Gíslason, 1998); and lands classified as barren desert have increased from 10–15% cover to >35% cover (CitationArnalds, 2000). Erosion by water in Iceland is significant and a significant fraction of this erosion occurs in conjunction with winter rains and spring snowmelt (CitationArnalds et al., 2001). An understanding of how vegetation and land cover change affects hydrological processes influencing erosion is central to the development of land management protocols aimed at curtailing land degradation and restoring dysfunctional ecosystems. In this study, we sought to quantify the relationship between vegetation cover, soil frost formation, and infiltration in Icelandic Andisols.
The maritime winter climate of Iceland is characterized by freeze-thaw cycles and thin and fluctuating snow cover (CitationEinarsson, 1984). We therefore expected that high frequencies of freeze-thaw events, coupled with the high water holding capacity of Andisols, would cause the formation of impermeable, ice-rich zones. We hypothesized that (a) infiltration rates in such zones would be reduced, hence increasing runoff and soil erosion potential following winter rain and snowmelt runoff, but that (b) the formation of such zones would vary as a function of the type and amount of vegetation cover. As Andisols have intrinsically high infiltration rates, we also hypothesized that (c) type of vegetation cover would not affect infiltration rates in unfrozen soil. To test these hypotheses, we quantified rates of water infiltration and soil frost dynamics in contrasting land cover types during one year.
Study Sites
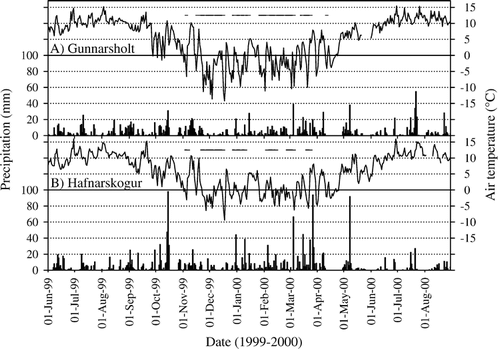
Studies were conducted at two sites in the lowlands of South and West Iceland. These sites represented contrasting land cover types spanning a range of vegetation degradation and regeneration states. The Gunnarsholt site (G-site; 63°52′N, 20°12′W; ca. 100 m a.s.l.) consisted of revegetated landscapes, whereas the Hafnarskogur site (H-site; 64°30′N, 21°55′W; ca. 30 m a.s.l.) represented degraded landscapes (). Mean January and July temperatures at these sites are −1 to −2 °C and 10 to 11 °C, respectively; mean annual precipitation is 1200 mm (G-site) and 1500 mm (H-site). Temperatures are typically less extreme and snow cover shallower at the H-site compared to the G-site, owing to its proximity to the sea (<1 km versus 25 km, respectively). Weather conditions at both sites during the study years 1999 to 2000 are shown in .
FIGURE 1 Location of the Gunnarsholt (G) and Hafnarskogur (H) study sites in Iceland.
FIGURE 2 Daily precipitation (bars) and air temperature (line), and days with 100% snow cover (horizontal lines) at (A) Gunnarsholt and (B) Hafnarskogur, June 1999 to August 2000 (CitationVeðráttan, 1999, Citation2000). Gunnarsholt, data from weather station Hella (11 km WSW of site); Hafnarskogur, temperature from Hafnarmelar (5 km SSW of site), precipitation and snow cover from Andakílsárvirkjun (12 km ENE of site).
The G-site, excluded from livestock grazing since the late 1920s, is comprised of small hills formed by post-glacial basaltic lava flows onto otherwise relatively level terrain. Soils are deep (>1 m) Vitricryands (O. Arnalds, unpublished; CitationStrachan et al., 1998) or Brown Andosols (Icelandic classification scheme; CitationArnalds, 2004) developed in eolian and tephra materials deposited on the lava field. Sand encroachment covered much of the area with dunes in the late 1800s (CitationSigurjónsson, 1958). Reclamation efforts in the 1900s stabilized these dunes and created a mosaic of plant communities, five of which were used in this study. These communities occur in two areas (referred to as western and eastern) located about 1 km apart. Birch (Betula pubescens Ehrh.) woodland, lupine (Lupinus nootkatensis Donn ex Sims) fields, and lava fields sparsely vegetated with Festuca grasses and ericaceous dwarf shrubs occur in the western area; and a Sitka spruce (Picea sitchensis (Bong.) Carrière) woodland and grassland dominated by Festuca and Agrostis grasses and Galium herbs occur in the eastern area. We refer to these different plant communities as G-birch, G-lupine, G-lava, G-spruce, and G-grassland, respectively. The birch woodland developed from seed planted in 1939 and 1945 (CitationMagnússon and Magnússon, 1989), and consists of birch (mean height 2.2 m) and willows (Salix phylicifolia L. and S. lanata L.) with a density of stems >30 cm height = 9774 stems ha−1. The first lupines were introduced at the site in 1988 (Sveinn Runólfsson, personal communication). Vegetation on the lava field is self-regenerated. The grassland community developed from Leymus arenarius (L.) Hochst. seeds sown around 1930. The Sitka spruce stand (15 × 100 m; 2525 trees ha−1; mean height 5–6 m) was planted in early 1960s (Sveinn Runólfsson, personal communication). Plant nomenclature follows CitationKristinsson (2005).
The H-site includes hummocky birch woodland (density of stems > 30 cm height = 5344 stems ha−1; mean height = 1.7 m) and Agrostis-Festuca-Deschampsia grassland considered to have had some woodland cover in earlier times. These will subsequently be referred to as H-birch and H-grassland. Soils are deep (>1m) Haplocryands (O. Arnalds, unpublished; CitationStrachan et al., 1998) or Histic Andosols (Icelandic classification scheme; CitationArnalds, 2004) formed in eolian and tephra materials deposited on an old (about 10,000 yr) sand/gravel shoreline. Sheep grazing was discontinued in the early 1990s, but horses graze the grassland, except for short periods in mid-summer. The H-grassland research plot was fenced in August 1999 to exclude livestock and prevent damage to instrumentation.
Methods
Infiltration Rates
Metal double-ring infiltrometers (CitationBouwer, 1986) were installed in each plant community (n = 6) in June/July 1999. The rings were 40 cm high, with 22.5 and 39.0 cm inner and outer diameters, respectively. Rings were driven 30 cm into the soil. Placement of rings was between hummocks, and midway between trees in woodlands. Because one of our objectives was to quantify infiltration rates on frozen soils, and because installation of rings in frozen soils would have been impossible without causing unacceptable levels of disturbance, rings were left in place for the duration of the study. A repeated-measures design was thus used (e.g., Citationvan der Kamp et al., 2003). Initial readings were made between 17 July and 5 August 1999. Due to time constraints, no measurements were made on G-grassland. Winter and spring measurements were made during three periods in 2000: 5–11 January, 15–21 April, and 14–22 May. The G-birch, G-lupine, and G-lava communities were inaccessible in January due to snow.
A water head of 4 cm was maintained within rings during infiltration trials, and the water required to maintain the head in the inner ring was recorded at five minute intervals for one hour. Terminal infiltration rates (TIRs; mm h−1) were calculated as the mean of the last three measurements. The temperature of water used approximated that of rainfall during the measurement periods: 0°C in January, 2–4°C in April, and 6–8°C in May and July/August.
Snow, Ice, and Soil Frost
Snow cover (%) and depth (cm) were measured within infiltration rings and along transects in each plant community before infiltration trials in January, but only within rings in April, as snow measured in rings and transects were comparable. Ice cover within rings was estimated visually.
Soil frost depth (cm) and type were quantified during infiltration trials in January, April, and May. Frost depth was measured in small pits near each infiltrometer by visually noting the occurrence of ice crystals. The size and number of ice crystals observed with a 10× hand lens was used to classify frost into one of three categories using criteria in CitationPost and Dreibelbis (1942) and CitationStoeckeler and Weitzman (1960): concrete frost—a high density of small crystals; porous concrete frost—resembled concrete frost, but with lower crystal density; and porous frost—fewer and larger ice crystals than the concrete frost types. Soil frost measurements were not possible in G-birch, G-lupine, and G-lava in January because of snow. Shallow standing water in depressions in H-grassland in January limited soil frost measurements (n = 3).
Ground Cover and Soil
Ground cover and plant composition of inner infiltration rings was recorded in July/August 2000 using a point-frame (5 cm pin interval; 15–17 pins per ring). For each pin drop, all contacts with vegetation, litter, stones, and soil were recorded. Aboveground biomass (g m−2) of the ground layer was quantified by clipping vegetation and litter within the inner rings, sorting by life form, and oven-drying at 60 °C. Vascular plant species diversity was computed as richness (number of species) and as an abundance-weighted richness (Simpson's Diversity Index; CitationHuston, 1994) in 30 plots per community using a point-frame (n = 5 pins per plot).
Soil properties within each inner infiltration ring (to 30 cm depth; by horizon) were determined on samples obtained at the end of the study (August/September 2000). Bulk density (BD) was determined by the core method (CitationBlake and Hartge, 1986). Organic carbon (OC) was quantified by dry combustion (Leco CR-12 carbon analyzer; CitationNelson and Sommers, 1982). Sand content was determined on field-moist samples. Field texture and water content of field-moist surface soil samples at –0.033 MPa (pressure chambers; CitationKlute, 1986) were determined on samples from the soil pedons described in each plant community.
The sequence and designation of genetic horizons in the soils varied greatly between sites (). Therefore, BDs in the different communities were not compared on the basis of horizon designations but on their three upper horizons: surface horizons were denoted horizon 1; the subtending horizon was denoted horizon 2; and the deepest horizons (down to ca. 30 cm depth) were denoted as horizon 3 (and 4 at H-grassland). The properties of the genetic horizons represented in these groups are summarized in .
TABLE 1 Topographic setting and selected soil and vegetation characteristics in the contrasting land cover types at the Gunnarsholt (G) and Hafnarskogur (H) study sites in South and West Iceland. See for bulk density data.
TABLE 2 Mean (± 1 SE) bulk density (g cm−3) of soil horizons from the surface to 30 cm depth in plant communities at Gunnarsholt (G) and Hafnarskogur (H). Means are n of 6 samples except: horizons 1 and 2 in G-lupine (n = 5); and horizon 3 in G-lupine (n = 4). See for horizon designations for each plant community.
Experimental Design and Data Analysis
Logistical constraints forced us to choose between surveying numerous plant communities or studying one or two replicated communities. We opted for the former, as information on the infiltration/frost dynamics of Andisols with different vegetation cover is lacking. Infiltration rings were treated as the unit of replication (n = 6). Extrapolation of our results are thus predicated on the assumption that the stands we sampled are broadly representative of the community type, an assumption that should be evaluated in future research. Statistical analyses were performed with SAS v.8.01 (CitationSAS, 2000). Ground cover and depth of soil frost were analyzed using analysis of variance (ANOVA), where communities (G-spruce, H-birch, etc.) were treated as main effects. Between-community comparisons of snow depth measured along transects were ascertained with a Kruskal-Wallis test. Spearman rank correlation coefficients (r s) were determined for the relationship between January soil frost depth and snow depth within the infiltrometers.
TIR data were analyzed separately for communities in the revegetated and degraded sites. The effect of soil frost depth on TIR was tested with analysis of covariance (ANCOVA) for the H-site April data. Simple linear regression was used to describe the relationship between soil frost depth and TIR at the G-site in April, excluding rings with 100% ice cover. At the H-site, plant community differences in TIR were analyzed separately with ANOVA on a given date. This was also done at the G-site for the January and April TIRs, but plant community effects on TIR in unfrozen soils were assessed with univariate, repeated measures ANOVA, with an error term consisting of the nested variation of the infiltration rings within the plant communities (CitationLittell et al., 1998). Correlation coefficients (r s) were determined for the relationship between vegetation and soil properties and the July/August 2000 TIRs. When overall tests were significant (p < 0.05), sequential Bonferroni multiple comparison tests (CitationRice, 1989) were used to make pair-wise comparisons of means. Data were transformed prior to analysis if assumptions were not met.
Results
Ground Cover and Soils
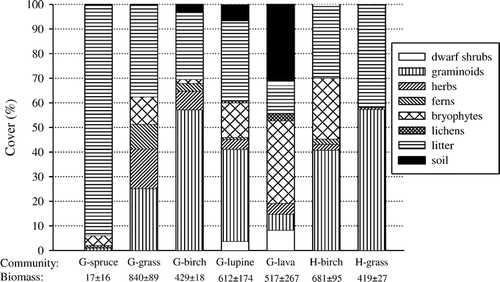
Soils, vegetation, and topography are summarized in . OC content was highest in surface (19.6–25.0%) and subsurface (8.0–20.2%) horizons at the H-site and lowest in G-lupine and G-lava soils (1.6–2.4%). At the H-site, soil field texture was finer (loam, clay loam) than at the G-site (sand, sandy loam, loam, silt loam). Water-holding capacity of the H-birch and H-grassland surface soils was high (162% and 167% at −0.033 MPa, respectively) as was that of the G-spruce surface soil (125%); water-holding capacity in other communities was ≤81%. Surface horizon BD ranged from 0.24 to 0.76 g cm−3 and was lowest in the G-spruce, H-birch, and H-grassland, and highest in G-lupine and G-lava ().
Vascular plant species richness of ground cover was greatest in the G- and H-birch woodlands and G-lava communities and least in G-spruce woodland (). Bare soil cover was high in the G-lava (31%) community, but <10% in all others (). Litter dominated the G-spruce surface. Vascular plant ground cover differed between plant communities (p < 0.001): cover was lowest in G-spruce, highest and comparable in G-birch, G-grassland, G-lupine, H-birch, and H-grassland, and intermediate in G-lava. Total aboveground biomass ranged from 17 g m−2 in G-spruce to 840 g m−2 in G-grassland ().
FIGURE 3 Relative ground cover (%) and total aboveground biomass (mean ± 1 SE; g m−2) within inner infiltration rings in plant communities at Gunnarsholt (G) and Hafnarskogur (H). Data are means of 6 rings per plant community (n = 5 for G-lupine) in July/August 2000. The biomass of litter (duff layer) in the spruce woodland not included.
Snow, Ice and Soil Frost
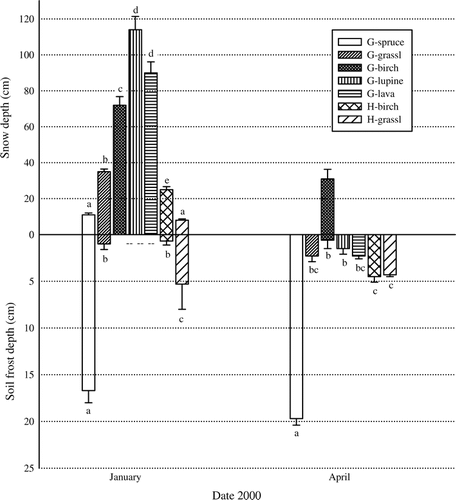
The winter 1999–2000 was characterized by more persistent and greater snow cover than in average years (CitationVeðráttan, 1999, Citation2000). Data from the weather station nearest each study site provide a crude index of snow cover persistence and continuity (). Transect measurements on our sites documented continuous snow cover in early January in all plant communities, except H-grassland where cover was 86%. However, mean snow depth differed between communities (p < 0.001), being deepest in the G-birch, G-lupine, and G-lava communities (72–114 cm), and shallowest (≤11 cm) in G-spruce and H-grassland (). By April, snow had disappeared in all but the G-birch community (), where it persisted until the end of April. The snow cover data from weather station Hella (), and unpublished soil temperature data in the G-birch, G-spruce, and G-lava communities suggest that snow in the other G-communities disappeared by the end of March.
FIGURE 4 Upper panel: mean snow depth (± 1 SE; cm) measured along transects (n = 13–49) in January, and within inner infiltration rings in April. Lower panel: mean soil frost depth (± 1 SE; cm) near infiltration rings in plant communities at Gunnarsholt (G) and Hafnarskogur (H) in January and April 2000. Means followed by different letters within each date were significantly different (p < 0.05). Means of soil frost are n of 6 samples except: in January n = 3 for H-grassland; in April n = 5 for G-birch. Concrete frost characterized G-spruce soil; porous concrete frost characterized H-grassland soils; and porous frost characterized soils in all other communities. Deep snow prevented soil frost data collection in January for G-birch, G-lupine and G-lava.
G-grassland and H-birch infiltration rings were ice-free in January 2000. Five of six H-grassland rings had ice cover (100% cover in four) as did three of six G-spruce rings (but none had 100% cover). Ice was only observed in two G-spruce rings in April (100% cover in each) and only one G-birch ring (25% cover). The observed occurrence of ice cover in rings was qualitatively representative of the patchiness of ice distribution in the broader communities (personal observation).
Soil frost depth in January ranged from a mean of 0.7 cm in H-birch to 16.7 cm in G-spruce. In April the mean frost depth ranged from 0.6 cm in G-birch to 19.7 cm in G-spruce (). G-spruce was the only community where soil frost was observed in May (data not shown). By this time, surface soils had thawed but a frost lens beginning at 8.4 ± 2.7 cm depth and extending to 15.0 ± 2.5 cm was still evident. Soil frost depth differed between communities both in January (p < 0.001) and April (p < 0.001) (). It is noteworthy that at the H-site, although soil frost in January had penetrated to greater depths in the grassland compared to the deciduous birch woodland (5.3 cm and 0.7 cm, respectively), depths were comparable in April (4.3 and 4.5 cm) owing to increases in woodland frost depths. Increased frost depths from January to April were also observed in the evergreen G-spruce woodland, although soil frost there was already deep in January. In contrast, soil frost depths in the grassland communities were comparable in January and April. January soil frost depth and snow depth within infiltrometers were negatively correlated both at the G-site (r s = −0.90, p < 0.001; includes the spruce woodland and grassland) and the H-site (r s = −0.51, p < 0.16).
All three structural types of soil frost (concrete, porous concrete, and porous) were observed during January and April. Concrete frost characterized all soil samples in G-spruce. Porous concrete frost characterized all samples in H-grassland, whereas porous frost characterized soils in G- and H-birch, G-grassland, G-lupine, and G-lava. The frost types did not appear to change over time within a community. Qualitative observations indicated ice crystals in G-spruce and H-grassland organic soil horizons were larger than those occurring in the mineral horizons below. Needle ice was observed among the porous frost of G-lava soils (three of six April samples).
Infiltration Rates
Terminal infiltration rates (TIRs) were typically achieved within 10 to 20 minutes. Winter TIRs ranged from 3 to 306 mm h−1. Summer TIRs ranged from 28 to 369 mm h−1. As a rule, rates were lowest in the H-grassland community and highest in the G-birch woodland ().
FIGURE 5 Mean (± 1 SE) terminal infiltration rates (mm h−1) in plant communities at Gunnarsholt (G) and Hafnarskogur (H) on five dates. Means are from six double-ring infiltrometers except: G-birch and G-lupine (n = 5); G-lava in July/August 1999 (n = 5); and G-birch in April 2000 (n = 4).
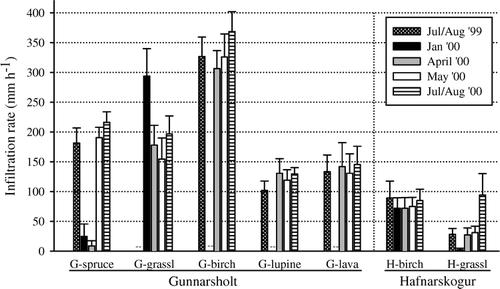
TIRs within the plant communities varied with time, the greatest differences occurring between the summer and winter dates (). TIRs in H- and G-birch woodlands, and the G-lupine and G-lava field communities varied between dates by a factor of 1.1 to 1.3. G-grassland TIRs varied by a factor of 2 (January vs. May 2000), and G-spruce woodland TIRs varied by a factor of 25 (April vs. July/August 2000). The greatest date difference was observed in H-grassland where the January and July/August 2000 TIRs varied by a factor of 37, and summer measurements varied by a factor of 3. Peak TIR occurred in summer for all communities except G-grassland, where peak TIR occurred in January.
Winter Rates
Infiltration was totally inhibited in rings where surface ice cover was 100%. There was no indication of soil thawing during the 60-minute trials. In January, at the H-site, birch woodland had greater TIR than grassland (p < 0.001); however, TIRs in G-grassland were much greater than those in H-grassland and far exceeded those of G-spruce woodland (p < 0.001) (). At the H-site in April, TIR was most strongly correlated with BD of the surface soil horizon (r s = −0.81, p < 0.01). Correlations between TIR and soil frost depth were not significant (r s = −0.30, p = 0.35) and accounting for soil frost depth did not improve prediction of TIR (ANCOVA; p = 0.24, R 2 = 0.40). TIRs in the two H-site communities differed at p = 0.067 with no interaction between plant community type and soil frost depth (p = 0.85). At the G-site, April TIRs in the five plant communities were most strongly correlated to soil frost depth (r s = −0.75, p < 0.001). Exclusion of the spruce woodland, which exerted a strong influence on the data spread by virtue of its substantially greater soil frost depth (), reduced the Spearman r to −0.59 (p < 0.01). Using all G-site data points, the relationship between soil frost depth and TIR was best described by a non-linear regression function (p < 0.001, R 2 = 0.61; ), whereas, when excluding G-spruce, a linear relationship explained most variability in the data (p < 0.001, R 2 = 0.45; ). TIRs in the five communities differed (p < 0.001) in April in the rank order G-spruce < G-lupine ≈ G-lava ≈ G-grassland < G-birch woodland.
FIGURE 6 Relationship between terminal infiltration rate (TIR; mm h−1) and soil frost depth (cm) in April 2000, in plant communities at Gunnarsholt. Non-linear fit (all data [TIR = 262.11 − 204.17 × log (frost depth + 1)]) and linear fit (excluding G-spruce [TIR = 266.63 − 48.69 × (frost depth)]) are presented.
![FIGURE 6 Relationship between terminal infiltration rate (TIR; mm h−1) and soil frost depth (cm) in April 2000, in plant communities at Gunnarsholt. Non-linear fit (all data [TIR = 262.11 − 204.17 × log (frost depth + 1)]) and linear fit (excluding G-spruce [TIR = 266.63 − 48.69 × (frost depth)]) are presented.](/cms/asset/ca1ad33a-f454-41d3-b1df-6e5cb92e2c21/uaar_a_11957328_f0008.gif)
Summer Rates
Summer TIRs differed (p < 0.001) between G-site plant communities in the rank order spruce woodland ≈ lupine ≈ lava field < birch woodland. At the H-site, TIRs in the birch woodland were higher than those in the grassland in July/August 1999 (p = 0.04) and May 2000 (p = 0.04), but were comparable in July/August 2000 (p = 0.82) (). Variables strongly related to July/August 2000 TIR in the G-site communities were surface soil BD (r s = −0.59, p < 0.01) and organic cover (r s = 0.50, p < 0.01). At the H-site, BD of the surface soil horizon was the only variable significantly correlated with TIR in July/August 2000 (r s = −0.85, p < 0.001).
Discussion
The results indicate that the formation of ice-rich zones in Andisols reduces infiltration rates. The formation of these ice-rich zones appeared to vary with the type of soils and vegetation cover, but not with the amount of plant ground cover, as ice-rich zones with low TIRs occurred in both sparsely (G-spruce) and well-vegetated (H-grassland) Andisols. Contrary to expectations, type of vegetation cover also had a significant influence on TIRs of unfrozen soils.
The TIR data presented here were generated using ring infiltrometers left in place for a full annual cycle. Infiltration rates measured with ring infiltrometers do not represent rates occurring under natural rainfall (CitationBranson et al., 1981); therefore, the rates we present are an assessment of the relative differences between plant communities and soils. There are some considerations when using this technique in high latitude environments. Metal rings with their high thermal conductivity may influence soil frost formation. However, similarities in frost type and depth in soils adjacent to and away from rings suggest this was not a problem in our study (data not shown). To avoid ring-induced ice formation, we recommend drilling small holes in the rings just above the ground surface, such that water can freely exit downslope. During measurements, the holes are plugged. Insertion of rings to 30 cm depth was sufficient to prevent ejection by frost heaving.
Infiltration Rates in Summer
In a review of infiltration rates in soils derived from volcanic ash in Central America, CitationForsythe (1975) reported 2-hour rates of 70–210 mm h−1. Our results for unfrozen Icelandic Andisols extend this range substantially in both directions (28–369 mm h−1). TIRs in Icelandic Andisols were generally higher in the sandier soils at the G-site (102–369 mm h−1), compared to in the finer textured soils at the H-site (28–94 mm h−1) (, ). Land cover influences superimposed against this backdrop of texture effects contributed to the broad range of observed TIRs. For example, TIR in G-birch woodland was about 4× that in the H-birch woodland and TIR in G-grassland was 2–6× that of H-grassland.
Plant life-form effects on TIR were substantial () and these have significant implications for management and restoration of Iceland's young, erosion-prone soils. TIRs were typically highest in woodlands dominated by native birch. The high TIRs in birch woodlands may be attributed to the dominance of shrub growth forms, which typically augment infiltration by enhancing soil structure (CitationThurow, 1991) and macropore formation (CitationBeven and Germann, 1982). Lower TIRs in the G-spruce compared to G-birch woodland may reflect the sparser plant ground cover in the spruce woodland (). Birch stands are also known to support greater earthworm biomass than spruce stands in Iceland (CitationGudleifsson, 2007). A greater abundance of other soil biota (CitationSaetre, 1998) which alter soil structure to promote infiltration may also foster higher rates of infiltration in birch communities.
Variability in summer TIRs within plant communities was relatively small, with one exception. In the H-grassland, TIR was three times higher in July/August 2000 compared to July/August 1999 and May 2000 (). Horses were grazing the site in July/August 1999, whereas in July/August 2000 the area had been excluded from grazing for one year and cover and biomass of vegetation were noticeably higher (qualitative observations). We hypothesize that the grazing exclusion led to increased plant root production which acted in combination with winter freeze-thaw events to counteract soil compaction caused by hoof action. The threefold greater TIR, observed one year after grazing was discontinued, suggests that Andisols are hydrologically resilient where vegetation cover is continuous and soil OC content high. Low TIR in H-grassland compared to H-birch communities in summer 1999 may thus reflect grazing rather than plant life-form effects. In the absence of grazing, H-grassland sites may have TIRs comparable to those of birch woodlands in unfrozen soils.
Soil Frost Dynamics and Infiltration Rates in Winter
TIRs in frozen versus unfrozen soils were generally comparable within the G- and H-birch woodlands, G-lupine, and G-lava field communities (). These sites all had continuous, relatively deep (≥25 cm) snow cover in January (). By contrast, TIRs in frozen and unfrozen soils associated with the G-spruce woodland and G- and H-grasslands varied dramatically. Moreover, the direction of change in TIR varied from summer to winter, with summer rates being highest in G-spruce woodland and the H-grassland, and winter (January) TIR being highest in the G-grassland. These summer vs. winter differences, or the lack thereof, appear to be related to the distinctive soil frost conditions occurring in each plant community.
Deep soil frost was associated with shallow snow depth (), as expected (e.g. CitationTyrtikov, 1976; CitationHardy et al., 2001). Shallow snow cover (≤11 cm) was observed in the open H-grassland community where snow was directly exposed to wind and solar radiation, but also in the G-spruce woodland where the dense nature of the spruce canopy seemed to cause high interception loss allowing little snow to reach the forest floor (CitationShanley and Chalmers, 1999; CitationJones et al., 2001). Infiltration-retarding concrete and porous concrete soil frost formed in these communities, whereas soils in the other communities developed porous frost. H-grassland and G-spruce had very high surface soil water retention (>100%; ); but despite similar water retention in the H-birch woodland, infiltration-retarding soil frost did not form there. Deeper snow cover in the H-birch woodland and/or lower soil water content at the time of freezing might be the explanation, but the greater snow cover can likely be attributed to woodland vegetation structure (CitationTyrtikov, 1976; CitationJones et al., 2001). Topography (slope inclination; ) appears to account for the difference in snow cover in the western vs. eastern portions of the G-site. Our observations reveal the importance of studying the influence of soil water content and snow cover at the time of freezing on the type and depth of frost formation in Andisols.
Deep concrete soil frost greatly reduced winter TIRs in the spruce woodland. Concrete frost has been found to hinder infiltration completely at frost depths from ca. 8 to 10 cm, measured at small scales (CitationPost and Dreibelbis, 1942; CitationMegahan and Satterlund, 1962), and to reduce infiltration significantly at spatial scales ranging from points to small catchments (e.g. CitationDunne and Black, 1971; CitationKane and Stein, 1983; CitationBlackburn and Wood, 1990; CitationShanley and Chalmers, 1999). In the spruce woodland, water infiltrated in only two (January) and one (April) of the six infiltrometers. Although the deep matrix of concrete soil frost was largely impermeable, root channels and other macropores that remain free of ice (CitationTrimble et al., 1958; CitationMegahan and Satterlund, 1962; CitationDunne and Black, 1971) may have allowed modest infiltration to occur in those rings. The negative exponential relationship between soil frost depth and TIR in the five G-site plant communities in April () supports the existence of such water conducting soil macropores. The logarithmic curve is consistent with what has been described for temperate forests (CitationRybakova, 1990), where water normally infiltrates through coarse root channels and voids created by soil organisms and freeze-thaw activity.
Porous concrete soil frost and patchy ice cover, common between hummocks, characterized the H-grassland in January when TIR was reduced compared to other times of the year. Even rings with partial or no ice cover had TIRs near zero, which is consistent with field observations of standing water at this site. However, by April, TIR had returned to pre-winter conditions () and was not influenced by soil frost, despite the fact that soil frost conditions were comparable to those in January (). These seemingly contradictory results may be an artifact of biased sampling of the soil in January, as frost depth could only be determined where water had not ponded. The limited number of measurements made at this time (n = 3) may therefore have underestimated average frost depth. Reports on the influence of porous concrete frost on infiltration are few and inconsistent. CitationStoeckeler and Weitzman (1960) measured reduced rates in soils characterized by porous concrete frost compared to unfrozen soil, whereas CitationHaupt (1967) measured reductions in infiltration on burned or sparsely vegetated plots, but not on plots where plant and litter cover was high. Our results imply that porous concrete frost can reduce infiltration in well-vegetated Andisols (H-grassland), but that depth of frost penetration may govern the influence on TIRs.
Plant communities characterized by soils with porous frost had comparable TIRs in frozen versus unfrozen soils ( and ) with one exception. G-grassland TIRs in January were elevated relative to those in summer, which may be attributable to increases in the number and/or size of pore spaces associated with frost action. Despite similar soil frost conditions in January and April, G-grassland April TIRs were lower than those recorded in January. This may be due to increased soil ice content following freeze-thaw cycles during snowmelt periods in January and March (), as snowmelt infiltration followed by soil freezing would increase soil ice content (CitationJohnsson and Lundin, 1991).
Conclusions
The high summer TIRs observed in this study imply that runoff is likely minimal on vegetated, level, unfrozen Andisols, especially since most precipitation comes in relatively small pulses. During winter months, infiltration-retarding soil frost occurred in herbaceous land cover type where exposure to wind and solar radiation limit snow accumulation. Deep, infiltration-retarding, concrete-type soil frost also formed in spruce woodland, a relatively new Icelandic ecosystem type (CitationPétursson, 1999). Concrete soil frost types were not observed in birch woodlands, and their high snow capture potential make them less prone to such frost formation. Even so, the extensive snow cover in winter 1999–2000 likely resulted in less extensive soil frost than might be expected in average years. Results from winters characterized by frequent thaw periods, and more shallow and ephemeral snow cover may thus prove different. Nonetheless, in this study the soils of birch woodlands were characterized by porous frost and exhibited high TIRs whether frozen or unfrozen. We thus expect that hydrological processes and soil frost dynamics may have changed significantly with the extensive loss of the birch woodlands in Iceland, leading to conditions that would be more conducive to water erosion during winter warm spells and periods of rainfall and snow melt in winter and spring. Open, sparsely vegetated areas have increased substantially in areal extent, because of land degradation. The tendency of such sites to develop deep, infiltration-retarding soil frost in winter suggests that these landscapes would be a likely source of runoff much of the year. Frequent river flooding during winter thaw events (CitationRist, 1990) supports this hypothesis. We also saw evidence for reductions in TIRs associated with reduced vegetation cover caused by livestock grazing, a widespread, traditional land use of the Icelandic rangelands. Effects of sheep grazing in high altitude Andisolic grasslands has been shown to create conditions for eolian erosion, runoff, and gully erosion (CitationPodwojewski et al., 2002). Extensive harvest of the native birch woodlands in concert with continuous livestock grazing by settlers in Iceland (CitationAradottir and Arnalds, 2001) may therefore have combined to effect local changes in hydrologic function that promote soil erosion.
Acknowledgments
This research was supported in part by the Icelandic Research Council, the Icelandic Soil Conservation Service, the Agricultural University of Iceland, and the Icelandic Agricultural Productivity Fund. Support from the Department of Rangeland Ecology and Management at Texas A&M University included a Springfield Research Award and a Graduate Student Support Fund award. We thank the Icelandic Soil Conservation Service and the landowners of Hafnarskogur for allowing us to conduct this study on their land. Special thanks to Fanney Gisladottir for making the map of the study sites, and to the staff at the Agricultural University of Iceland and the Soil Conservation Service who provided advice and unconditional logistical support through all aspects of the study.
Related Research Data
References Cited
- Aradottir, A. L. and O. Arnalds . 2001. Ecosystem degradation and restoration of birch woodlands in Iceland. In Wielgolaski, F. E. , editor. Nordic Mountain Birch Ecosystems Paris Parthenon Publishing Group. 293–306.
- Archer, S. and C. Stokes . 2000. Stress, disturbance and change in rangeland ecosystems. In Arnalds, O. and S. Archer , editors. Rangeland Desertification Dordrecht Kluwer Academic Publishers. 17–38.
- Arnalds, O. 2000. The Icelandic ‘rofabard’ soil erosion features. Earth Surface Processes and Landforms 25:17–28.
- Arnalds, O. 2004. Volcanic soils of Iceland. Catena 56:3–20.
- Arnalds, O. , C. T. Hallmark , and L. P. Wilding . 1995. Andisols from four different regions of Iceland. Soil Science Society of America Journal 59:161–169.
- Arnalds, O. , E. F. Thórarinsdóttir , S. Metúsalemsson , Á Jónsson , E. Grétarsson , and A. Árnason . 2001. Soil Erosion in Iceland Reykjavík Agricultural Research Institute and Soil Conservation Service. 121 pp.
- Beven, K. and P. Germann . 1982. Macropores and water flow in soils. Water Resources Research 18:1311–1325.
- Blackburn, W. H. and M. K. Wood . 1990. Influence of soil frost on infiltration of shrub coppice dune and dune interspace soils in southeastern Nevada. Great Basin Naturalist 50:41–46.
- Blake, G. R. and K. H. Hartge . 1986. Core method. In Klute, A. , editor. Methods of Soil Analysis. Part 1 Madison, Wisconsin American Society of Agronomy and Soil Science Society of America. 364–367.
- Bouwer, H. 1986. Intake rate: cylinder infiltrometer. In Klute, A. , editor. Methods of Soil Analysis. Part I Madison, Wisconsin American Society of Agronomy and Soil Science Society of America. 825–844.
- Branson, F. A. , G. F. Gifford , K. G. Renard , and R. F. Hadley . 1981. Rangeland Hydrology. 2nd edition Dubuque, Iowa Society for Range Management. 340 pp.
- Dunne, T. and R. D. Black . 1971. Runoff processes during snowmelt. Water Resources Research 7:1160–1172.
- Einarsson, MÁ 1984. Climate of Iceland. In Loon, H. V. , editor. World Survey of Climatology Amsterdam Elsevier. 673–697.
- Fahey, T. J. and G. E. Lang . 1975. Concrete frost along an elevational gradient in New Hampshire. Canadian Journal of Forest Research 5:700–705.
- Forsythe, W. M. 1975. Soil-water relations in soils derived from volcanic ash of Central America. In Bornemisza, E. and A. Alvarado , editors. Soil Management in Tropical America Raleigh, NC Soil Science Department, North Carolina State University. Proceedings of a Seminar held at CIAT, Cali, Colombia, February 10–14, 1974. 155–167.
- Gudjónsson, G. and E. Gíslason . 1998. A vegetation map of Iceland: general overview. Reykjavík Icelandic Institute of Natural History.
- Gudleifsson, B. E. 2007. ICEWOODS: Earthworms in Icelandic forest soils. In Halldórsson, G. , E. S. Oddsdóttir , and Ó Eggertsson , editors. Effects of afforestation on ecosystem, landscape and rural development Proceedings of the AFFORNORD conference, Reykholt, Iceland, June 18–22, 2005. 127–131.
- Hardy, J. P. , P. M. Groffman , R. D. Fitzhugh , K. S. Henry , A. T. Welman , J. D. Demers , T. J. Fahey , C. T. Driscoll , G. L. Tierney , and S. Nolan . 2001. Snow depth manipulation and its influence on soil frost and water dynamics in a northern hardwood forest. Biogeochemistry 56:151–174.
- Haupt, H. F. 1967. Infiltration, overland flow, and soil movement on frozen and snow-covered plots. Water Resources Research 3:145–161.
- Huston, M. A. 1994. Biological diversity. The coexistence of species on changing landscapes Cambridge Cambridge University Press. 681 pp.
- Johnsson, H. and L-C. Lundin . 1991. Surface runoff and soil water percolation as affected by snow and soil frost. Journal of Hydrology 122:141–159.
- Jones, H. G. , J. W. Pomeroy , D. A. Walker , and R. W. Hoham . 2001. Snow Ecology. An Interdisciplinary Examination of Snow-Covered Ecosystems Cambridge, U.K Cambridge University Press. 378 pp.
- Kane, D. L. and J. Stein . 1983. Water movement into seasonally frozen soils. Water Resources Research 19:1547–1557.
- Kimble, J. M. , C. L. Ping , M. E. Sumner , and L. P. Wilding . 2000. Andisols. In Sumner, M. E. and L. P. Wilding , editors. Handbook of Soil Science Boca Raton CRC Press. E209–E224.
- Klute, A. 1986. Water retention: laboratory methods. In Klute, A. , editor. Methods of Soil Analysis. Part 1 Madison, Wisconsin American Society of Agronomy and Soil Science Society of America. 635–662.
- Kristinsson, H. 2005. Checklist of wild flowering plants and ferns in Iceland (http://www.floraislands.is/plantlist.htm).
- Littell, R. C. , P. R. Henry , and C. B. Ammerman . 1998. Statistical analysis of repeated measures of data using SAS procedures. Journal of Animal Science 76:1216–1231.
- Ludwig, J. A. , D. J. Tongway , D. O. Freudenberger , J. C. Noble , and K. C. Hodgkinsons . 1997. Landscape Ecology, Function and Management: Principles from Australia's Rangelands Melbourne CSIRO Publishing. 158 pp.
- Ludwig, J. A. , B. P. Wilcox , D. D. Breshears , D. J. Tongway , and A. C. Imeson . 2005. Vegetation patches and runoff-erosion as interacting ecohydrological processes in semiarid landscapes. Ecology 86:288–297.
- Luo, L. F. , A. Robock , K. Y. Vinnikov , C. A. Schlosser , A. G. Slater , A. Boone , H. Braden , P. Cox , P. de Rosnay , R. E. Dickinson , Y. J. Dai , Q. Y. Duan , P. Etchevers , A. Henderson-Sellers , N. Gedney , Y. M. Gusev , F. Habets , J. W. Kim , E. Kowalczyk , K. Mitchell , O. N. Nasonova , J. Noilhan , A. J. Pitman , J. Schaake , A. B. Shmakin , T. G. Smirnova , P. Wetzel , Y. K. Xue , Z. L. Yang , and Q. C. Zeng . 2003. Effects of frozen soil on soil temperature, spring infiltration, and runoff: Results from the PILPS 2(d) experiment at Valdai, Russia. Journal of Hydrometeorology 4:334–351.
- Magnússon, S. H. and B. Magnússon . 1989. Factors in the ecology of birch and its use in restoration. In Arnalds, A. and A. G. Thórhallsdóttir , editors. Icelandic Yearbook of Soil Conservation, II Reykjavík Soil Conservation Service. 97–108. (in Icelandic).
- Megahan, W. F. and D. R. Satterlund . 1962. Winter infiltration studies on abandoned and reforested fields in central New York. Proceedings of the 19th Eastern Snow Conference 121–132.
- Nelson, D. W. and L. E. Sommers . 1982. In Page, A. L. , editor. Total carbon, organic carbon, and organic matter. Methods of Soil Analysis. Part 2 Madison, Wisconsin American Society of Agronomy and Soil Science Society of America. 539–579.
- Nishimura, T. , M. Nakano , and T. Miyazaki . 1993. Properties of surface crust of an Andisol and their effects on soil-hydrological processes. In Poesen, J. W. A. and M. A. Nearing , editors. Soil Surface Sealing and Crusting Selected papers of the International Symposium on Soil Crusting: chemical and physical processes, May 30–June 1, 1991,. Athens, Georgia, USA Destedt: Catena Verlag. 17–28.
- Pellant, M. , P. Shaver , D. A. Pyke , and J. E. Herrick . 2000. Interpreting Indicators of Rangeland Health. Version 3 Denver, Colorado U.S. Department of the Interior, Bureau of Land Management, National Science and Technology Center, Information and Communications Group.
- Pétursson, J. G. 1999. The first 100 years of tree planting in Iceland. Skógræktarritið 1999 2:49–53. (in Icelandic; English summary).
- Pierce, R. S. , H. W. Lull , and H. C. Storey . 1958. Influence of land use and forest condition on soil freezing and snow depth. Forest Science 4:246–263.
- Podwojewski, P. , J. Poulenard , T. Zambrana , and R. Hofstede . 2002. Overgrazing effects on vegetation cover and properties of volcanic ash soil in the páramo of Llangahua and La Esperanza (Tungurahua, Ecuador). Soil Use and Management 18:45–55.
- Post, F. A. and F. R. Dreibelbis . 1942. Some influences of frost penetration and microclimate on the water relationships of woodland, pasture, and cultivated soils. Soil Science Society Proceedings 7:95–104.
- Poulenard, J. , P. Podwojewski , J-L. Janeau , and J. Collinet . 2001. Runoff and soil erosion under rainfall simulation of Andisols from the Ecuadorian Páramo: effect of tillage and burning. Catena 45:185–207.
- Proulx, S. and J. Stein . 1997. Classification of meteorological conditions to assess the potential for concrete frost formation in boreal forest floors. Canadian Journal of Forest Research 27:953–958.
- Rice, W. R. 1989. Analyzing tables of statistical tests. Evolution 43:223–225.
- Rist, S. 1990. Vatns er þörf Reykjavík Menningarsjóður. 248 pp. (in Icelandic).
- Rybakova, N. A. 1990. Water permeability of frozen soils under tree stands in the forest-steppe zone. Soviet Soil Science 22:106–112.
- Saetre, P. 1998. Decomposition, microbial community structure, and earthworm effects along a birch-spruce soil gradient. Ecology 79:834–846.
- SAS The SAS system for Windows. Release 8.01. 2000. Cary, North Carolina SAS Institute Inc.
- Shanley, J. B. and A. Chalmers . The effect of frozen soil on snowmelt runoff at Sleepers River, Vermont. Hydrological Processes 1999. 13:1843–1857.
- Shoji, S. , M. Nanzyo , and R. A. Dahlgren . Volcanic Ash Soils. Genesis, Properties and Utilization 1993. Amsterdam Elsevier. 288 pp.
- Sigurdsson, S. Gudmundsson, H. , editor. Birch in Iceland. Skógarmál 1977. Reykjavík Edda. 146–172. In . (in Icelandic).
- Sigurjónsson, A. Sandgræðslan. Minnst 50 ára starfs Sandgræðslu Íslands [Fifty Years of Soil Conservation in Iceland] 1958. Reykjavík Búnaðarfélag Íslands og Sandgræðsla ríkisins. 359 pp. (in Icelandic).
- Soil Survey Staff Soil Taxonomy: a Basic System of Soil Classification for Making and Interpreting Soil Surveys. 2nd edition 1999. Washington, D.C U.S. Department of Agriculture, Natural Resources Conservation Service. 869 pp.
- Stoeckeler, J. H. and S. Weitzman . Infiltration rates in frozen soils in northern Minnesota. Soil Science Society Proceedings 1960. 24:137–139.
- Storey, H. C. Stefferud, A. , editor. Frozen soil and spring and winter floods. Water. The Yearbook of Agriculture 1955 1955. Washington, D.C U.S. Department of Agriculture. 179–184. In .
- Strachan, I. B. , O. Arnalds , F. Pálmason , H. Thorgeirsson , B. D. Sigurdsson , H. Sigurdardóttir , and G. Novoselac . Soils of the Gunnarsholt experimental plantation. Icelandic Agricultural Sciences 1998. 12:15–26.
- Thurow, T. L. Heitschmidt, R. K. and J. W. Stuth , editors. Hydrology and erosion. Grazing Management: an Ecological Perspective 1991. Portland, Oregon Timber Press, Inc. 141–159. In .
- Thurow, T. L. , W. H. Blackburn , and C. A. Taylor Jr . Hydrologic characteristics of vegetation types as affected by livestock grazing systems, Edwards Plateau, Texas. Journal of Range Management 1986. 39:505–509.
- Trimble, G. R. , J. R. Richard , S. Sartz , and R. S. Pierce . How type of soil frost affects infiltration. Journal of Soil and Water Conservation 1958. 13:81–82.
- Tyrtikov, A. P. Effects of Vegetation on the Freezing and Thawing of Soils.. 1976. New Delhi Amerind Pub. 196 pp.
- van der Kamp, G. , M. Hayashi , and D. Gallen . Comparing the hydrology of grassed and cultivated catchments in the semi-arid Canadian prairies. Hydrological Processes 2003. 17:559–575.
- Veðráttan, Monthly and Annual Weather Report 1999. Icelandic Meteorological Office, Reykjavík (in Icelandic).
- Veðráttan, Monthly and Annual Weather Report 2000. Icelandic Meteorological Office, Reykjavík (in Icelandic).
- Wada, K. , O. Arnalds , Y. Kakuto , L. P. Wilding , and C. T. Hallmark . Clay minerals of four soils formed in eolian and tephra materials in Iceland. Geoderma 1992. 52:351–365.